
Abstract
Volumetric muscle loss (VML) due to traumatic injury results in the abrupt loss of contractile units, stem cells, and connective tissue, leading to long-term muscle dysfunction and reduced regenerative potential. Muscle connective tissue contains a proregenerative extracellular matrix (ECM), and our lab harnesses the regenerative capacity of decellularized muscle matrix (DMM) to treat VML, a condition with limited treatment options. However, a major limitation is that muscle often comes from aged donors. Previous work from our lab showed that aged donor muscle contains higher levels of advanced glycation end-product (AGE) cross-links compared to muscle from younger donors. This study aimed to determine whether increased AGE cross-links reduce the regenerative capacity of DMM. To test this, we first generated AGEs in DMM with direct D-ribose incubation. We then removed ∼35% of the gastrocnemius muscle in a model and treated it with either AGE-DMM or standard DMM (no AGEs), comparing results to controls. Although muscle force results remained unchanged between AGE-DMM and DMM, AGEs led to reduced muscle mass in histological sections, fewer fibers, and smaller fiber diameters. AGEs also increased collagen levels in histology, but protein assays showed reduced collagen production. We investigated the canonical receptor for AGEs, the receptor for AGEs (RAGE), and found elevated levels in AGE-treated VML compared to DMM alone, along with increased levels of the noncanonical receptor galectin-3. Both RAGE and galectin-3 are associated with inflammation, and proteomics revealed higher inflammatory markers in AGE-treated muscle than in DMM alone. In conclusion, our data suggest that AGEs impair the regenerative potential of DMM, highlighting the importance of considering donor age when sourcing muscle for DMM therapies.
Impact Statement
This study investigates advanced glycation end-product cross-links in skeletal muscle extracellular matrix (ECM) as a way to model its deleterious effects on muscle regeneration in vivo. We demonstrate here that ECM glycations reduce muscle regeneration, enhance inflammatory markers, reduce ECM protein production, and proteomic analysis identified unique targets that could be explored in future research endeavors.
Introduction
Skeletal muscle extracellular matrix (ECM) plays a crucial role in muscle development and regeneration through muscle-specific cues.1,2 Decellularized muscle matrix (DMM), which is processed skeletal muscle ECM, retains key promyogenic characteristics, including an aligned architecture and essential chemical components that promote muscle regeneration. 3 By removing cellular content, DMM reduces the immunogenic response while preserving the structural benefits of the native ECM. 4 These features make skeletal muscle-derived ECM a promising bio-scaffold for supporting muscle regeneration. Muscle stem cells (MuSCs), fibroblasts, macrophages, and other supporting cells interact with the ECM to regulate regeneration. 5 However, the impact of the donor’s age on DMM’s effectiveness in muscle regeneration has not been thoroughly studied. 6
According to the Organ Procurement and Transplantation Network, 33% of donated tissues are from donors aged 50 years or older (as of March 13, 2023). Given the prevalence of muscle injuries in individuals 65 years and older, this study aims to explore how donor age affects the regenerative capacity of ECM.7,8 Previously, we found that aged mouse muscle (∼20 months old) accumulated advanced glycation end products (AGEs) 6 a type of nonenzymatic cross-link formed on the ECM through the Maillard reaction. AGEs tend to accumulate on collagen fibers due to the presence of repeating arginine and lysine residues, which enhance the Maillard reaction by allowing monosaccharides to interact stochastically with proteins. Furthermore, these glycation cross-links stiffen the collagenous matrix, suggesting that AGE accumulation in the ECM may negatively impact muscle regeneration. 9 To test the effect of AGEs on muscle regeneration, we used a volumetric muscle loss (VML) injury model, which was developed in our lab. 3 Our previous research demonstrated that DMM scaffolds could promote muscle regeneration in such injuries3,10,11; however, these scaffolds were derived from young rodent muscles, which do not represent the typical donor age.
To model aged rat ECM, we followed a previously established protocol to induce AGE cross-links in young DMM by exploiting the Maillard reaction with D-ribose. 12 We found that using 250 mM D-ribose produced a concentration of AGE cross-links comparable to that observed in natural aging. We then hypothesized that increased AGEs would diminish DMM-mediated muscle regeneration in a Sprague-Dawley gastrocnemius VML model. VML injuries enhance fibrogenesis and inflammation, involving a complex array of cells that respond to the injury site, so we selected a proteomic approach to characterize any factors involved in inflammation and ECM remodeling. 13 The goal of this study was to use the VML model to understand how AGEs might limit the regenerative capacity of bio-scaffolds by affecting muscle differentiation, fibrosis, and inflammation.
Materials and Methods
Decellularization and D-ribose incubation
Decellularization involves the complete removal of cellular elements, including DNA and organelles. 14 SAS Sprague-Dawley rats (250–300 g) were obtained from Charles River (Wilmington, MA), euthanized with CO2, and the skin was dissected from the hindlimbs following a VCU-approved protocol (IACUC No. AD10000675). The biceps femoris muscles were removed, and the gastrocnemius muscles were extracted at the Achilles tendon and proximal to the posterior knee joint, ensuring that the plantaris and soleus muscles were separated and removed. The gastrocnemius muscles were frozen at −80°C, then thawed in a 37°C water bath before decellularization. Both heads of the gastrocnemius muscle were butterflied to a 3–5 mm thickness to ensure thorough penetration of the decellularization solutions. Muscles were decellularized in 50 mL beakers with constant stirring using a proprietary method involving enzyme and detergent treatments. Peracetic acid (0.5% for 24 h) was used at the end of the process for sterilization. DMMs were then frozen at −80°C, lyophilized overnight, and stored at −80°C.
For all AGE formation experiments, DMMs were incubated in 100- and 250-mM D-ribose (Sigma-Aldrich, St. Louis, MO) solution (2.22 mL/mg dry DMM weight) in six-well plates in vitro for 5 days at 37°C with 5% CO2. Excess D-ribose was removed with three 5-min washes in 10 mL deionized water. To avoid additional freeze-thaw cycles, DMMs used in the in vivo VML model were kept in 0.9% saline at 37°C with 5% CO2 overnight after AGE induction (before implantation).
