
Abstract
Senescence and osteogenic differentiation potential loss limited bone nonunion treatment effects of bone marrow-derived mesenchymal stem cells (BMSCs). MiR-100-5p/Lysine(K)-specific demethylase 6B (KDM6B) can inhibit osteogenesis, but their effects on bone union remain unclear. This study aims to investigate the effects of miR-100-5p/KDM6B on osteogenic differentiation and bone defects. Wild-type or microRNA 100 (miR-100) knockdown mice underwent critical-size defect (CSD) cranial surgery and collagen I/poly-γ-glutamic acid scaffold treatment. The crania was observed using microcomputed tomography, hematoxylin and eosin staining, Masson staining, alkaline phosphatase (ALP) staining, immunohistochemistry, and immunofluorescence. Primary-cultured BMSCs transfected with miR-100-5p mimic/inhibitor and KDM6B cDNA were evaluated for osteogenic differentiation using Alizarin Red staining, ALP activity detection, and Western blot analysis. Genetic transcription levels were detected using quantitative reverse transcription polymerase chain reaction. This study found that miR-100 depletion promotes defect healing in mouse calvaria, increases the proportion of new bone and osteoblasts in calvaria, and activates the expression of KDM6B and osteocalcin (OCN) proteins, promoting the transcription of bone morphogenetic protein-2, Runt-related transcription factor 2 (Runx2), OCN, and KDM6B, while methylation of lysine 27 on histone H3 (H3K27me3) decreased. Furthermore, miR-100-5p mimics suppressed osteogenic differentiation by inhibiting KDM6B with increased H3K27me3, ALP, Runx2, OCN, and osteopontin protein expression, while miR-100-5p inhibitors have opposite effects. Moreover, KDM6B can reverse miR-100-5p mimic effects. Notably, scaffolds carrying miR-100-5p mimics/inhibitors transfected BMSCs were placed in CSD mice and found that miR-100-5p inhibitors have a better effect on CSD healing and increase new bone without inflammatory cell infiltration. This study proved that miR-100-5p depletion promotes bone union and osteogenic differentiation of BMSCs via KDM6B/H3K27me3.
Impact Statement
This study proves that miR-100 depletion promotes defect healing in mouse calvaria, increases the proportion of new bone and osteoblasts in the calvaria, and activates the expression of KDM6B and osteocalcin (OCN) proteins, promoting the transcription and translation of bone morphogenetic protein-2, Runt-related transcription factor 2, OCN, and KDM6B. Furthermore, miR-100-5p in bone marrow-derived mesenchymal stem cells (BMSCs) suppressed osteogenic differentiation by inhibiting KDM6B. Moreover, BMSCs depleted with miR-100-5p have a better effect on mouse calvaria defect healing. This study provides scientific evidence and a new direction for the development of strategies for BMSC treatment of fractures.
Introduction
Most fractures can heal after clinical treatment. However, factors such as old age, diabetes, obesity, or large bone gaps may lead to slower healing and raise amputation risk. 1 Serious fractures can even lead to mortality. Approximately 33% of male hip fracture victims and 22% of female patients will die within a year, with old age, comorbidities, and gender being significant risk factors. 2 Moreover, nonunion is a major concern for fracture treatment, with 8.1% requiring rehospitalization within 2 years due to complications such as nonunion, imposing substantial financial and medical burdens on both the patient and society. 3 Bones, the body’s hardest structure, offer framework and protection for other tissues, facilitate movement, contribute blood cells, and can also maintain homeostasis of calcium and acidity/alkalinity. 4 After bone injuries, bone union is crucial in repairing defects, mainly driven by cells such as osteoblasts, osteoclasts, and osteocytes. 5 Generally, this process comprises three sequential stages: inflammation, repair, and bone remodeling, culminating in new bone formation. 6 Currently, autografts, allografts, xenografts, and synthetic biomaterials can be used to speed up bone union; however, chronic pain from nerve damage, implant rejection, and infection post-transplantation hinder bone union. 7 Ideal bone graft materials should exhibit superior biodegradable, biocompatible, mechanical, and osteogenic properties for treating bone defects. 8 However, the optimal therapeutic strategy for bone defects remains elusive, requiring further investigation into bone union biology.
Bone marrow-derived mesenchymal stem cells (BMSCs), progenitors of osteoblast, adipocyte, and chondrocyte, are essential in bone formation. 9 Evidence indicates that BMSC therapy is comparable to autografts for treating nonunion with fewer drawbacks; notably, BMSCs can enhance repair effect of scaffolds. 10 Also, BMSCs enhanced the healing of rat calvaria critical-size defects (CSDs). 11 However, the continuous culture of BMSCs in vitro often leads to cellular senescence that hinders bone formation potential, which adversely affecting BMSCs’ capacity at promoting bone union postimplantation. 12 Augmenting the osteogenic potential of BMSCs is pivotal for enhancing its use in bone nonunion treatment.
Histone methylation plays a key role in epigenetic control of bone union. It regulates mesenchymal stem cell osteogenic and adaptogenic differentiation. 13 Methylation of lysine 27 on histone H3 (H3K27me3) accumulation can inhibit the activation of Runt-related transcription factor 2 (Runx2) that is a key osteoblast marker gene. 14 Lysine(K)-specific demethylase 6B (KDM6B or JMJD3) is capable of demethylating Histone H3K27me2/3, which is associated with gene repression. 15 Research shows that KDM6B expression increased in osteogenic, while its downregulation suppresses osteogenesis. 16 Also, depletion of KDM6B significantly impairs osteogenic potential of BMSCs, promoting adipogenesis. 17 Furthermore, KDM6B can be recruited to the promoter regions of bone morphogenetic protein (BMP) and HOX genes, catalyzing H3K27me3 demethylation, thereby enhancing mesenchymal stem cell osteogenesis and suppressing adipogenesis. 18 These studies suggest that KDM6B can inhibit H3K27me3 to enhance osteogenesis and studying its regulatory molecules will promote the development of therapeutic strategies.
