
Abstract
Cartilage is the first skeletal tissue to be formed during embryogenesis leading to the creation of all mature cartilages and bones, with the exception of the flat bones in the skull. Therefore, errors occurring during the process of chondrogenesis, the formation of cartilage, often lead to severe skeletal malformations such as dysplasias. There are hundreds of skeletal dysplasias, and the molecular genetic etiology of some remains more elusive than of others. Many efforts have aimed at understanding the morphogenetic event of chondrogenesis in normal individuals, of which the main morphogenetic and regulatory mechanisms will be reviewed here. For instance, many signaling molecules that guide chondrogenesis—for example, transforming growth factor-β, bone morphogenetic proteins, fibroblast growth factors, and Wnts, as well as transcriptional regulators such as the Sox family—have already been identified. Moreover, extracellular matrix components also play an important role in this developmental event, as evidenced by the promotion of the chondrogenic potential of chondroprogenitor cells caused by collagen II and proteoglycans like versican. The growing evidence of the elements that control chondrogenesis and the increasing number of different sources of progenitor cells will, hopefully, help to create tissue engineering platforms that could overcome many developmental or degenerative diseases associated with cartilage defects.
Introduction
These regulatory events conduct a set of morphogenetic and phenotypic changes to the MC within the embryonic mesoderm, giving rise to a preskeletal tissue composed mainly of chondroblasts. Here, chondrogenesis is defined as the process through which the MCs differentiate into chondroblasts that subsequently either develop into adult chondrocytes or undergo hypertrophy and apoptosis. 4 In endochondral ossification, a part of this embryonic cartilage subsequently turns into bone. 5
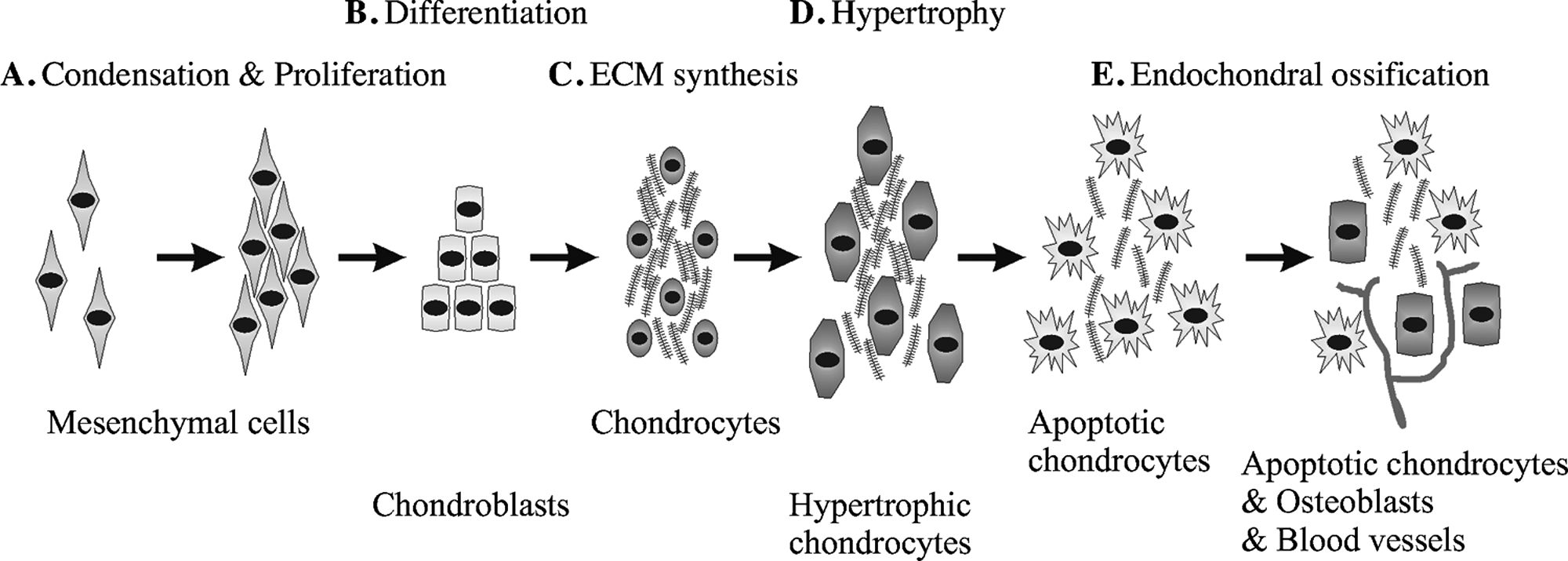
Endochondral ossification can be divided into five main stages (Fig. 1). These stages are clearly defined by specific events:
Commitment of MCs to become cartilage caused by paracrine factors, such as fibroblast growth factor (FGF) and Hedgehog pathways.