DMM characterization
Following decellularization, DMMs and control muscles were homogenized in 0.05% Triton X-100 using a bead homogenizer (BeadBugTM Cat No. 31–212, Genesee Scientific, San Diego, CA) at 4000 rpm for three 60-s cycles. The homogenate was centrifuged at 2,000 g for 30 s, and the supernatant was electrophoresed on a 1% agarose gel and stained with ethidium bromide to compare DNA fragmentation between DMM and control whole muscle biopsies. After confirming DNA removal, a sample of DMM was fixed in 10% neutral buffered formalin (VWR, Radnor, PA), processed histologically, and stained with Masson’s trichrome and Hoechst stains (Sigma-Aldrich, St. Louis, MO). Masson’s trichrome was used to differentiate between collagen and muscle morphology, while Hoechst stain assessed DNA presence.
VML surgery
SAS Sprague-Dawley rats (250–300 g) were obtained from Charles River and divided into four groups: sham, empty defect, DMM (DMM + 0 mM D-ribose), and AGE (DMM + 250 mM D-ribose), with eight rats per group. The rats were provided with standard pellets, water, and environmental enrichment, and housed individually. All surgical procedures were performed under a VCU-approved protocol (IACUC No. AD10000675). VML surgeries were performed as previously described. 3 Briefly, a 1.5 cm × 1 cm defect was cut in the lateral gastrocnemius. Sham surgeries involved the same procedure without muscle removal. In grafted groups, the DMM or AGE-DMM was aligned with the muscle and sutured to the gastrocnemius using horizontal mattress knots at the proximal and distal ends. The biceps femoris was sutured, and the skin was stapled. All but one AGE animal survived. After 8 weeks, muscle force testing was performed using the Aurora Scientific 1305A 3-in-1 Whole Animal System (Aurora Scientific, Aurora, ON, Canada). 11 Animals were euthanized by cervical dislocation under anesthesia after testing.
Tissue processing
The gastrocnemius muscles (right and left) were isolated, with care taken to preserve any fibrosis or remaining DMM material. Gross morphological images were taken with a surgical ruler. The right gastrocnemius muscles were bisected cross-sectionally, with the proximal half used for histology and the distal half for AGE and protein analysis. To ensure adequate penetration during formalin and dehydration steps, the proximal half was cut into 2–3 mm thick sections at the center and edge (25% of the distance between the proximal and distal sutures) of the injury. These sections were fixed in 10% neutral buffered formalin. The distal half was frozen at −80°C until homogenization.
AGE analysis
After AGE induction, 30 mg of minced DMM was homogenized in 400 µL 1 × PBS using a bead homogenizer at 4000 rpm for five 60-s cycles, with tubes kept on ice for at least 5 min between runs. Collagenase type III (STEMCELL Technologies Inc., Vancouver, Canada) (100 µL, 4000 U/mL in 1 × PBS) was added to the homogenate and incubated at 37°C for 1 h. EDTA (Sigma-Aldrich, St. Louis, MO) (0.5 M, 10 µL) was then added to quench the digest, followed by centrifugation at 13,000 rpm for 3 min.
For AGE and protein analysis, 10 mg of the injury site tissue was minced and homogenized in denaturing buffer. The homogenization was performed with a bead homogenizer at 4000 rpm for twenty 60-s cycles, with tubes kept on ice for at least 5 min between runs. The lysates were analyzed using an AGE ELISA (STA-817, Cell Biolabs, San Diego, CA) and normalized with the Pierce™ BCA Protein Assay Kit and hydroxyproline assays (Chondrex, Woodinville, WA). Lysates were hydrolyzed with 5N HCl for 24 h at 120°C before hydroxyproline assay analysis.
Histology
The center and edge of the gastrocnemius muscle were removed from the fixative after 24 h, dehydrated with increasing concentrations of ethanol followed by xylene, and embedded in paraffin blocks. Microtome sections (5 μm) were taken, placed on slides, deparaffinized with xylene, rehydrated, and stained with Masson’s trichrome using Wiegert’s hematoxylin, Biebrich’s scarlet-acid Fuchsia, and aniline blue (Sigma-Aldrich, St. Louis, MO). Coverslips were mounted with xylene-based media and allowed to dry before imaging. Stitched images of the entire histological section were taken using a 10× objective.
Histomorphometry
Histomorphometry was performed using Zen Pro software (Carl Zeiss Meditec AG, Jena, Germany). Healthy muscle from sham-operated animals served as a positive control. Masson’s trichrome staining was used to identify muscle fibers (red), collagen (blue), and nuclei (dark purple). The injury border was conservatively determined by overlaying the contralateral muscle belly outline onto the injured muscle. The area of total muscle, healthy muscle (morphologically similar to sham), regenerative muscle (increased collagen, centrally-located nuclei, and decreased fiber diameter), collagen (blue stain), and fat was quantified manually using the area selection tool in the Zen Blue Pro software. Muscle fiber diameter was measured using the minimum Feret diameter method, whereby the minimum diameter was measured manually using the line tool in the Zen Blue Pro software.
Protein analysis
Thirty milligrams of muscle biopsies were homogenized in NP-40 lysis buffer (BP-119, Boston BioProducts, Ashland, MA) with a protease inhibitor cocktail and 25 mM NaF using a bead homogenizer at 4000 rpm for five 60-s cycles. The homogenate was centrifuged at 9703 rpm (10,000× g) for 10 min, and the lysate was used for dot blotting according to the manufacturer’s protocol. Target proteins were normalized by total protein signal and included RAGE, phosphorylated p38, Pax7, MyoD, Myf5, myogenin (MyoG), neonatal myosin, and myosin heavy chains using secondary antibodies to identify the targets (926-68073, 926-32210, and 926-32211, LI-COR Biosciences, Lincoln, NE).
Mass spectrometry analysis
Three random samples per group (Sham, DMM, and AGE) were selected, and 40 µL of protein isolate were sent to UVA’s mass spectrometry core. The samples were solubilized using a bead mill homogenizer and stainless steel balls, followed by acetone precipitation. The protein pellet was resolubilized, reduced with DTT, alkylated with iodoacetamide, and digested overnight with trypsin. The samples were cleaned and analyzed using a Thermo Orbitrap Exploris 480 mass spectrometer with an Easy Spray ion source. The data were analyzed by database searching using the Sequest algorithm against the Uniprot Rat database.
Statistical analysis
Each variable was tested using N = 8 independent animals unless specified. Data are presented as mean ± SEM, with analysis done using GraphPad Prism 6.0. Outliers were removed according to Grubb’s test (α = 0.05). Analysis comparing two groups was performed by unpaired Student’s t-test, while one-way ANOVA with Tukey’s post hoc test was used for comparisons involving more than two groups. Proteomic data were analyzed with ANOVA followed by Benjamini-Hochberg correction. A Fisher’s exact test was used to compare AGE and DMM groups to each other. A p value <0.05 was considered significant.
