
Abstract
The activation of chondrogenic progenitor cells (CPCs) in articular cartilage during a traumatic injury is vital for cartilage regeneration. Although our understanding of the mechanisms underlying CPC chondrogenic activation remains incomplete, there is evidence that exosomal microRNAs (miRNAs or miRs) are involved in tissue healing due to their regulating role of posttranscriptional gene expressions. In this study, we profiled enriched and differential expression of miRNAs in exosomes derived from bovine joint cells (CPCs, chondrocytes, and synoviocytes) via Next Generation Sequencing analysis and validated the potential therapeutic effects of candidate exosomal miRNAs for cartilage regeneration. For CPC-based cartilage regeneration, we tested the impact of administering miR-107, miR-140, and miR-148a on CPCs because we found that these miRNAs were highly and differentially expressed in chondrocytes-derived exosomes (CC-Exo). We found that: (1) miR-140 induced chondrogenic gene expression including SRY-box transcription factor 9, collagen type 2A1, and aggrecan, and (2) miR-107 suppressed catabolic gene expression including matrix metalloproteinase 3, a disintegrin and metalloproteinase with thrombospondin motifs 5, and nitric oxide synthase 2. Our findings indicate that transfection of CPCs with specific chondrogenic miRNAs present in CC-Exo have the potential to promote CPC-based cartilage regeneration and could be an important component of posttraumatic osteoarthritis prevention.
Impact Statement
Chondrocytes, chondrogenic progenitor cells (CPCs), and synoviocytes secrete exosomal microRNAs (miRNAs) that contribute to joint health and disease. These miRNAs could also have important implications for improving cartilage repair and regeneration. In this study, we identified candidate miRNAs that were enriched in chondrocytes-derived exosomes and found that these miRNAs induced chondrogenic gene expression or suppressed catabolic gene expression in a CPC monolayer culture system. These findings suggest that miRNA-based cartilage repair strategies could be developed to regenerate damaged and diseased cartilage.
Introduction
Joint injuries can often cause focal cartilage lesions that expand over time and eventually result in cartilage degeneration and posttraumatic osteoarthritis (PTOA). PTOA, the most common disabling condition worldwide, is estimated to account for over 10% of all cases of osteoarthritis (OA).1–3 Thus, early intervention to repair focal lesions is essential to preventing disease progression. 4 Autologous chondrocyte transplantation is a clinical technique used to treat cartilage focal defects.5–7 Besides autologous chondrocytes, mesenchymal stem cells (MSCs) such as bone marrow stromal cells,8,9 adipose-derived stem cells (ADSCs), 10 and umbilical cord MSCs11,12 have also been investigated for their potential of cell-based cartilage regeneration. However, chondrocyte expansion is challenging because these cells quickly lose their chondrogenic phenotype in culture. 13 Similarly, MSC transplantation can result in hypertrophic differentiation and subsequent endochondral ossification. 14 A cell homing strategy that obviates the need for cell isolation and expansion by attracting endogenous host cells into the injured tissue is a promising approach. In the previous study, we found a distinct population of endogenous chondrogenic progenitor cells (CPCs) that spontaneously congregated at injured sites of bovine cartilage and differentiated into chondrocytes with proteoglycan-rich extracellular matrix (ECM) when exposed with exogenous chemotactic (stromal cell-derived factor 1 alpha: SDF-1α) and chondrogenic peptides (transforming growth factor beta 1: TGF-β1, insulin-like growth factor 1: IGF-1).15–17 Thus, a CPC-based therapy that results in CPC activation for homing and differentiation could be a key strategy for cartilage regeneration.
Exosomes are small extracellular vesicles (approximately 30–150 nm) containing deoxyribonucleic acid (DNA), ribonucleic acid (RNA), lipids, and proteins, that facilitate intercellular communication. 18 Scientific and clinical interest in exosomes secreted by stem and progenitor cells has grown rapidly in recent years due to their potential to promote the repair and regeneration of injured tissues.19,20 In particular, exosomal microRNAs (miRNAs or miRs) play an important role in targeting regulatory-related gene messenger RNAs (mRNAs), regulating posttranscriptional gene expressions, contributing cell–cell communication, and cellular processes.21–24 However, some researchers suspect their biological effects due to low copy numbers of miRNAs in exosomes.25,26 Despite the concerns, numerous studies have been identified the potential of MSC-derived exosomes and exosomal miRNAs for cartilage regeneration and OA treatment. 27 For example, miR-140 is an attractive candidate that promotes chondrogenic differentiation, cartilage development and homeostasis, and OA prevention.28,29
In this study, we characterized exosomes derived from different joint cells (CPCs, chondrocytes, and synoviocytes) with the hypothesis that these exosomes will have cell-specific characteristics and cargoes, especially with respect to exosomal miRNAs. We then validated the potential therapeutic effects of the candidate exosomal miRNAs we identified for CPCs-based cartilage regeneration.