Differentiation of the condensed MCs into chondrocytes in the process of which the transcription factor Sox9 plays an essential role in controlling the expression of downstream genes specific for cartilage tissue development.
Rapid division of chondrocytes and production of the cartilage-specific ECM.
Halted proliferation of chondrocytes and a several fold increase in size entering hypertrophy. The composition of the ECM (mainly collagen type X and fibronectin) is also changed, and chondrocytes start to mineralize the environment with calcium salts.
Invasion of blood vessels takes place, and the hypertrophic chondrocytes die by apoptosis. At this stage, osteoblast precursors invade the remodeling tissue and start forming bone using the cartilaginous matrix as template and replacing it by mineralized matrix.
In this review, we will focus on the morphogenetic changes, the regulatory mechanisms, and the interactions that take place during developmental chondrogenesis. In this context, a precise knowledge on the extrinsic factors (environmental) and intrinsic factors (genetic regulation) controlling chondrogenesis can be easily translated into tissue engineering research to obtain more reliable therapeutic products. Therefore, we will place special emphasis on the importance of the ECM during the development of cartilage and its biomechanical function to help recreate similar processes during the development of regenerative platforms. Moreover, we will also summarize recent descriptions of chondroblastic progenitors that are expected to mimic natural chondrogenesis and might play a key role in biomedical research in the near future. Finally, we will discuss some of the novel biomaterials and its applications currently used for cartilage and, eventually, bone repair.
Morphogenetic Changes Associated with Chondrogenesis
Natural chondrogenesis of the different skeletal structures begins at different time points during embryonic development, but all chondrogenic events share the same cellular origin, the MCs. These MCs may arise from three different sources: (1) neural crest cells of the neural ectoderm that eventually form the craniofacial bones; (2) the sclerotome of the paraxial mesoderm, which gives rise to the axial skeleton; and (3) the somatopleure of the lateral plate mesoderm, which yields the skeleton of the long bones. 6 Under the proper signaling, these MCs begin a physical compaction called mesenchymal condensation, which begins with the MC increasing their cellular interaction established both with the ECM and with the surrounding cells (Fig. 1A). Specifically, N-cadherin and neural cell adhesion molecule (N-CAM)–mediated cell–cell interactions are of outermost importance during mesenchymal condensation. 7
Schematic representation of the chondrogenesis and endochondral ossification. (
When the cells aggregate, MCs begin to produce collagen I, fibronectin, and proteoglycans. 8 The result of the strong interactions that cells establish with their environment is the formation of a dense mass of MCs that immediately begins to differentiate into chondroblasts (Fig. 1B). Condensed MCs start expressing mainly the transcription factor Sox9 that controls downstream genes involved in chondrogenesis, promoting these progenitor cells to secrete cartilage-specific ECM molecules (Fig. 1C).9–12