Results
Decellularization
Masson’s trichrome stain confirmed a collagenous matrix in DMM with no visible nuclei (Supplementary Fig. S1A–B, Fig. 1A). Hoechst staining showed no stained nuclei at all magnifications, indicating the absence of DNA (Fig. 1B). Agarose gel electrophoresis of DMM, stained with ethidium bromide, revealed no DNA larger than 100 bp compared to the whole muscle control (Fig. 1C), confirming undetectable DNA levels. AGEs were successfully induced in DMM at 250 mM, with significantly more AGEs per hydroxyproline compared to 0 and 100 mM (Fig. 1D). In addition, AGE induction at 250 mM led to reduced soluble hydroxyproline compared to 0 mM, with a similar reduction observed at 100 mM (Fig. 1E). Although total AGEs (without normalization) increased significantly from 0 to 250 mM, wet weight remained constant across D-ribose treatments (Supplementary Fig. S2A–B). AGEs per wet weight increased dose-dependently with D-ribose, with 100 mM higher than 0 mM and 250 mM higher than 0 and 100 mM (Supplementary Fig. S2C). Protein data followed the hydroxyproline trend, with reduced protein levels at 100 and 250 mM (Supplementary Fig. S2D). AGEs per protein increased only at 250 mM compared to 0 and 100 mM (Supplementary Fig. S2E).
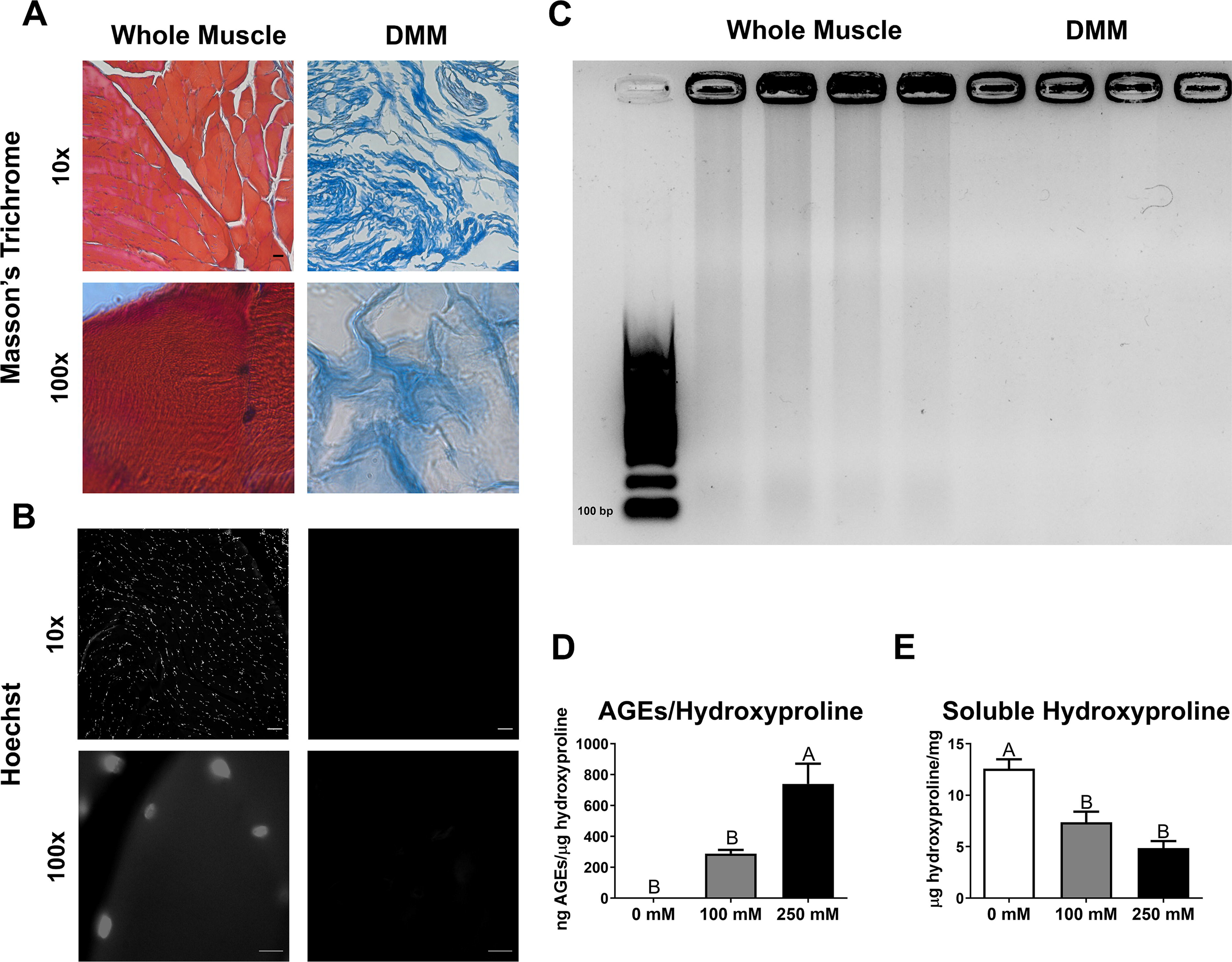
Decellularizing whole muscle into DMM produced a collagenous scaffold devoid of nuclear and cytoplasmic elements, allowing for AGE formation. A–B) Masson’s Trichrome staining (red = muscle, blue = collagen, dark purple = nuclei)
Effect of AGEs on muscle force
Over 8 weeks, animal body weights increased from 300 to 400 grams (Supplementary Fig. S3). Gross morphology of the gastrocnemius after 8 weeks revealed a volumetric deficit in the empty group (Fig. 2A). Muscle weight and width decreased with VML injury, and DMM treatment with and without AGEs did not restore these parameters (Fig. 2B and C). Interestingly, the AGE group had a larger width than the empty group, though not as large as the sham (Fig. 2C). Maximum tetanic force was reduced in empty defects compared to sham, validating our VML model (Fig. 2D). However, maximum tetanic force did not differ between DMM and AGE-treated injuries compared to sham and empty groups. The maximum rate of relaxation decreased in the empty group, with no significant differences in other force-time analysis outcomes (Supplementary Fig. S4). The graft area was larger in the AGE group compared to DMM (Fig. 2E). Muscle biopsies from the injury site showed elevated soluble hydroxyproline in DMM compared to AGE, inversely correlated with collagen cross-linking (Fig. 2F). However, AGEs per hydroxyproline did not differ significantly between AGE groups and DMM, despite an elevated mean (Fig. 2G).