Materials and Methods
Joint cell isolation
A total of 4 bovine stifle joints (15–24 months old) were used in this study due to their large size to isolate cells, ready availability from a local abattoir (Bud’s Custom Meats, Riverside, IA, USA), and feasibility for a defect-injury model. The joints with any sign of swelling/inflammation, traumatic injury, and abnormal tissue color change were excluded in this study. Primary chondrocytes and CPCs were isolated from the tibia plateau including the central loaded area of the articular surface, and synoviocytes were isolated from posterior joint capsules. Briefly, finely chopped cartilage fragments were digested overnight in a 0.25% (w/v) collagenase/proteinase (Sigma-Aldrich, St. Louis, MO, USA) cocktail to isolate chondrocytes. For CPC isolation, scratch injuries were created on the surface of osteochondral explants using a sterile 26-gauge needle. 16 After 10–14 days incubation, CPCs were collected by 0.1% trypsin-ethylenediaminetetraacetic acid (EDTA; Thermo Fisher Scientific). The inner membrane of the joint capsule (i.e., intima) was scraped gently with a sterile scalpel immediately after opening the capsule, and synoviocytes were isolated after washing with Hanks’ Balanced Salt Solution (HBSS; Thermo Fisher Scientific, Waltham, MA, USA) and centrifuging. All cells were cultured in Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F-12; Thermo Fisher Scientific) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific), 100 U/mL penicillin-streptomycin (Thermo Fisher Scientific) under hypoxic conditions (5% O2/CO2 at 37°C). Only cells of passage 1 through 4 were used in all experiments.
Exosome isolation
The joint cells were plated in T-75 flasks at a density of 2.1 × 106 cells per flask. At 40–50% confluency, the plates were washed thrice with HBSS, and culture medium was replaced with DMEM/F-12 containing 10% exosome-depleted FBS (System Biosciences, Palo Alto, CA, USA) to avoid the contamination of bovine exosomes present in regular FBS. After 48 h (h), the conditioned media (CM) were collected for exosome isolation. Highly concentrated exosomes were isolated by a modified precipitation method using an ExoQuick-TC® ULTRA extracellular vesicle isolation kit (System Biosciences). Briefly, the CM was concentrated 20–35 folds using 10 kDa Amicon® Ultra Centrifugal Filters (MilliporeSigma, Burlington, MA, USA) and then mixed with 20% (v/v) ExoQuick-TC® solution. Following overnight incubation on ice, the mixture was centrifuged at 3,000 g for 10 minutes (min) and stored in −80°C for further studies.
Characterization of exosomes
The characteristics of exosomes were confirmed by Nanoparticle Tracking Analysis (NTA), Scanning Electron Microscope (SEM), and exosome-specific antibody array. Multilaser NTA (ViewSizer 3000, HORIBA Scientific, Kyoto, Japan) was used to characterize the distribution and size of purified exosomes. 30 Latex beads with a 100 nm diameter (Sigma-Aldrich) were used as a reference. The exosomes stock was diluted in sterile Dulbecco’s Phosphate-Buffered Saline (DPBS, Thermo Fisher Scientific) for vesicle optimal capture and mechanically stirred for three seconds between video captures to ensure even dispersion.
For SEM imaging, silicon wafers (Ted Pella, Redding, CA, USA) were precoated with 1% aqueous poly-l-lysine solution (Thermo Fisher Scientific), dried on a 37°C sterile hot plate for up to an hour, and fixed on SEM aluminum stubs with double-sided carbon tape. A few microliters of well-mixed exosome concentrate diluted with DPBS were added to the surface-treated silicon wafers and air-dried for 15 min under sterile conditions. After coating with 5 µm-thick iridium sputters (Quorum Technologies, Sacramento, CA, USA) to improve surface conductivity, the final product was imaged in a high-resolution SEM (S-4800, Hitachi High-Tech, Ibaraki, Japan).