Noncoding microRNAs (miRNAs) are pivotal modulators of bone union and ideal therapeutic targets. 19 Ding et al. reported that miR-100 is highly expressed in osteoporotic patient plasma. 20 MiR-100-5p is a mature fragment of miR-100. Research reported that miR-100-5p showed significant negative correlation with bone density; it was significantly in higher levels in low bone mass groups. 21 Furthermore, miRNAs serve critical roles in regulating mesenchymal stem cell differentiation. 22 Wang et al. found that miR-100-5p overexpression inhibited the proliferation and osteogenic differentiation of human BMSCs. 23 Moreover, exosomes with high levels of miR-100-5p inhibit human BMSC osteogenesis. 24 Ai et al. reported that miR-100-5p has been found to antagonize the promoting effect of therapeutic drugs on KDM6B, suggesting that miR-100-5p is a regulator of KDM6B in promoting osteoporosis. 11 However, the involvement of miR-100-5p in bone union remains unclear.
Therefore, this study proposes a hypothesis that miR-100-5p negatively regulates KDM6B, which may inhibit the osteogenic differentiation of BMSCs and hinder the healing of CSDs. Depleting miR-100-5p may enhance the osteogenic differentiation of BMSCs and strengthen the bone union of CSD. By using cranial CSDs mouse models and miR-100 knockdown (KO) mice to evaluate the effects of miR-100-5p on BMSC osteogenic and bone union, this study aims to provide scientific support for the development of strategies for BMSC treatment of fractures.
Methods and Materials
Animal information and ethics
A total of 33 C57BL/6J mice (♀, 8 weeks) and 9 miR-100 KO C57BL/6J mice (♀, 8 weeks) were purchased from Shanghai SLAC Laboratory Animal Co., Ltd., Shanghai, China, and GemPharmatech Co., Ltd., Jiangsu, China. A standardized barrier system for housing mice was 50–60% relative humidity, 22 ± 2°C with alternating light/dark cycle every 12 h. Mice were housed in groups of 6 per cage. All animal experiments were approved by the Animal Experimentation Ethics Committee of Zhejiang Eyong Pharmaceutical Research and Development Center and the approval number is ZJEY-20230914-1. All institutional and national guidelines for the care and use of laboratory animals were followed.
Establishment of CSD model
CSD cranial surgery for wild-type (WT)/miR-100 KO (miR-100-/-) mice were performed after adapting to a feeding for 1 week. After isoflurane anesthesia, top of mouse heads was strictly disinfected and shaved. The parietal bone was exposed after a skin incision on the sagittal midline, then, CSD at 3.5 mm were created using a trephine, meanwhile, keeping the dura mater undisturbed. 25 After placing the blank scaffolds according to the grouping requirements, the skin incision was sutured. The blank scaffold (collagen I/poly-γ-glutamic acid scaffold) was prepared according to Cho et al.’s 26 report. Briefly, collagen I (1 mL, 13.3 mg/mL, HY-NP109, MCE, USA), poly-γ-glutamic acid (222 µL, 20 mg/mL, G1049, Sigma-Aldrich, USA), and ProNectin® F Plus (100 µL, 1 mg/mL, F8141, Sigma-Aldrich, USA) were stirred and mixed in sequence. Triple antibiotic ointment and bupivacaine were applied topically. After 0, 4, and 8 weeks, mice were euthanized with carbon dioxide (CO2), and skull defect tissue was removed for fixation in 10% formalin or freezing at −80°C. WT and miR-100 KO mice were divided into two groups (n = 6). Furthermore, a 20 µL blank scaffold was mixed with 1 × 106 primary BMSCs for animal experiments. 26 Primary BMSCs were derived from C57BL/6J, mice and the method was described in the “Isolation and culture of primary BMSCs” section. WT CSD mice were treated with cell scaffolds inoculated with autologous BMSCs; these BMSCs were transfected with miR-100-5p mimics/inhibitors and their control and were divided into four groups: mimics-negative control (NC), miR-100-5p mimics, inhibitor-NC, and miR-100-5p inhibitors.
CSD area analysis
Microcomputed tomography (micro-CT) was used to measure defect area. MCT-III micro-CT machine (Zhongkekaisheng/ZKKS, Guangzhou, China) analyzed calvarias under the following conditions: voltage, 70 kV; current, 200 µA; exposure time, 300 ms; and resolution, 14.8 µm. Then, 3D images were reconstructed using NRecon software (ZKKS), and the defect areas were measured using ImageJ (National Institutes of Health [NIH], MD, USA). Regenerated bone area = Defect Area0-week − Defect Area4/8-week
Histological analysis
Calvarias were fixed and decalcified. Subsequently, samples were washed and dehydrated, followed by burial in paraffin. Paraffin blocks were cut to 6 µm serial sections. Sections were used for hematoxylin and eosin (HE) staining and Masson staining. For HE staining, sections were dewaxed to water using xylene and ethanol and stained with hematoxylin (H3136, Sigma, MO, USA) and eosin (E4009, Sigma) dye in a sequence. For Masson staining, sections were also dewaxed to water and then stained for 10 min using Weigert’s iron hematoxylin dye (R20387, Yuanye, Shanghai, China). After differentiation using hydrochloric alcohol, sections were stained for 10 min using a Ponceau acid fuchsin dye (71033761, Sinopharm Group Chemical Reagent, Shanghai, China), followed by differentiating for 15 min using phosphomolybdic-phosphotungstic acid solution (XW514297441, OKA, Beijing, China). Afterward, the samples were transferred into an aniline solution and stained for 5 min. Following a rinse with water, the sections were dehydrated using absolute ethanol and subsequently exposed to xylene. At the last, the sections were coated with neutral resins for sealing. Tissues were observed under a Nikon Eclipse light microscope (Tokyo, Japan) and a panoramic scanner (KF-PRO-120, KFBIO, Zhejiang, China).
Alkaline phosphatase staining
Alkaline phosphatase (ALP) is a marker of mature osteoblasts. ALP staining kits (GP1050, Servicebio, Wuhan, China) were used to analyze mature osteoblasts. Briefly, tissue paraffin sections were dewaxed using a 60°C oven for 1 h. Tissue sections were stained using 500 µL ALP dye for 30 min. The ALP dye is prepared by mixing working solution A (40 µL), B (40 µL), and buffer solution (1 mL). Finally, the tissue sections were sealed, photographed under a Nikon Eclipse light microscope, and analyzed using ImageJ (NIH). Three slices from each group were selected at random. At least three different fields of each slice were chosen for ImageJ analysis. ImageJ was used to calculate the proportion of brown area.