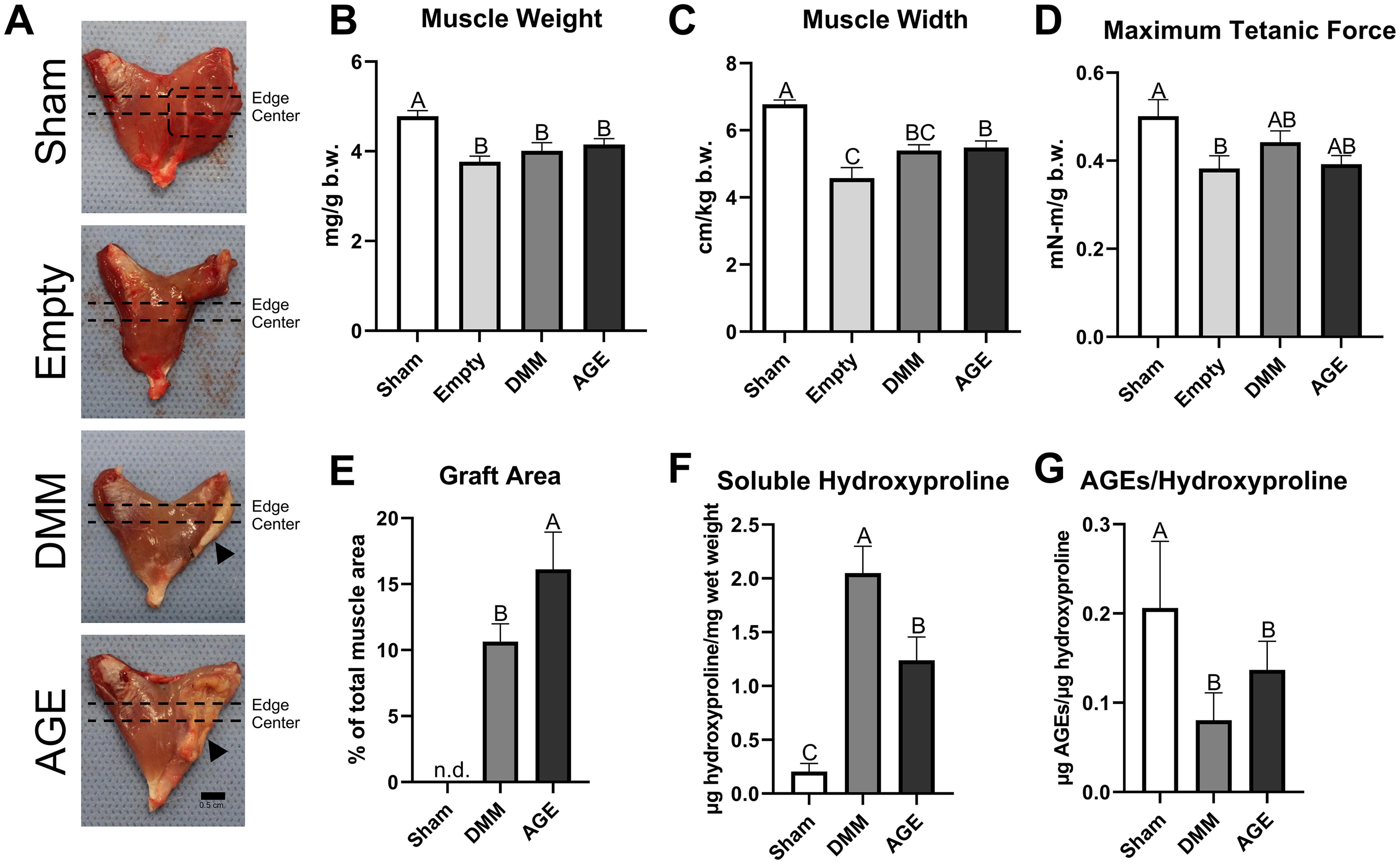
VML injury led to significant muscle mass and force deficits, with AGEs increasing graft area and collagen cross-linking.
Histological assessment of VML-treated injuries
Masson’s trichrome staining indicated increased collagen levels (blue staining) in DMM and AGE groups, with evidence of muscle fiber regeneration (red staining); DMM showed qualitatively healthier muscle compared to AGE at the center (Fig. 3A). Histomorphometry revealed that total muscle area at the edge and center was lower in DMM and AGE compared to sham (Fig. 3C). Total muscle fiber number was reduced at the edge in AGE only compared to sham, while both AGE and DMM had reduced total muscle fiber numbers compared to sham at the center (Fig. 3D). There was a reduction in total muscle fiber diameter in DMM and AGE at the edge only (Fig. 3E). AGE-treated animals exhibited more collagen staining compared to DMM and sham at both the edge and center (Fig. 3F). DMM also had more collagen area than sham at both locations (Fig. 3F). Mature muscle area was reduced in DMM and AGE relative to sham at the edge and center (Fig. 3G). Mature muscle fiber number was reduced in DMM and AGE at both histological sites (Fig. 3H), and mature fiber diameter was reduced in AGE at the center compared to the other groups (Fig. 3I). Fat content was higher in DMM compared to sham and AGE at the edge and center of the defect (Fig. 3J). DMM and AGE animals displayed more de novo muscle at the edge compared to sham, indicating active regeneration (Fig. 3K). Regeneration was increased in DMM compared to sham at the center (Fig. 3K). Regenerative muscle fiber number and diameter increased in DMM compared to sham at the edge and center (Fig. 3L and M). Frequency distributions of muscle fiber diameters showed similar group differences (Supplementary Fig. S5).