For exosome identification, 8 exosome markers including CD63, epithelial cell adhesion molecule (EpCAM), annexin A5 (ANXA5), tumor susceptibility gene 101 (TSG101), flotillin-1 (FLOT1), intercellular adhesion molecule 1 (ICAM), apoptosis-linked gene-2-interacting protein X (ALIX), and CD81 were evaluated using an Exo-CheckTM array (System Biosciences). Cis-golgi matrix protein (GM130) as a negative marker was used for cellular contamination during exosome isolation. The exosomes were labeled according to the manufacturer’s protocol, 31 and the membrane was developed with WesternBright Sirius HRP substrate (Advansta, Menlo Park, CA, USA).
Next generation sequencing (NGS) analysis
Isolation of exosomal ribonucleic acids (RNAs)
Two batches (passage 1) of chondrocytes-derived exosomes (CC-Exo), synoviocytes-derived exosomes (SC-Exo), and CPC-derived exosomes (CPC-Exo) were processed for NGS analysis. Exosomes (approximately 1.5 x 1012 particles) were isolated from the CM of a T-75 flask. Exosomal RNAs were extracted using an ExoQuick® Exosome RNA Column Purification kit (System Biosciences). Briefly, supernatant was carefully aspirated to retain only the exosomes pellet, which was then resuspended in 350 µL of lysis buffer and briefly vortexed. After adding 200 µL of 100% ethanol, the samples were vortexed again prior to column-based purification. Initial flow-though and the two washes from spin column were discarded. Up to 30 µL of elution buffer was applied and centrifuged at 2,000 rpm for 2 min to load the membrane with buffer, before increasing it to 13,000 rpm for 1 min to recover exosomal RNAs. The quality and quantity of extracted RNAs were checked with a Bioanalyzer RNA Pico kit (Agilent, Santa Clara, CA, USA). Mean value of total quantities/RNA Integrity Numbers (RINs) were 26.2 ng/2.75, 129.85 ng/9.7, and 66.2 ng/9.5 for CC-Exo, SC-Exo, and CPC-Exo, respectively.
Library generation and sequencing
One ng RNA was used as starting material in a small RNA library prep kit (Qiagen, Germantown, MD, USA). The final loading concentration was 1.45 pM, and no PhiX was added during this process. The dilution ratio used for the adaptors was as follows: 3′: 5× dilution, 5′: 2.5× dilution. Libraries were amplified by polymerase chain reaction (PCR; 22 cycles) and successful libraries were purified by gel extraction to ensure isolation of the correct size. A minimum final concentration of 1 nM was required to move on to the sequencing step. The sequencing was performed on NextSeq (Illumina, San Diego, CA, USA) with single-end reads (75 bp) at an approximate depth of 10–15 million reads per sample.
Bioinformatics analysis of small RNAs
Analysis of raw data was performed by the Bioinformatics Division of the Iowa Institute of Human Genetics according to the previous protocol. 31 A nextflow-based workflow (https://github.com/nf-core/smrnaseq) was used to quantitate known small RNAs. Bovine (Bos taurus, build Btau_5.0.1) genomic sequence (GCA_000003205.6_Btau_5.0.1_genomic.fna) was downloaded from National Center for Biotechnology Information (NCBI). Genome annotation (build accession GCA_000003205.6) and the sequences for both mature and hairpin small RNAs were downloaded from miRBase. The adaptor protocol used was ‘qiaseq’, but the default values were used for all other parameters. To assess read quality and to confirm the presence of the expected adapters before trimming, we used FastQC (version 0.11.9). To trim adapters from the read sequences, Trim Galore! (version 0.6.6) was used with the following parameters: quality Phred score cutoff 20, maximum nucleotide length 40 bp, and minimum length 17 bp. FastQC was run again after trimming to confirm the efficient removal of adapter sequences. Bowtie (version 1.3.0) was used for aligning reads to small RNA sequences with the “best” parameter set to true to ensure that only alignments considered to be of the highest quality were reported. Read counts of all miRNAs with nonzero expression were imported into R (version 3.5.2) and normalized using DESeq2 (version 1.22.2). For visualization purposes, the data was log-transformed with the rlog function and Principal Components Analysis (PCA) was used to visualize the clustering of samples. To determine differential expression, a linear model incorporating all the experimental factors was created and Wald tests were used to compute statistical metrics. A small RNA was considered to have a statistically significant change in expression if the False Discovery Rate (FDR) was less than 10% and the absolute log2 fold-change was greater than 1. Results from the differential expression analysis were visualized using heatmaps created with the pHeatmap (version 1.0.12). To identify novel miRNAs, sequence read files (fastq) from samples in the same group were first concatenated together then ran through the same nf-core smrnaseq workflow as above with the mirDeep2 program (version 2.0.1.2) used for novel miRNA identification. The results from mirDeep2 were filtered to include only potential novel miRNAs with miRDeep2 scores greater than or equal to 4 (higher quality).