Immunohistochemistry for KDM6B and osteocalcin
Dewaxed tissue sections were soaked in gradient concentrations of ethanol. Then, 100 µL H2O2 was used to incubate tissues for blocking the activity of endogenous peroxidase. Subsequently, sections were performed hot antigen repair by boiling Tris-ethylenediaminetetraacetic acid (E9884, Sigma) and then were blocked using 5% Bovine Serum Albumin (B2064, Sigma) for 20 min at 20 ± 2°C and stained by incubating with anti-KDM6B (ab38113, Abcam, Cambridge, UK) or anti-osteocalcin (OCN) (ab198228, Abcam) antibodies at 4°C overnight. The next day, the sections were washed and incubated with 50 µL goat antirabbit secondary antibodies (ab97080, Abcam) at 37°C for half an hour. And the sections were incubated with diaminobenzidine (36201ES03, YEASEN, Shanghai, China) for chromogenic reaction and then with hematoxylin (Bry-0001–01, Runnerbio, Shanghai, China) for counterstaining. After dehydration and transparent, the sections were sealed with neutral gum (10004160, Sinopharm Group Chemical Reagent) and were photographed and analyzed with ALP staining.
Immunofluorescence for H3K27me3+ Nestin+ cells
Immunofluorescence (IF) was used for colocalization to calculate the proportion of H3K27me3-positive BMSCs cells. Its method is like immunohistochemistry (IHC), except for the use of anti-Nestin (DF7754, Affinity, OH, USA), anti-H3K27me3 (DF6941, Affinity), and goat antirabbit secondary antibodies (ab150079, Abcam). Subsequently, tyramine salt-488/salt-cy3 (Bry-try488/trycy3, Runnerbio) were added and incubated avoiding exposure to light for 20 min at 20 ± 2°C. Finally, the antifade mounting medium (P0131, Beyotime, Shanghai, China) was used to seal the sections. ImageJ (NIH) was used to analyze the fluorescence intensity of green and red. Three slices from each group were selected at random. At least three different fields of each slice were chosen for ImageJ analysis. The mean ratio of positive cell numbers to total cells is used for statistical analysis.
Isolation and culture of primary BMSCs
Sterile femur and tibia were isolated from euthanized and C57BL/6J mice. Alpha-modified minimal essential medium (α-MEM) (M6074, sigma, MO, USA) was added with 10% fetal bovine serum (FS301-02, TransGen, Beijing, China) and 1% penicillin-streptomycin (FG101-01, P/S, TransGen) as complete medium. It was used to flush the bone marrow cavity and collect bone marrow plugs. The mesenchymal population was isolated on the basis of its ability to adhere to the culture plate as Li et al. reported. 27 After filtering, cells were cultured in complete medium and placed in an incubator at 37°C with 5% CO2. Passage 4 is used for subsequent experiments. A AE2000 microscope (Motic, Xiamen, China) was used to observe cell morphology.
Identification of BCMS by flow cytometry
According to previous reports, 28 CD90 and CD44 are BMSC’s antigens; CD34 and CD31 are not BMSC’s antigens. The antimouse CD90 (553016, BD, NJ, USA), antimouse CD44 (560570, BD), CD31 (553373, BD), and CD34 monoclonal antibody (56–0341-82, Invitrogen, CA, USA) were used for flow cytometry. Therefore, cell suspension at 1 × 106/mL was incubated with above antibodies at 4°C for 30 min avoiding light. After phosphate-buffered saline washing and resuspension, 400 µL cells were used for flow cytometry analysis on a flow cytometer (NovoCyte, Agilent, CA, USA). The homotypic monoclonal antibody was used as a control. CD90 and CD44 positivity rate is used to analyze the purity of BMSCs.
Cell transfections and osteogenic induction
Cell transfections were performed according to Lipofectamine® 3000 (L3000-008, Invitrogen) (Lipo3000)’s instructions and previous reports. 29 Lentiviruses carrying KDM6B cDNA plasmid or small interfering RNA (si)-KDM6B were designed and purchased from Shanghai GeneChem Co., Ltd., Shanghai, China. Cells at 2 × 105 per well were cultured in 6-well plates with 2 mL complete medium. Then, half of complete medium was replaced with α-MEM, which added lentiviruses (multiplicity of infection/MOI = 50), and cells were cultured for 4 h, followed by addition of 1 mL α-MEM again. After 48 h, the transfected culture medium was replaced with fresh complete culture medium. Then, on day 4, puromycin selection was performed (1 µg/mL) for 7 days. 30 Lipo3000 was used for transfecting miR-100-5p mimics, inhibitors, and their control according to its instructions. Briefly, 5 × 104/mL cells were cultured in 500 µL complete medium on 24-well plates. Opti-MEM (31985–070, Gibco, CA, USA) and working solution from Lipo3000 were mixed and added into cell cultured medium for 15 min incubation. Finally, fresh complete culture medium was used to culture transfected BMSCs. Furthermore, all BMSCs were cultured in an osteogenic induction medium: 31 Dulbecco's Modified Eagle Medium includes 10 mM β-glycerophosphate, 100 nM dexamethasone, and 50 µM ascorbate-2-phosphate.
Grouping of in vitro experiments
To explore the effects of miR-100-5p/KDM6B on osteogenic induction of BMSCs, we compared cells transfected with miR-100-5p mimics/inhibitors to their NC cells and divided them into control, mimic-NC, miR-100-5p mimic, inhibitor-NC, and miR-100-5p inhibitor groups. Additionally, BMSCs overexpressing KDM6B (named KDM6B) or NC were exposed to miR-100-5p mimic and its NC, while KDM6B silencing (named si-KDM6B [small interfering RNA targeting KDM6B])/si-NC BMSCs were used; they were divided into control, miR-100-5p mimic, miR-100-5p mimic+KDM6B, si-NC, and si-KDM6B groups. All cells were cultured for 72 h followed by performing detection and analysis.