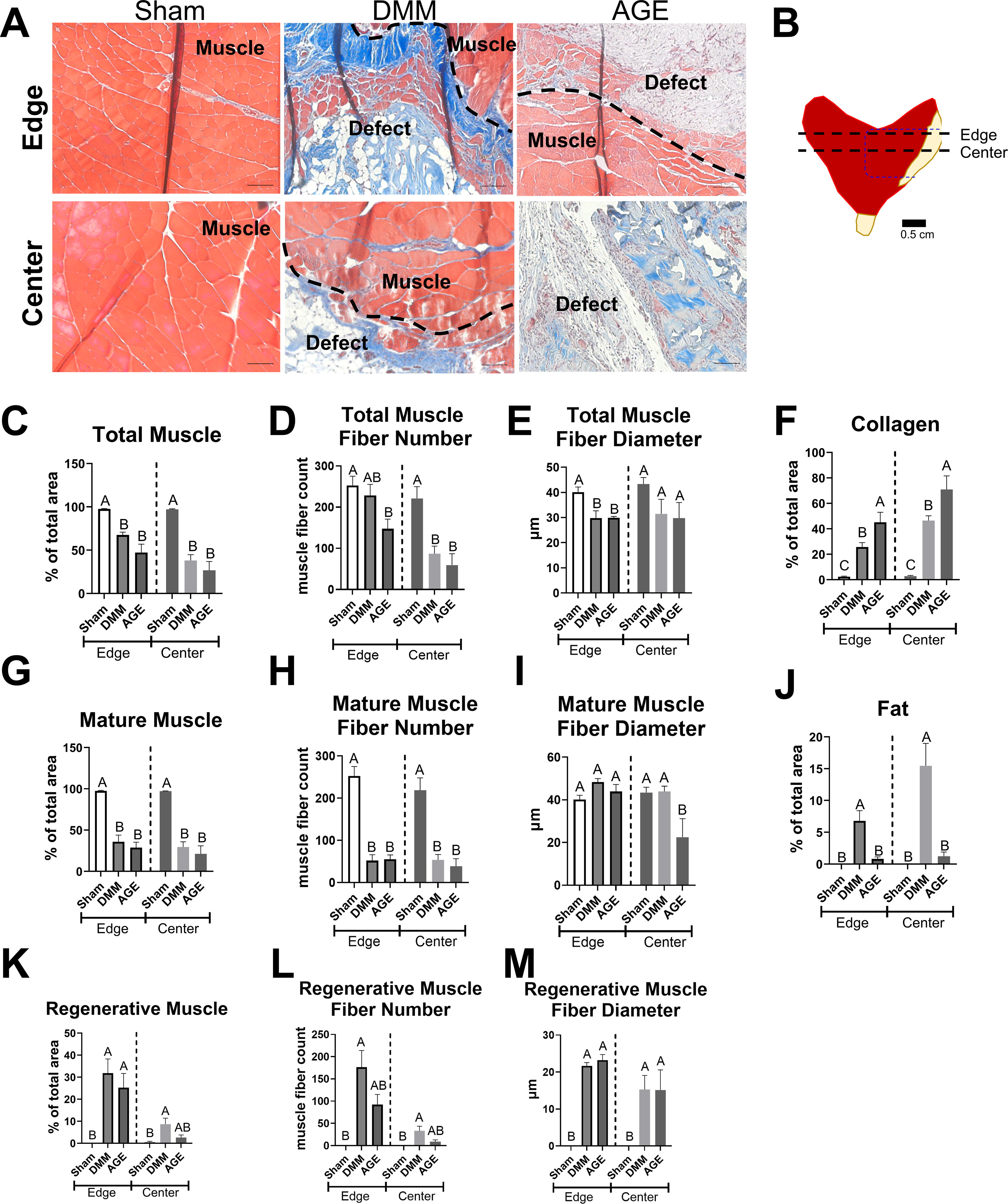
Histological analysis of cross-sections: AGE group shows increased collagen and reduced regeneration compared to DMM
Protein expression analysis of VML treatments
Muscle differentiation and maturation markers were measured in sham, DMM, and AGE groups to validate histological data. Early myogenic markers Pax7 and MyoD were higher in DMM and AGE compared to sham (Fig. 4A and B), with AGE muscles showing more Myf5 compared to DMM and sham (Fig. 4C). MyoG was elevated in DMM and AGE compared to sham (Fig. 4D). The MyoG/Myf5 ratio, positively correlated with myogenic differentiation, was increased in the DMM group compared to AGE (Fig. 4E). Neonatal myosin, fast-twitch MyHC, and slow-twitch MyHC were elevated in grafted groups relative to sham, but AGEs did not differ compared to DMM (Fig. 4F–H). Total RAGE was highest in sham compared to VML-treated injuries, 11 and significantly elevated due to AGEs (Fig. 4I). Phosphorylated p38, a downstream member of the RAGE pathway, was unchanged between DMM and AGE (Fig. 4J).

AGE-induced changes in muscle differentiation and RAGE levels.
Proteomic analysis of VML treatments
Mass spectrometry identified 1994 unique proteins, showing a unique VML injury signature (Fig. 5A). Among treatment and control groups, 38 proteins were unique to the AGE group, compared to 9 in DMM. A Fisher’s exact test identified 111 significant proteins among the 210 shared between DMM and AGE. A Volcano plot displayed 58 proteins upregulated and 53 downregulated in the AGE group (Fig. 5C). Among the 15 muscle-specific proteins identified, 13 were significantly downregulated by AGEs (Fig. 5D). When analyzing significant proteins from DMM or AGE across all groups, GO pathway matching revealed increased immune system processes and decreased extracellular region proteins in the AGE group compared to DMM (Fig. 5E).

Mass spectrometry reveals distinct protein expression in AGE and DMM.
Further analysis with Fisher’s exact test revealed 11 proteins significantly regulated by AGEs under the immune system process umbrella, with 8 increased. In addition, 20 proteins in the extracellular region category were regulated by AGEs, with 15 downregulated (Fig. 6A and B). Notably, immune-associated proteins such as Ptprc, Coro1a, C6 protein, Actr2, Cd14, Cd74, Ndrg1, Sgpl1, and Myh9 were upregulated by AGEs (Fig. 6C–E, I–K, and O–Q). Extracellular-associated proteins such as Col1a2, adipocyte enhancer-binding protein 1 (Aebp1), Lamb2, Comp, Olfml1, Fmod, Clec11a, Thbs4, and Carboxylesterase 1D (Ces1d) were reduced by AGEs (Fig. 6F–H, L–N, and R–T).