miRNA transfection
The pre-customed miRNA mimics for miR-107, miR-140, miR-148a, and a negative control were purchased from Thermo Fisher Scientific (mirVanaTM). The miRNAs at a final concentration of 50 nM were transfected in CPCs using a Lipofectamine RNAiMAX kit (Thermo Fisher Scientific). The CPCs (passage 1) were plated in 24-well plates at a density of 5 × 104 cells per well (n = 3) and incubated in serum-free DMEM/F-12 supplemented with 10 µg/mL insulin/5.5 µg/mL transferrin/6.7 ng/mL selenium (ITS-G; Thermo Fisher Scientific), 50 µg/mL l-ascorbic acid (Thermo Fisher Scientific), 100 U/mL penicillin-streptomycin (Thermo Fisher Scientific), and 100 U/mL amphotericin B (Thermo Fisher Scientific).
Gene expression
After 6 days, total RNAs were extracted from CPCs with TRIzol reagent (Life Technologies, Carlsbad, CA, USA) 16 and Direct-zol RNA kit (Zymo Research, Irvine, CA, USA) as the manufacturer’s instructions. A microvolume spectrometer, NanoDrop 2000 (Thermo Fisher Scientific), was used to assess the yield and purification of extracted RNAs. Quantitative reverse transcription PCR (qRT-PCR) was performed with a PrimeTime® One-Step RT-qPCR Master Mix (Integrated DNA Technologies, Coralville, IA, USA) and TaqMan® probes (Applied Biosystems, Waltham, MA, USA) using QuantStudio™ 3 System (Applied Biosystems) as recommended by the manufacturer. Premade probes (Thermo Fisher Scientific) related to chondrogenic markers, SRY-box transcription factor 9 (SOX9; Bt07108872_m1), collagen type 2A1 (COL2A1; Bt03251861_m1), and aggrecan (ACAN; Bt03212186_m1)) and catabolic markers, matrix metalloproteinase 3 (MMP3; Bt04259490_m1), a disintegrin and metalloproteinase with thrombospondin motifs 5 (ADAMTS5; Bt04230785_m1), nitric oxide synthase 2 (NOS2; Bt03249599_m1) were prepared, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Bt03210913_g1) was used as a housekeeping gene. The relative changes in gene expression levels were calculated by a delta-delta-cycle threshold (Ct) method. 32 In order to estimate the stability of housekeeping gene, coefficient of variances (CVs) of Ct values were calculated by dividing the standard deviation of the Ct values by their mean (expressed as a percentage). 33 Scatter plots of gene expression depicting mean values and standard deviation were prepared using GraphPad Prism (Version 10.1.2; San Diego, CA, USA). Data analysis was performed by one-way analysis of variance (ANOVA) with the Tukey posthoc test using SPSS Statistics software (Version 29; IBM, Armonk, NY, USA). Statistical significance was set at p < 0.05.
Results
Characterization of exosomes
In NTA (Fig. 1a), latex beads as a reference control were on average 94.3 ± 1.5 nm in diameter. 50% of these particles (D50) were smaller than 91 nm. All exosomes were monomodal distributed with average size of approximately 70–90 nm, but each type had slightly different particle size and distribution. The average particle diameters of CC-Exo, SC-Exo, and CPC-Exo were 86.6 ± 2.5 nm, 76.3 ± 1.5 nm, and 69.9 ± 2.0 nm, respectively. SEM images provided visual confirmation of spheroid-like morphology (Fig. 1b). In Exo-CheckTM antibody array, exosomes in all groups showed positive expressions of CD63, ANXA5, TSG101, FLOT1, ICAM, ALIX, and CD81 as exosome positive markers, and a negative expression of GM130 as a exosome negative marker (Fig. 1c).