Quantitative reverse transcription polymerase chain reaction
The expression levels of osteogenic differentiation marker genes in the cranial bones and the transfection efficiency of BMSCs cells were analyzed using quantitative reverse transcription polymerase chain reaction (qRT-PCR). A total RNA extraction reagent kit (10606ES60, YEASEN) and a miRNA extraction reagent kit (19331ES50, YEASEN) were used to collect total RNA from cells and cranial bones followed by total RNA was stratified using chloroform and centrifuged. The topmost liquid (containing RNA) was collected. After ethanol precipitation purification and detection of concentration and purity, they were used for reverse transcription and qRT-PCR. miRNA 1st Strand cDNA Synthesis Kit (11148ES10, YEASEN) and miRNA Universal qPCR SYBR Master Mix Kit (11171ES08, YEASEN) were used to measure miR-100-5p gene levels. PrimeScript™ reverse transcription kits (RR037Q, Takara, Osaka, Japan) and qPCR SYBR Green kits (11201ES08, YEASEN) were used to measure osteogenic differentiation marker gene levels. The reagent ratio and experimental conditions were carried out according to the instructions. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6 serve as internal reference genes for RNA and miRNA, respectively. The analysis method used was the 2-ΔΔCt method, 32 and primer information is shown in Table 1.
Primer Information
BMP2, bone morphogenetic protein 2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; OCN, osteocalcin; Runx2, runt-related transcription factor 2.
Evaluation of osteogenic differentiation
The osteogenic differentiation degree of BMSCs was analyzed using Alizarin Red staining and ALP activity detection. Alizarin Red staining can evaluate the formation of mineralized matrix nodules. Briefly, 5 × 105 per well of BMSCs were seeded into 12-well plates for 72-h osteogenic differentiation and were fixed using 4% paraformaldehyde for 10 min, followed by permeabilization with 0.3% TritonX-100 (FG301-01, TransGen). Then, cells were stained using 1% Alizarin Red dye (G1452, Solarbio, Beijing, China) for 20 min. Calcium precipitates were orange and observed under a microscope (Ts2-FC, Nikon). The area of orange mineral precipitation (%) is used for semiquantitative evaluation using ImageJ (NIH). Additionally, BMSCs at 2000 cells per well were seeded into 96-well plates and cultured for 72 h. Then, cells were lysed and centrifuged to obtain supernatants for ALP activity detection according to the instruction of an ALP kit (P0321S, Beyotime, Shanghai, China). In brief, working reagents were added into cell supernatants, and then they were incubated for 10 min and absorbance was measured using a microplate reader at a wavelength of 405 nm.
Western blot for detecting protein expression
After collecting total proteins of BMSCs, protein samples at 20 µg were added into sodium dodecyl sulfate-polyacrylamide gel electrophoresis glue holes to protein isolate. Then, samples were electroblotted onto a polyvinylidene fluoride membrane (10600023, GE Healthcare Life, PA, USA). After blocking and washing, samples were incubated with antibodies at 4°C overnight. Some antibodies were from Affinity: anti-KDM6B (1: 5000, DF13101), H3K27me3 (1: 5000, DF6941), BMP2 (1: 5000, AF5163), ALP (1: 5000, DF6225), OCN (1: 5000, DF12303), and osteopontin (OPN) (1: 5000, AF0227) antibodies, others were from the Bioss, Beijing, China: Runx2 antibodies (1: 5000, bs-1134R) and GAPDH (1:10000, bs-10900R). Subsequently, samples were incubated with a goat antirabbit secondary antibody (1: 5000, ab97080, Abcam). Finally, the washed membranes were incubated with the developer, were photographed in a chemiluminescence imager (ChemiScope 6000, CLINX, Shanghai, China), and were analyzed for relative grayscale values using ImageJ (NIH).
Statistics
Statistical analysis was performed with SPSS 20.0 (IBM, NY, USA). Data were reported as mean ± standard deviation. All data conformed to normal distribution. Student’s t-test was used to compare two independent groups. Additionally, multiple group comparisons were assessed using one-way analysis of variance followed by Tukey tests. A p value < 0.05 deemed statistically significant.
Results
MiR-100 knockout promotes CSD healing in mouse calvaria
For observing effects of miR-100 on CSD healing, we used miR-100 KO mice and WT C57BL/6 mice to establishing CSD models in mouse calvaria. At 0, 4, and 8 weeks, we measured the defect area using a micro-CT system. The regenerated bone areas in miR-100 KO mice were about 76% and 212% larger than those in WT mice, respectively (p < 0.01) (Fig. 1A). Furthermore, we used HE, Masson, and ALP staining kits for the histological analysis in calvaria. From the HE staining results, it was observed that the miR-100 KO group has more newly formed bones and less defect areas; furthermore, the western blot (WB) group showed more obvious inflammatory envelopes (Fig. 1B). Using Masson staining for observing nascent bone (blue) and mature bone (red), the WT group tissues showed a larger proportion of mature bone (Fig. 1C). ALP is a marker of mature osteoblasts, it was brown in Figure 1D, and there were more brown areas in miR-100 KO mouse tissues (p < 0.01). Micromorphological analysis results suggest that miR-100 KO promoted CSD healing.