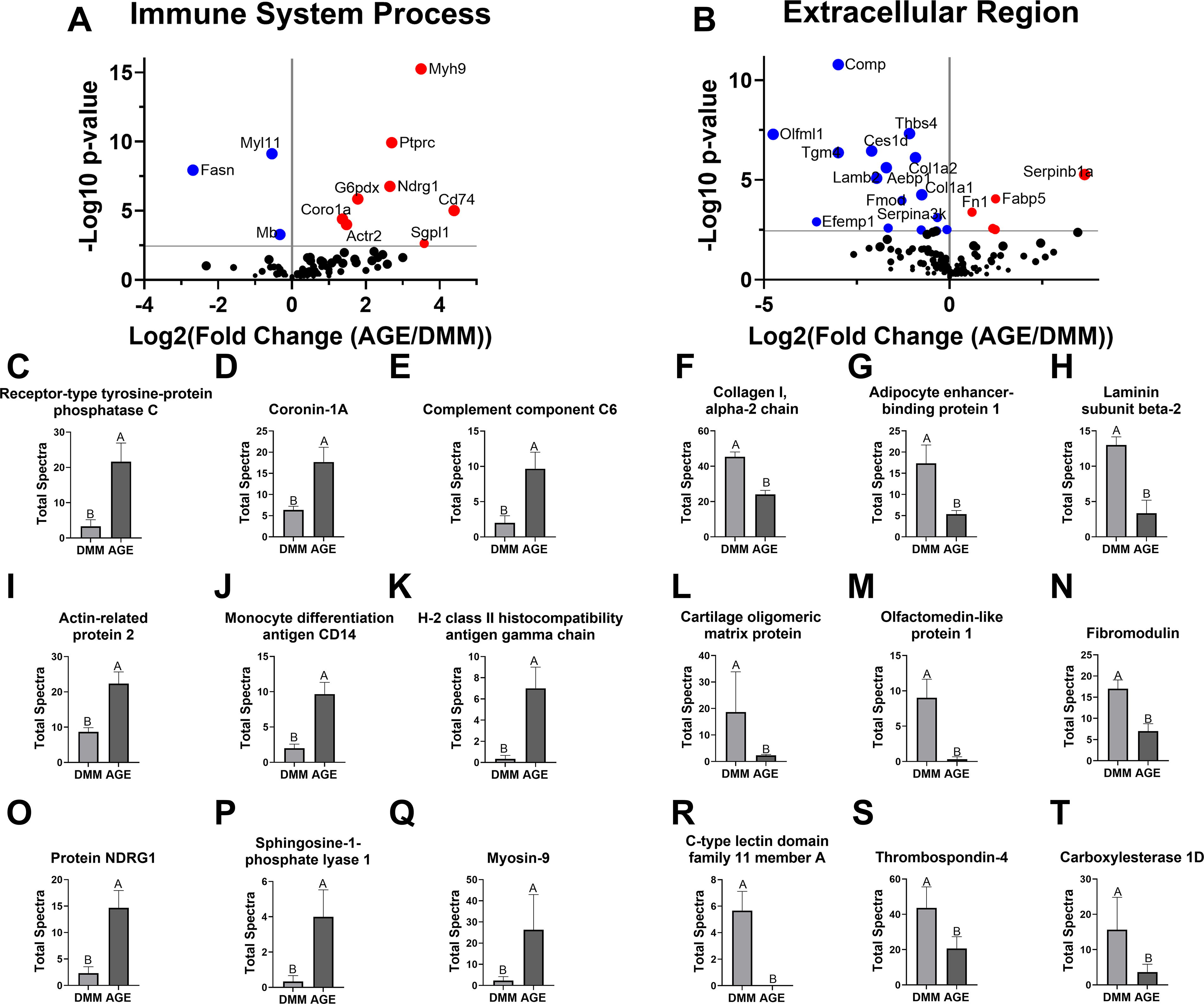
Immune system and extracellular region proteins affected by AGEs.
Analysis of AGE and RAGE-specific proteins showed an increase in Galectin-3, a noncanonical AGE receptor, in the AGE group (Fig. 7A). Platelet glycoprotein 4 and Scavenger receptor class B member 1, other noncanonical AGE receptors, were detected but did not significantly differ with AGE presence (Fig. 7E and I). Analysis of RAGE-pathway-related proteins revealed higher levels of P38α due to AGEs, though not statistically significant (Fig. 7B). No significant differences were detected in Erk1, but osteopontin increased with AGEs (Fig. 7F and J). Among RAGE ligands, only Protein S100-A4 showed a significant increase due to AGEs (Fig. 7C and D, G and H, and K and L).
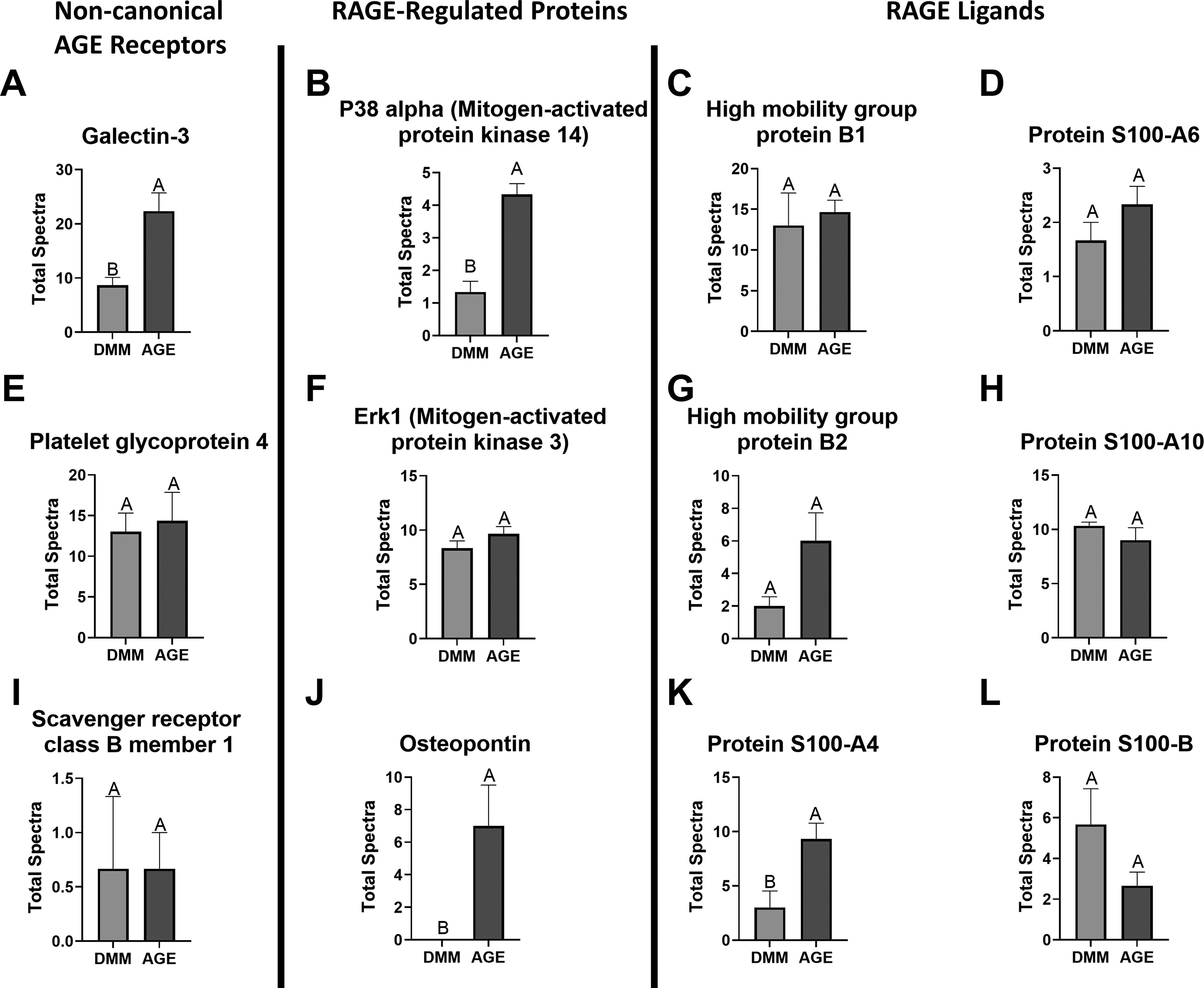
AGE and RAGE-specific protein analysis.