Characterization of exosomes.
miRNA profiles
In NGS analysis, individuals in a Principal Component Analysis (PCA) plot were clustered together in the same groups, indicating unique characteristics of each exosome group (Fig. 2a). We found a total of 382, 568, 464 mature miRNAs in CC-Exo, SC-Exo, and CPC-Exo, respectively. Table 1 shows the list of the 30 most prevalent mature miRNAs. In particular, 3 miRNAs (bta-let-7b: 9.25%, bta-miR-3596: 9.25%, bta-let-7a-5p: 9.09%) in CC-Exo, 1 miRNA (bta-miR-21-5p: 9.74%) in SC-Exo, and 3 miRNAs (bta-let-7a-5p: 13.01%, bta-let-7b: 9.48%, bta-miR-3596: 9.48%) in CPC-Exo represented 9% or more of all miRNAs sequenced in a sample. Among the highly expressed miRNAs, unique and common miRNAs across the 3 kinds of exosomes were visualized in a Venn diagram (Fig. 2a). miRNAs common to all three exosomes sources included bta-let-7a-5p/−7b/−7f/−7i, bta-miR-16a, bta-miR-21-5p, bta-miR-23a/b-3p, bta-miR-24-3p, bta-miR-25, bta-miR-27b, bta-miR-29a, bta-miR-122, bta-miR-125a/b, bta-miR-148a/c, bta-miR-152, bta-miR-199a-3p, bta-miR-221, bta-miR-423-5p, bta-miR-3596, bta-miR-3604, and bta-miR-10174-3p. The miRNAs unique to a given type of exosome were bta-miR-140 (1.22%) and bta-miR-103 (0.48%) in CC-Exo, bta-miR-143 (1.00%) in SC-Exo, and bta-miR-29b (0.46%), bta-miR-26b (0.41%), and bta-miR-99b (0.40%) in CPC-Exo.
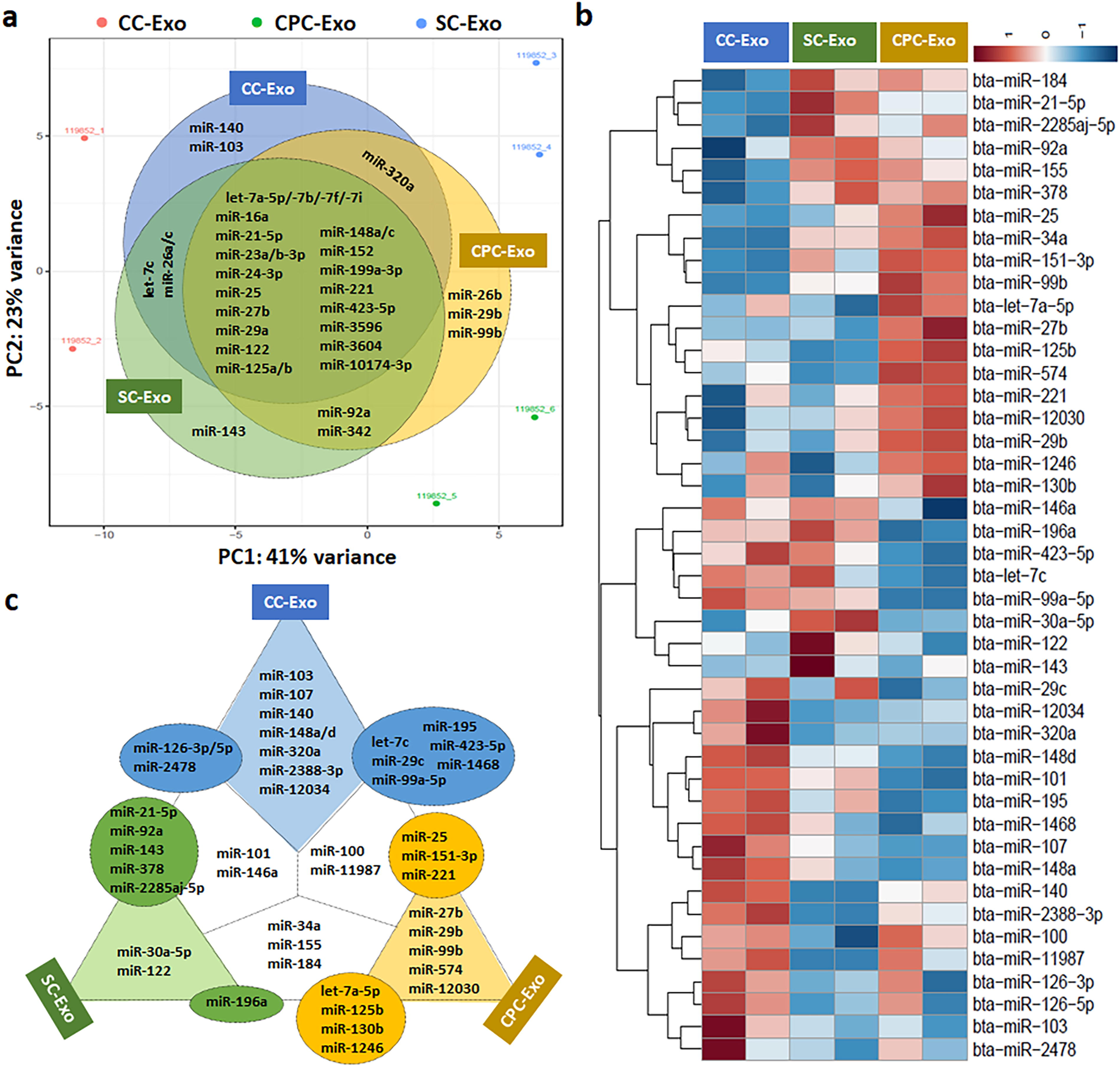
Next Generation Sequencing (NGS) analysis.
List of 30 Most Prevalent Mature microRNAs
bta, Bos taurus; CC-Exo, chondrocytes-derived exosomes; CPC-Exo, chondrogenic progenitor cells-derived exosomes; miR, microRNA; SS-Exo, synoviocytes-derived exosomes; %, percentage of total reads.