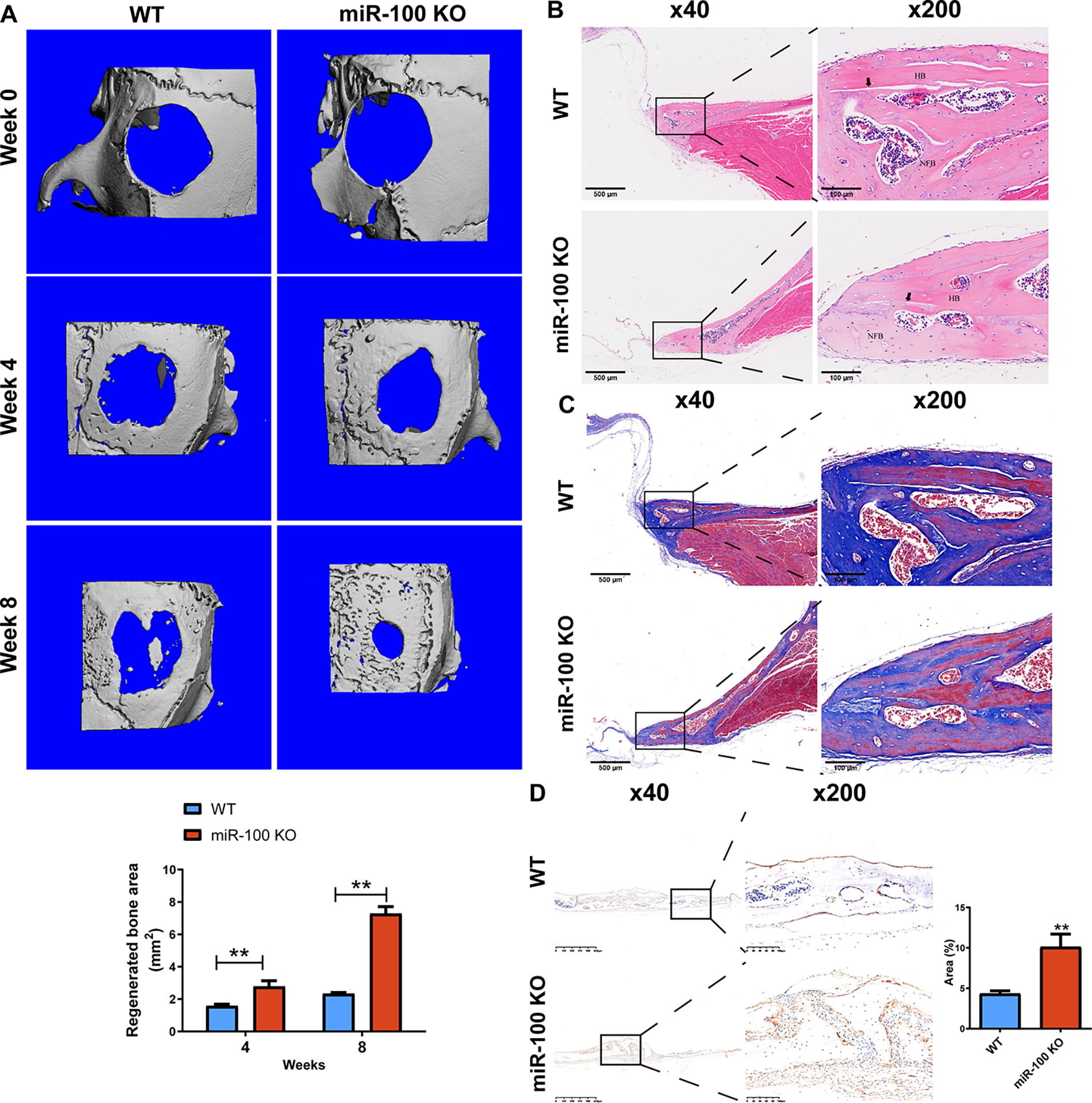
Microstructure and histological analysis of critical-size defects (CSDs) in the calvaria of miR-100 knockout mice. Wild-type (WT) C57BL/6J mice (♀, 8 weeks) and 9 miR-100 knockdown (KO) C57BL/6J mice (♀, 8 weeks) were used to establish CSD mouse models.
MiR-100 knockout increased osteogenic differentiation gene expression in mouse calvaria
IHC and IF staining are used to observe protein expression and localization in tissues. KDM6B and OCN expression in the miR-100 KO group increased by 43.7% and 88.1% compared to the WT group, respectively (p < 0.01) (Fig. 2A, B). The H3K27me3 and Nestin co-IF staining showed that H3K27me3+ BMSCs were decreased in the miR-100 KO group (Fig. 2C).

KDM6B, OCN, H3K27me3, and Nestin expression in the calvaria of miR-100 KO mice. WT C57BL/6J mice (♀, 8 weeks) and 9 miR-100 KO C57BL/6J mice (♀, 8 weeks) were used to establish CSDs mouse models.
Additionally, we used qRT-PCR to measure miR-100 and osteogenic differentiation gene levels (BMP2, Runx2, OCN, and KDM6B). MiR-100 levels decreased over 70% in miR-100 KO mice than these in WT mice; meanwhile, levels of BMP2, Runx2, OCN, and KDM6B genes increased 44.8%, 192.6%, 72.3%, and 116.5%, respectively (p < 0.01) (Fig. 3). The results suggest that miR-100 KO may promote CSD healing.

MiR-100 and osteogenic differentiation marker gene transcription levels. The transcription levels of miR-100, BMP2, Runx2, OCN, and KDM6B genes in the cranial bones were analyzed using quantitative reverse transcription polymerase chain reaction. Student’s t-test was used, and data were reported as mean ± standard deviation. *: means p < 0.05; **: means p < 0.01. BMP2, bone morphogenetic protein 2; Runx2, Runt-related transcription factor 2.
Establishing MiR-100-5p/KDM6B overexpression/silencing BMSC models
The combined use of BMSCs and collagen I/poly-γ-glutamic acid scaffolds represents a potential therapeutic strategy for bone repair. Enhancing BMSCs’ osteogenic differentiation ability can promote bone healing. 33 Therefore, we explored the effects of miR-100-5p on BMSCs. First of all, we collected primary BMSCs with spindle shape from mouse bone marrow based on the BMSCs’ ability to adhere to the culture plate (Fig. 4A). Flow cytometry is used to identify the purity of isolated BMSCs. CD90 and CD44 positive rates were 98% and 97.4%, while for CD34 and CD31 were 0.5% and 0.7% (Fig. 4B).