Discussion
The goal of this study was to investigate the role of glycations in DMM-mediated regeneration. Previously, AGEs have been measured in decellularized human muscle, retained in decellularized adipose tissue after whole tissue AGE induction, and accumulated in cardiac and skeletal muscle in old age.6,15–17 We developed an aged DMM model by inducing AGE cross-links in young DMM, resulting in a higher number of collagen cross-links compared to control DMM. The increased AGE levels per hydroxyproline (0.8 µg AGEs/µg hydroxyproline) were similar to those seen in aged murine DMM (0.5 µg AGEs/µg hydroxyproline). 6 However, a limitation is that it is not known whether aged human DMM contains similar AGE levels. This model allows us to study the effects of AGEs on regeneration without other confounding aging factors such as increased total collagen, laminin, decreased fibronectin, and ECM growth factors.6,18–20
We selected a challenging 1.5 × 1 cm full-thickness gastrocnemius defect in Sprague Dawley rats as our VML model, previously characterized by our group to produce a sufficient deficit to test DMM’s regenerative capacity.3,10,11 We validated our model by demonstrating that muscle weight, width, and tetanic force were all reduced in untreated animals (empty defect). As shown in previous studies, DMM treatment partially recovered tetanic force, and AGEs did not negatively impact muscle force. This may be due to functional fibrosis that transmits muscle force, comparable to nonglycated DMM. 21 However, our model’s limitation is the measurement of all posterior crural muscles rather than isolating the lateral head of the gastrocnemius, likely contributing to overall force output.
AGEs regulate collagen degradation and deposition, preventing enzymatic breakdown through cross-linking and amplifying the immune system’s proinflammatory response, leading to fibrosis.22–25 Supporting this, more collagen was measured in AGE-treated DMM compared to nonglycated DMM, with increases of 45% at the edge and up to 71% in the center. Collagen synthesis by fibroblasts, fibroadipogenic progenitor cells (FAPs), and myoblasts is altered by AGE cross-links, promoting a myofibroblast phenotype via TGF-β signaling.26,27 This suggests that AGE-treated DMM may influence fibroblasts in our model. 28
FAPs, a mesenchymal-like cell population, can differentiate into either adipogenic, myofibroblast, or MuSC lineages to support muscle regeneration, increase fat, or fibrosis.28,29 Interestingly, total fat was highest in DMM, while AGE-treated DMM suppressed fat formation near sham levels. Excess fat levels in DMM may play a role in the regeneration process by providing a source of stem cells, however, more study is needed to confirm this hypothesis. 28 Previous studies indicated that substrate stiffness increased α-smooth muscle actin, enhancing myofibroblast differentiation in FAPs. 30 AGEs, which stiffen the ECM, 12 could be pushing FAPs toward a more fibrotic phenotype, but further research is needed to clarify if AGEs are preventing FAPs from following a MuSC differentiation cascade.
Muscle regeneration relies on ECM, MuSCs, fibroblasts, FAPs, and macrophages. Understanding how AGE cross-links in muscle ECM regulate these cell populations is crucial for developing regenerative bio-scaffolds.5,31 We found that AGEs reduced the number of de novo muscle fibers within the injury site during regeneration compared to DMM. Although Feret diameter in regenerating fibers was similar between AGE and DMM-treated injuries, suggesting suppression of muscle fiber formation by AGEs. This leads to an important question: do AGE cross-links exert a unique effect on muscle regeneration different from soluble AGEs? Current studies have focused on soluble AGEs, not ECM-based cross-links, which are more challenging to deliver in vivo. Further, mature muscle fiber diameter was on average smaller in AGE treatments compared to DMM, with DMM fibers similar to sham, implicating AGEs in age-associated sarcopenia.32–34 Since AGE cross-links elicit altered mechanical signaling and direct receptor signaling, a hybrid model is necessary to explain AGE-dependent aging of the skeletal muscle regenerative niche. Notably, our prior research shows that while AGEs stiffen collagen matrices, RAGE signaling alone can reduce muscle fiber formation in vitro. This suggests that signaling, independent of mechanical changes, may account for the observed effects. 12 However, further research is needed to confirm these findings in vivo, and whether altered passive forces may impact muscle regeneration.
Muscle regeneration is tightly regulated by MuSC activation and differentiation.35,36 In adults, MuSCs primarily express Pax7 when quiescent, but upon activation, they express Myf5 and MyoD. 37 While Myf5 plays a role in myoblast proliferation, chronic muscle injury and disease environments upregulate Myf5,38,39 leading to MuSC over-activation and limiting self-renewal. This process also occurs with aging. 40 In our model, Pax7 and MyoD levels were unchanged between DMM and AGE treatments, but AGEs increased Myf5 levels. Recent work from our lab observed increased Myf5 in myoblast cultures grown on AGE scaffolds, 12 indicating that AGEs stimulate Myf5, though the mechanisms remain unclear.