The tissue then undergoes a series of morphological and physical changes during maturation, including an increase in rigidity to acquire its typical cartilage stiffness due mainly to ECM components. Moreover, if cartilage turns into bone, chondrocytes increase in size turning into hypertrophic chondrocytes and enriching the ECM mainly with collagen X (Fig. 1D). 10 Afterward, ECM is progressively degraded, and hypertrophic chondrocytes undergo apoptosis generating spaces used by blood vessels to invade the cartilage (Fig. 1E). Simultaneously, osteoblast progenitors use the blood stream to enter this vascularized tissue. The progressive death of chondrocytes and degradation of cartilage collagens and proteoglycans is synchronized with the synthesis of bone ECM and the proliferation of the osteoblast population within the tissue. In addition to this endochondral ossification that requires a cartilaginous intermediate, there is a second type of bone formation called intramembranous ossification that does not require the transitional step of chondrogenesis. This direct osteogenic process consists of the conversion of MCs into bone tissue (reviewed in Franz-Odendaal et al. 13 ). Intramembranous ossification is the characteristic way in which flat bones of the skull develop. During this process, neural crest–derived MCs (NCMCs) proliferate and condense forming aggregates where some of the cells form capillaries and other differentiate into osteoblasts that secrete a collagen–proteoglycan matrix that is able to promote calcium salt deposition. 14
Finally, it has been noted above that expression of proteoglycans is not exclusive of the synthesis of ECM in mature cartilage formation, but is also important during mesenchymal condensation. For instance, versican, a chondroitin sulfate proteoglycan, has recently been shown to be crucial in mesenchymal condensation to the extent that the absence of the active form of this proteoglycan disrupts subsequent chondrogenesis. 15
Onset of Chondrogenesis: Transcription Factor Sox9 and Genes Related to Its Regulation
Cartilage formation is the first step in the development of almost all skeletal elements in mammal organisms and presents two independent ontogenetic origins. As stated above, craniofacial cartilage proceeds mainly from NCMCs, while other cartilages come from MCs of mesodermal origin.16,17 Although these origins are significantly different from each other, transition of the MC to mature cartilage or bone is very similar in both cases.
As we mentioned above, one of the first steps in chondrogenesis is the mesenchymal condensation first mediated by paracrine factors and subsequently by Sox9, a transcription factor that belongs to the SRY family and contains the high mobility group box (HMG-box) DNA binding domain. 18 The expression of Sox9 is regulated by members of the FGF, transforming growth factor-β (TGF-β), bone morphogenetic protein (BMP), and Wnt families (Fig. 2).8,19 The key role of the transcription factor Sox9 in skeletal development is evidenced by its tight regulation and by the fact that abnormal expression of Sox9 leads to severe skeletal disorders like campomelic dysplasia. Sox9 is responsible for the expression of some of the key genes in chondrogenesis: Sox5, Sox6, and collagen II α1 (Col2a1) (Fig. 2). Moreover, Sox9 regulates expression of collagen XI α2 (Col11a2) and CD-RAP (cartilage-derived retinoic acid-sensitive protein) before matrix deposition in the cartilage anlagen.20–22 L-Sox5 and Sox6 in turn are required for the expression of collagen IX α1 (Col9a1) and aggrecan, which is the most abundant proteoglycan in cartilage (Fig. 2). 23 Moreover, Akiyama et al. have proven that all osteo-chondroprogenitors stem from Sox9-expressing cells, confirming the principal role of Sox9 in chondrogenesis. 24

Upstream and downstream regulation of Sox9. Chondrogenesis begins with the upregulation of N-cadherin and Sox9 by paracrine factors like TGF-β, FGFs, or BMPs, while Wnts downregulate the expression of Sox9. Expression of N-cadherins and Sox9 is accompanied by the condensation and proliferation of MCs, a process called mesenchymal condensation. Sox9 activates the expression of Sox5 and Sox6, and with them and the help of cofactors such as CBP, induces the MC to differentiate into chondroblasts after the condensation. Chondroblasts produce cartilage-specific ECM, including collagen II, IV, IX, and XI; proteoglycans such as aggrecan; and also link proteins and COMP.