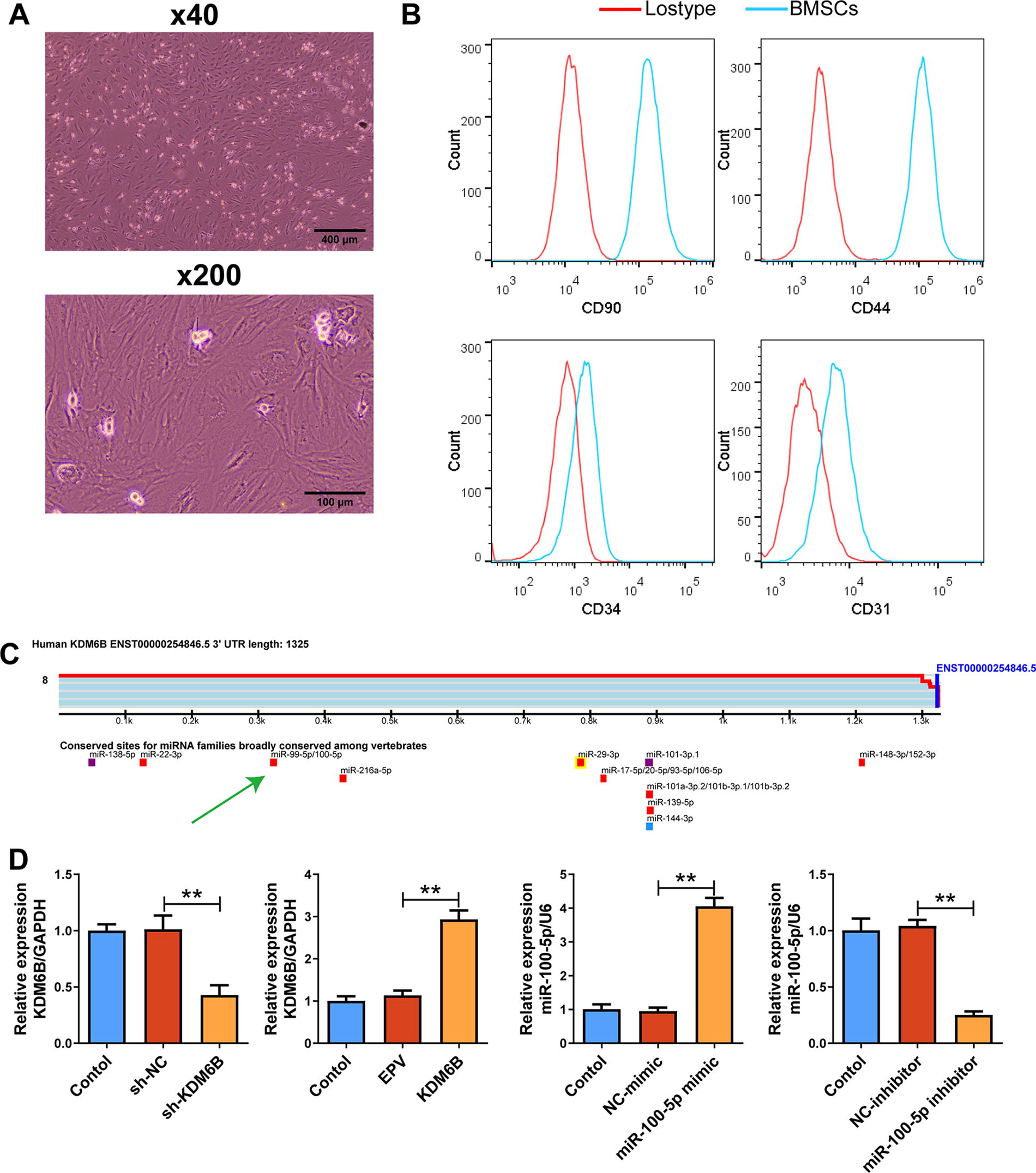
Detecting purity of primary bone marrow-derived mesenchymal stem cells (BMSCs) and gene transcription level posttransfection. Primary BMSCs are isolated from WT C57BL/6J mice.
Target Scan Release7.2 database search (key word: KDM6B) showed that KDM6B is a downstream target of miR-100-5p (Fig. 4C). For observation of their effects on osteogenic differentiation of BMSCs, we used cell transfection to regulate gene expression. The results of qRT-PCR showed that KDM6B mRNA decreased over 57.8% in KDM6B silenced BMSCs and increased 159.2% in KDM6B overexpression BMSCs (p < 0.01) (Fig. 4D). Furthermore, miR-100-5p mimics and inhibitors were used to establish miR-100-5p overexpression or silenced BMSCs. MiR-100-5p increased 325.7% in the miR-100-5p mimic group, while miR-100-5p decreased 75.9% in the miR-100-5p inhibitor group (p < 0.01) (Fig. 4D). We have successfully established BMSC models with KDM6B or miR-100-5p overexpression/silencing.
MiR-100-5p inhibits osteogenic differentiation of BMSCs
Cell mineralization is a definitive marker of osteoblast differentiation. We evaluated it by using Alizarin Red staining and ALP staining assays. Alizarin Red staining can reveal apparent orange-red calcium nodules. In the miR-100-5p mimic group, red calcium nodule number was decreased compared with its control (p < 0.01) (Fig. 5A, B) with decreased ALP concentration (p < 0.01) (Fig. 5C). On the contrary, the red calcium nodule number increased in the miR-100-5p inhibitor group, meanwhile, ALP concentration raised, compared with the inhibitor-NC group (p < 0.01) (Fig. 5A–C).

MiR-100-5p mimics/inhibitors regulated osteogenic differentiation of BMSCs. Lipofectamine® 3000 was used for transfecting miR-100-5p mimics, inhibitors, and their control into BMSCs. Osteoblast differentiation was evaluated by using
Furthermore, we measured the levels of osteogenic differentiation-related proteins using WB assay. Expression levels of KDM6B, BMP2, ALP, Runx2, OCN, and OPN were decreased in the miR-100-5p mimic group, while H3K27me3 increased (p < 0.01) (Fig. 5D–K). As expected, miR-100-5p inhibitors had opposite effects on these proteins compared with its controls (p < 0.01) (Fig. 5D–K). Together, these results support the inhibition of miR-100-5p on osteogenic differentiation of BMSCs.