All differentially abundant mature miRNAs are listed in Table 2. In the comparison of log2 fold changes, CC-Exo had 2.27- and 5.56-times higher expression of bta-miR-140 than CPC-Exo and SC-Exo, respectively. Similarly, SC-Exo and CPC-Exo had significantly higher expression of bta-miR-30a-5p (2.44-times higher than CC-Exo and 2.83-times higher than CPC-Exo) and bta-miR-99b (2.86-times higher than CC-Exo and 1.34-times higher than SC-Exo), respecively. All lists were visualized by a heatmap with hierarchical clustering (Fig. 2b) and summarized in Figure 2c. For example, 8 miRNAs (bta-miR-103, bta-miR-107, bta-miR-140, bta-miR-148a, bta-miR-148b, bta-miR-320a, bta-miR-2388-3p, and bta-miR-12034) were up-regulated in CC-Exo when compared with both SC-Exo and CPC-Exo, and 3 miRNAs (bta-miR-126-3p, bta-miR-126-5p, and bta-miR-2478) were up-regulated in CC-Exo when compared with SC-Exo (low expression in CPC-Exo) (Fig. 2c).
List of Significantly up/down-Expressed Mature microRNAs (≥ 0.1% Population)
CC-Exo, chondrocytes-derived exosomes; SS-Exo, synoviocytes-derived exosomes; CPC-Exo, chondrogenic progenitor cells-derived exosomes; bta, Bos taurus.
Besides mature miRNAs, we identified 2, 10, and 7 novel miRNAs in CC-Exo, SC-Exo, and CPC-Exo, respectively (Table 3). Especially, 2 novel miRNAs were highly detected with miRDeep2 scores greater than 5,000 in both SC-Exo (GACCCGCGGGCG CTCTCCAGTCCT) and CPC-Exo (GACCCGCGGGCG CTCTCCAGTCCT).
List of Novel microRNAs (miR; ≥ 4.0 miRDeep2 Score)
CC-Exo, chondrocytes-derived exosomes; CPC-Exo, chondrogenic progenitor cells-derived exosomes; SS-Exo, synoviocytes-derived exosomes.