The receptor for AGEs (RAGE) is involved in multiple cell types during myogenesis, including myoblasts, fibroblasts, macrophages, and Schwann cells.41–44 Despite the absence of AGE levels in explanted injury sites, RAGE expression was detected in the AGE group but barely in DMM. Unsurprisingly, RAGE is present at baseline levels in sham muscle due to its expression in fibroblasts, macrophages, and other cell types. Prior research showed that RAGE protein was suppressed in VML injuries compared to sham, suggesting that AGE delivery had a tangible effect on RAGE and may still be present at 8 weeks. The increased RAGE in AGE-treated VML potentially indicates an AGE/RAGE axis, with the cells associated with this RAGE expression yet to be determined. RAGE is expressed in activated myoblasts, and RAGE ligands, S100b and HMGB1, exert proliferative and promyogenic effects via RAGE-P38-MyoG signaling.45–47
To perform a more sensitive and unbiased analysis of the proteomic profile produced by AGE delivery, we used mass spectrometry. As expected, DMM and AGE-treated muscle had a significantly different proteomic profile compared to sham, supporting the idea that our challenging VML model was not yet resolved at 8 weeks. AGEs specifically suppressed muscle-specific proteins (Myh3 and Myh8), supporting histological reductions in regenerative muscle observed in the AGE group. In addition, we explored GO terminology under the biological process and cellular component categories to capture AGE-specific regulation. Interestingly, the immune system response was elevated, and the extracellular region was negatively impacted. We elected to dive deeper into the proteins related to the immune system and extracellular space as AGEs are known to have a profound impact on these wound-healing processes. 48
Several immune-specific proteins were clearly upregulated by AGEs, further supporting a proinflammatory environment. Major innate immune system markers included those for macrophages (Cd14, Actr2, and Cd74) and mast cells (NDRG).49–53 Macrophages are essential in skeletal muscle regeneration but strongly activated in VML injuries, contributing to fibrosis. 31 Furthermore, markers of adaptive immune cells known to play a role in skeletal muscle regeneration, such as the B and T cell markers PTPRC, Coronin-1A, and the T-cell promoting protein Sphingosine-1-phosphate lyase 1, were increased by AGEs.54–56 We previously showed that adaptive immunity regulates skeletal muscle regeneration by comparing immunocompromised to immuno-intact rats. 10 Finally, myosin-9 was increased due to AGEs, a marker of immune cell migration. 57 Chronic and elevated inflammation limits muscle regeneration and plays a critical role in muscle atrophy, and our proteomic data are supported by this prior research. 58
The ECM plays a complex spatiotemporal regulatory role in muscle repair and regeneration, and AGEs negatively regulated ECM production. Reduced collagen type I indicated that while overall collagen levels were elevated, collagen synthesis was suppressed. Prior literature suggested that AGEs disrupt ECM synthesis in fibroblasts, which could explain the reduction in fibrotic factors. 59 In addition, VML injuries in aged animals produce less collagen, and the reductions in ECM components we measured could indicate that AGEs promote an aged regeneration phenotype. 60 AEBP1 and cartilage oligomeric matrix protein, which were reduced in the AGE group, are markers of fibrosis in nonmuscle tissue, and their role in skeletal muscle remains unclear.61,62 Carboxylesterase 1D, an enzyme involved in lipid metabolism and inflammation, was reduced by AGEs, further supporting the idea that AGEs enhanced the inflammatory response. 63 Laminin, osteolectin, fibromodulin, and thrombospondin-4, all known proregenerative ECM proteins, were detected at lower levels in the AGE group, which could be associated with the reduced number of regenerating muscle fibers and the reduced fiber diameter observed.64–68
Aside from RAGE, galectin-3 acts as a separate extracellular receptor for AGEs, present on most innate immune cells, playing an important role in the immune response, and associated with musculoskeletal diseases.69,70 While endogenous galectin-3 also plays a role in normal skeletal muscle regeneration, its levels in AGE-treated muscle were more than double that of DMM, suggesting a role beyond repair or regeneration. 71 However, more study is needed to discern which cell types are responsible for the increased galectin-3. We next investigated RAGE-pathway members to determine the extent of the AGE/RAGE axis involvement. We detected P38α and Erk1; however, they were not significantly regulated by AGEs. AGEs enhanced osteopontin levels, which is regulated by RAGE signaling and associated with increased fibrosis. 72 Osteopontin increases in skeletal muscle with aging and is partly responsible for age-related reductions in skeletal muscle regeneration. 73 We also investigated known RAGE ligands, finding several. Interestingly, only S100-A4 was increased by AGEs, suggesting that AGEs could promote a positive RAGE signaling feedback loop with S100-A4. 74
Conclusion
The presence of AGEs in DMM promotes a fibrotic environment and disrupts myogenesis. AGE cross-linked DMM showed reduced performance in a challenging VML model, with increased fibrosis and collagen cross-linking in the injury, along with elevated RAGE and Myf5 levels, indicating MuSC dysregulation. Muscle regeneration and specific ECM remodeling factors were diminished by AGEs, while inflammation was heightened. Our findings suggest that AGE accumulation in muscle ECM from older donors may reduce efficacy compared to DMM from younger donors. Further research is needed to explore strategies for mitigating the effects of AGEs.
Footnotes
Acknowledgments
Ethics Statement
We confirm that we have read the Journal’s position on issues involved in ethical publication and affirm that this report is consistent with those guidelines.
Authors’ Contributions
L.C.O., M.J.M., and Z.S.: Conceptualization. L.C.O., A.Y.J., E.S.C., B.L., and D.J.C.: Data curation. L.C.O., A.Y.J., E.S.C., B.L., and S.E.P.: Formal analysis. M.J.M.: Funding acquisition. L.C.O., M.J.M., A.Y.J., and D.J.C.: Investigation. L.C.O., E.S.C., and D.J.C.: Methodology. M.J.M. and Z.S.: Project administration. M.J.M., Z.S., and D.J.C.: Supervision. L.C.O., A.Y.J., and B.L.: Validation. L.C.O. and M.J.M.: Writing—original draft. L.C.O., A.Y.J., E.S.C., S.E.P., B.L., D.J.C., Z.S., and M.J.M.: Writing—review and editing.
Disclosure Statement
The authors have no conflicts of interest or financial conflicts of interest to disclose.
Funding Information
This work was supported by the Musculoskeletal Transplant Foundation Junior Investigator Award; and the VCU Commercialization Award.