KDM6B functions as a downstream target of miR-100-5p to regulate BMSCs’ osteogenic differentiation
Through Alizarin Red staining and ALP staining assays, we noticed that KDM6B overexpression increased orange-red calcium nodules and ALP concentration in miR-100-5p mimic transfected cells (p < 0.01) (Fig. 6A–C). As expected, KDM6B overexpression activated protein KDM6B, BMP2, ALP, Runx2, OCN, and OPN expressions in miR-100-5p mimic transfected BMSCs, while H3K27me3 expression was reduced (p < 0.01) (Fig. 6D–K). To clarify the crucial role of KDM6B, we also inactivated KDM6B in BMSCs. After KDM6B silencing, orange-red calcium nodules and ALP concentration were both reduced (p < 0.01) (Fig. 6A–C). BMP2, ALP, Runx2, OCN, and OPN expressions were also reduced with decreased KDM6B expression, while H3K27me3 expression increased (p < 0.01) (Fig. 6D–K). These observations suggest that KDM6B may inhibit KDM6B to antagonize BMSCs’ osteogenic differentiation.

MiR-100-5p overexpression inhibited osteogenic differentiation of BMSCs via KDM6B. Lentiviruses and Lipofectamine® 3000 were used for transfecting plasmid carrying KDM6B gene, si-KDM6B, and miR-100-5p mimics into BMSCs. Osteoblast differentiation was evaluated by using
MiR-100-5p overexpression in BMSCs suppressed CSD healing of mouse calvaria
To verify the role of miR-100-5p overexpression/silencing BMSCs in mouse CSD healing, we established CSD models using WT C57BL/6 mice. CSD healing and bone maturation were evaluated using micro-CT and Masson staining. After 4 weeks of treatment, we noticed that compared with its control (the mimic-NC group), the miR-100-5p mimic group had a nearly 40% decrease in regenerated bone areas (p < 0.05) (Fig. 7A). Additionally, the miR-100-5p inhibitor groups had a 74% increase in regenerated bone areas compared with the inhibitor-NC group (p < 0.01) (Fig. 7A). According to Masson results, collagen fibers can be seen in the healing areas of the miR-100-5p mimic group’s bone tissue, and the areas of nascent bone tissue are smaller than those of the mimic-NC group. Additionally, there are more nascent bone and mature bone tissue in the healing area of miR-100-5p inhibitor groups compared with its control. Histological analysis was performed using HE staining. In the miR-100-5p mimic group, defect areas were larger with more collagen fibers compared with the NC group (Fig. 8). The defect areas were smaller, and more mature bone tissue was present with better integration in the miR-100-5p inhibitor group compared with the inhibitor-NC group (Fig. 8). These results suggest that miR-100-5p overexpression in BMSCs suppressed CSD healing of mouse calvaria, and silencing of miR-100-5p of BMSCs can promote CSD healing.
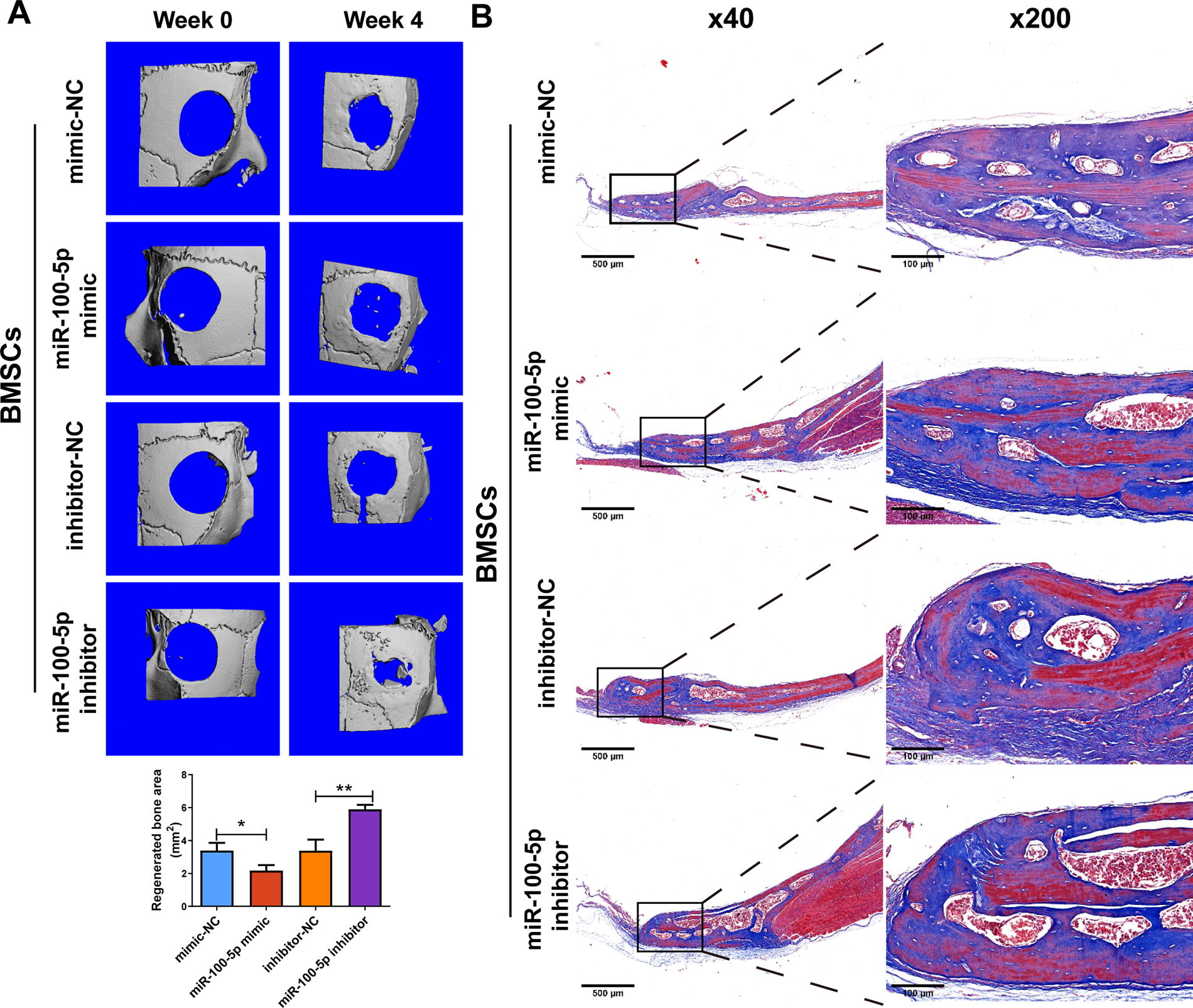
Microcomputed tomography and Masson staining for calvaria CSDs. CSD healing and bone maturation in C57BL/6J mice with BMSCs were evaluated using

Hematoxylin and eosin staining for calvaria CSDs. CSD healing and bone maturation in C57BL/6J mice with BMSCs were evaluated using hematoxylin and eosin staining. Before CSD calvaria surgery, BMSCs were transfected with miR-100-5p mimics, inhibitors, or their controls. One-way ANOVA followed by Tukey tests were used, and data were reported as mean ± standard deviation. *: means p < 0.05.