Potential therapeutic application of exosomal miRNAs for cartilage regeneration
As a stable housekeeping gene, GAPDH indicated 4.6% and 4.4% CV in chondrogenic (Fig. 3a) and catabolic markers (Fig. 3b), respectively. Based on NGS data that miR-107, miR-140, and miR-148a were the most differentially expressed miRNA found in CC and CPC exosomes, we evaluated potential therapeutic effects of each miRNA. The qPCR analysis of gene expression in CPCs transfected with miR-140 showed induction of canonical markers of chondrocyte differentiation, SOX9 (3.4-times higher versus control), COL2A1 (47.7-times higher versus control), and ACAN (3.5-times higher versus control), but transfection did not suppress catabolic markers, MMP3 (5.0-times lower versus control), ADAMTS5 (4.8-times lower versus control), and NOS2 (6.7-times lower versus control) (Fig. 3). Conversely, miR-107 transfection suppressed the catabolic markers.

Effect of targeted microRNAs (miRNAs or miR) on chondrogenesis and anticatabolism. Chondrogenic progenitor cells were transfected with miR-107, miR-140, or miR-148a (n = 3). After 6 days, the expressions of chondrogenic
Discussion
Mounting evidence indicates that chemokines,34,35 growth factors, 36 and cytokines 37 can stimulate the cellular activation. Besides these factors, miRNAs as an epigenetic factor play a crucial role in cartilage development, homeostasis, and pathology. 38 miRNAs consisting of approximately 18–25 nucleotides are small, single-stranded, highly conserved noncoding RNA molecules that function in regulating gene expression. Because exosomes contain these miRNAs, analysis of exosomal miRNA profiles was conducted to identify differential expression of miRNAs in joint cells in this study. In NGS analysis, enriched exosomal miRNAs were listed in Table 1, and common and unique miRNAs were summarized in Figure 2b. Among the unique miRNAs, both miR-29b strands were highly expressed in CPC-Exo (Fig. 2a and c). Although we were unable to distinguish between arm/strands (5′ or 3′) within our samples, the distinction has biological significance, as the strands can have different biological functionality and stability. For example, the expression of miR-29b-5p was down-regulated in an anterior cruciate ligament transection (ACLT) rat model. 39 Interestingly, sustained delivery of agomir-29b-5p in self-assembling peptide nanofiber hydrogels enhanced cartilage regeneration. 39 In contrast, miR-140 and miR-107 were highly and differentially expressed in CC-Exo. Several studies reported both prochondrogenic and anti-inflammatory effects of miR-140 which is the most highly detected miRNA in healthy cartilage and down-regulated in OA cartilage.28,40,41 For OA treatment, targeting miR-103a-3p up-regulation inhibited chondrocyte apoptosis and inflammation. 42 Thus, specific miRNAs can be differently expressed depending on the cell type and its function, and engineered modifications of miRNA content in exosomes or miRNA itself can be therapeutically used in tissue engineering and regenerative medicine. 27
In this study, we aimed to evaluate candidate miRNAs for CPC-based cartilage regeneration. Based on gene expression profiles, CPCs significantly over-expressed chemokines (interleukin 8 and C-C motif ligand 2) associating with inflammatory response against cartilage injury, while down-expressed cartilage matrix genes. 43 Our supplementary data could also support the gene expression differences of chondrogenic markers: 5.4-, 2,628.2-, and 5.8-times higher expression of SOX9, COL2A1, and ACAN, respectively, in chondrocytes versus CPCs (Supplementary Fig. S1). Therefore, we focused on differentiated miRNAs between CPC-Exo and CC-Exo (Table 2 and Fig. 2), and selected miR-107,44,45 miR-140,28,41 and miR-148a 46 which showed strong evidence for cartilage regeneration in the previous studies. In gene expression analysis, miR-140 and miR-107 showed predominant effects on anabolic (SOX9, COL2A1, and ACAN) and anticatabolic (MMP3, ADAMTS5, and NOS2) activities for cartilage regeneration, respectively (Fig. 3). On the other hand, CPCs transfected miR-148a induced over-expression of catabolic genes related to cartilage degradation unlike the outcomes of Vonk and colleagues. 43 We assume that the miRNA may affect differently to the target cells (CPCs versus OA chondrocytes). Although we tested only 3 miRNAs, other miRNAs (miR-99a-5p,47,48 miR-103, 49 miR-126-3p, 50 and miR-320a51,52) as well as novel miRNAs (GACCCGCGGGCGCTCTCCAGTCCT and AGCGAATGATTAGAGGTCTTGGG) can also be valuable candidates for cartilage regeneration (Fig. 2 and Table 3).
Because synovium (synovial membrane) is a thin primary barrier that lines the inside of joints, synoviocytes (synovial cells) are closely correlated to the onset and progression of joint disease including pain, inflammation, and cartilage destruction. 53 Therefore, therapeutics targeting synovium and synoviocytes are an attractive strategy for cartilage repair and inflammation inhibition. 54 In particular, exosomes derived from synovial MSCs (SMSCs) suppressed OA development and enhanced cartilage regeneration,55,56 and exosomes derived from miR-140-5p-rich SMSCs promoted cell migration and proliferation without ECM damage. 29 Therefore, the miRNA profiling of SC-Exo can identify enriched and differentiated miRNAs and underlying mechanisms of cellular function. For example, the up-regulation of miR-143, which was detected in SC-Exo (Table 1 and Fig. 2b), in MSC-derived exosomes inhibited the apoptosis of OA cells. 57 Moreover, SC-Exo contained higher levels of miR-92a 58 and miR-122 59 related to prochondrogenesis and anti-inflammation, respectively, compared to CC-Exo and CPC-Exo (Fig. 2a and c). In the future, it will be an interesting approach to compare the miRNA profiles of inflammation-induced synoviocytes to identify the changes of miRNAs contents under pathological condition like synovitis.
In this study, we firstly examined the miRNA profiles of CPC-Exo by comparing with CC-Exo and SC-Exo and identified candidate miRNAs for CPC-based cartilage regeneration. Potential limitations of our research include the confirmation of protein and RNA contamination during exosome isolation, 60 low sample size of NGS and validation of NGS data (Sanger sequencing or qRT-PCR), 61 monolayer culture for chondrogenic induction, dosage-dependent effect of miRNAs, validation of miRNA effects with an antagomir, and targeting only 3 miRNA candidates. However, our findings provide a foundation for the design of exosome-based strategies for cartilage regeneration as well as PTOA prevention.
Conclusions
This study evaluated miRNA profiles of CPC-Exo with the aim to find candidate miRNAs that could be therapeutically beneficial for cartilage regeneration. In the comparison analysis of NGS data, we identified unique mature miRNAs, highly detected in CC-Exo, to be used in targeting of miRNAs via CPC transfection. In particular, miR-140 induced chondrogenic gene expression including SOX9, COL2A1, ACAN, and miR-107 suppressed catabolic gene expression including MMP3, ADAMTS5, and NOS2. These findings suggest that pairing miR-140 and miR-107 would have complimentary effects on cartilage regeneration (Fig. 4).

Overall summary of exosomal microRNA (miRNA or miR), miR-107 and miR-140, for cartilage regeneration. SOX9: SRY-box transcription factor 9, COL2A1: collagen type 2A1, ACAN: aggrecan, MMP3: matrix metalloproteinase 3, ADAMTS5: a disintegrin and metalloproteinase with thrombospondin motifs 5, NOS2: nitric oxide synthase 2.
Footnotes
Acknowledgment
The authors thank Amelia Hurley-Novatny & Hongshuai Li for helping with the exosome antibody array and Abigail Smith & Jaycie Pergler for invaluable administrative support.
Authors’ Contributions
V.G.: Conceptualization, methodology, data curation, and writing-review and editing, visualization. R.H.: Methodology, data curation, and writing—review and editing. H.L.K.: Software, data curation, and writing-review and editing. A.K.S.: Resources, and writing—review and editing. E.A.S.: Writing-review and editing. K.S.: Resources, and writing—review and editing. J.A.M.: Conceptualization, writing—review and editing, and supervision. D.S.: Conceptualization, data curation, writing-original draft, visualization, and supervision.
Author Disclosure Statement
The authors declare no competing interest.
Funding Information
This work was supported by the Department of Orthopedics and Rehabilitation at the University of Iowa.