Discussion
This study evaluated the bone union of miR-100 KO/WT mice with CSD modeling and found that miR-100 KO accelerated the healing of calvaria defects with less inflammatory cell infiltration and more new bone and osteoblasts. Meanwhile, KDM6B expression increased and H3K27me3-positive osteoblasts were decreased. Furthermore, BMP2, Runx2, and OCN, the osteogenic markers, are increased after miR-100-5p silencing. The results from previous research 11 and the Target Scan Release7.2 database searches suggest that miR-100-5p has a regulatory effect on KDM6B. Wang et al. reported that miR-100-5p can inhibit the osteogenic differentiation of human BMSCs. 23 Liu et al. proved that KDM6B was recruited to BMP2 promoters improving osteogenic differentiation. 18 The results of this study correspond to previous reports and found that miR-100-5p mimic can inhibit BMSC osteogenic differentiation. Notably, KDM6B overexpression reversed it. Meanwhile, miR-100-5p silencing activated KDM6B to promote osteogenic differentiation of BMSCs. This study provides new evidence in vitro and in vivo that miR-100-5p blocks osteogenic differentiation by inhibiting KDM6B. MiR-100-5p not only has the potential to serve as a biomarker 21 but also as a key molecule for bone nonunion.
BMP2 is an osteogenic growth factor and can promote BMSC proliferation, also inducing osteogenic differentiation. 34 Wu et al. reported that KDM6B overexpression in dental pulp stem cells activated BMP2 gene by demethylating H3K27me3 in the promoter region of BMP2. 35 In line with this, this study found that KDM6B silencing increased H3K27me3 and decreased BMP2, Runx2, OCN, and OPN. Notably, BMP2/Smad/Runx2 axis promoted mouse bone formation. 36 Furthermore, this study demonstrated that miR-100-5p silencing activated KDM6B to promote osteogenic differentiation and bone formation of BMSCs via a mechanism involving the BMP2-Runx2 axis.
Runx2 is an important transcription factor for bone union of CSD. This study found that KDM6B overexpression inhibits H3K27me3 level to increase mineral nodule formation and ALP secretion in BMSCs, as well as Runx2 expression. Fitting with these results, research shows that KDM6B inhibits H3K27me3 to enhance osteogenesis via Runx2; H3K27me3 accumulation can inhibit the activation of Runx2.14,15 Ai et al. found that miR-100-5p/KDM6B can directly target RUNX2. 11 This suggests that miR-100-5p may directly target BMP2 and Rux2 transcription simultaneously, inhibiting osteogenic differentiation.
Bone nonunion is already the main reason for a poor quality of life and secondary hospitalization surgeries in fracture patients after surgery. 3 BMP2 is well known as an outstanding growth factor to promote bone union. Researchers are committed to developing BMP2-related drug formulations for bone nonunion treatment; but due to its serious side effects such as cervical and soft tissue swelling, airway compromise, and need for reoperation, its application is limited.37,38 The main reason for the occurrence of BMP2 adverse reactions may be that high doses of BMP2 have proinflammatory potential. 39 In this study, miR-100 KO did not cause inflammatory cell infiltration. It suggests that miR-100-5p inhibitors have the potential to serve as a replacement or adjuvant therapy for BMP2, although this hypothesis requires further clinical and basic research evidence to support it.
BMSCs have the advantages of easy availability and low immunogenicity for the treatment of bone defects. However, BMSCs with aging and loss of osteogenic differentiation potential in vitro culture limited their clinical applications. 12 Wang et al. reported that miR-100-5p can inhibit osteogenic differentiation of human BMSCs. 23 Also, miR-100-5p promotes cell proliferation. 40 The proliferation ability of cells is usually inversely proportional to their differentiation ability. Moreover, this study demonstrated that miR-100-5p-inhibited BMSCs have a better effect on improving CSD mice compared with the control BMSCs. Toyama et al. found that miR-100-5p was related with senescent vascular endothelial cells. 41 Overexpression of miR-100-5p may inhibit osteogenic differentiation and proliferation of BMSCs by promoting cell aging, which deserves further investigation.
In conclusion, this study used miR-100 KO and WT mice and proved that miR-100 depletion promotes defect healing in mouse calvaria, increases the proportion of new bone and osteoblasts in the calvaria, and activates the expression of KDM6B and OCN proteins, promoting the transcription and translation of BMP2, Runx2, OCN, and KDM6B. Furthermore, miR-100-5p in BMSCs suppressed osteogenic differentiation by inhibiting KDM6B. Moreover, BMSCs depleted with miR-100-5p have a better effect on mouse calvaria defect healing. This study provides scientific evidence and new direction for the development of strategies for BMSC treatment of fractures.
Ethics Approval and Consent to Participate
All animal experiments were approved by the Animal Experimentation Ethics Committee of Zhejiang Eyong Pharmaceutical Research and Development Center and the approval number is ZJEY-20230914-1. All institutional and national guidelines for the care and use of laboratory animals were followed.
Footnotes
Authors’ Contributions
X.G.: Conceptualization, data curation, funding acquisition, investigation, project administration, and writing—original draft. X.C.: Conceptualization, formal analysis, investigation, methodology, and writing—original draft. Z.M.: Data curation, formal analysis, investigation, and visualization. J.H.: Formal analysis, investigation, and methodology. S.J.: Investigation, methodology, and validation. W.W.: Validation and visualization. L.L.: Conceptualization, project administration, supervision, and writing—review and editing. X.Z.: Conceptualization, project administration, supervision, and writing—review and editing.
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare that there is no conflict of interest regarding the publication of this article.
Funding Information
This work was supported by the