In the interdigital region of developing limbs, it is TGF-β that induces the expression of Sox9. This induction is a quick process as the upregulation of Sox9 has been proven to happen only 30 min after exposure of chicken limb bud cells to TGF-β. 19 Apart from regulating the expression of Sox9, TGF-β ultimately initiates the expression of cartilage oligomeric matrix protein (COMP), aggrecan, N-cadherin, N-CAM, Coll II, Coll XI, fibronectin, and tenascin. 25 The strong chondrogenic effect of TGF-β is commonly used to induce chondrogenic commitment of mesenchymal stem cells (MSCs) in vitro, which will be discussed below.26,27
BMPs are another family of the TGF-β superfamily of growth factors that mediate important events within skeletogenesis and body patterning. BMPs induce and maintain the expression of the Hox genes Msx-1 and Msx-2, which are key regulators in antero-posterior limb signaling. 28 Further, BMP-4 and BMP-7 are responsible for specification of neural plate cells to neural crest–derived cells.29,30 In addition, the expression of Sox9 is dependent on BMP signaling via the BMP receptors 1RA and 1RB (Fig. 2). 31 Apart from these early patterning functions, BMPs are also related to the proliferation and maturation of chondrocytes. 32 Gene knockout experiments with mice have demonstrated that BMP-4 promotes the expression of the cartilage-specific (α2) collagen XI, and that BMP receptors promote the synthesis of collagen I, a type of collagen present in the ECM of MSCs throughout most of the chondrogenesis. 33
There is considerable crosstalk between the TGF and BMP signaling pathways, which is evidenced by a synergistic effect of TGF-β and BMP-2 on collagen X α1 (Col10a1) and an additive effect on Col2a1 mRNA expression. 34 Activation of both pathways simultaneously seems to promote chondrogenic maturation. In summary, the members of the TGF-β superfamily (TGF-β and BMPs) promote very early events in chondrogenesis and, directly or indirectly, maintain their regulation during the differentiation and maturation of chondrocytes.
During the development of the craniofacial skeletal structure, maturation of the neural crest cells is also initiated with the expression of Sox9, as happens in all other skeletal tissues laid down through endochondral ossification. However, in craniofacial bone development, expression of Sox9 can be activated by TGF-β–related growth factors and FGFs. The TGF-β superfamily is more commonly responsible for activation of Sox9, but in certain areas like the posterior facial suture, Sox9 is under the control of FGF-2, FGF-3, FGF-8, and its receptors. 35 Further, the FGF family of growth factors is important for the proliferation and survival of cranial neural crest–derived cells. 35 Apart from being involved in the craniofacial skeletogenesis, FGFs are very important patterning growth factors in limb development and regeneration. FGF-8, FGF-4, and FGF-10 have been identified as important factors for the formation and maintenance of the apical epidermal ridge, a structure that directs proximo-distal development of limbs.36,37
Finally, the Wnt family of signaling molecules is also involved in chondrogenesis (Fig. 2). Wnt proteins are distinguished by their capability to activate β-catenin into canonical Wnts (i.e., Wnt3a) and noncanonical Wnts (i.e., Wnt5a). The influence of Wnts on chondrogenesis is twofold and time dependent: at low levels Wnts promote chondroprogenitor differentiation, specifically through modulating Sox9 expression, and later at high levels they promote chondrocyte hypertrophy and subsequent endochondral ossification.38,39 CatnB controls the chondrogenic commitment of chondroprogenitors by negatively regulating Sox9 expression in particular through competing with Sox9 for the CatnB cofactor TCF, (T-cell factor) and upregulation of Wnt4.40,41 Because CatnB is a mutual antagonist of Sox9, 19 noncanonical Wnts, such as Wnt5a, promote early chondrogenesis in vitro while inhibiting terminal differentiation in vivo. 42 Specifically, Wnt5a misexpression delays the maturation of chondrocytes, while Wnt4 misexpression accelerates this process. 41 Further, Wnts also regulate the expression of N-cadherin, an important cell–cell adhesion protein that is directly involved in mesenchymal condensation. 43
Fine regulation of Sox9—more specifically, the balance between Sox9 and the osteogenic transcription factor Runx2/Cbfa1—is what drives osteo-chondroprogenitors to chondroblasts or osteoblasts. For instance, BMP-induced Smad1 and interactions between Smad1 and Runx2 regulate the transition of chondrocytes into hypertrophy44–46 as evidenced in Runx2-deficient mice, in which the late stages of chondrocyte hypertrophy are blocked.47,48 The expression patterns of TGF-β–related proteins, FGFs, and Wnts are responsible for regulating the balance between Sox9 and Runx2 and, therefore, the patterning of the skeletal tissue.
Events Happening After the Expression of Sox9
The expression of Sox9 is the first step toward chondrogenesis, for it induces MCs to become osteo-chondroprogenitors during mesenchymal condensation. After these events, two other Sox transcription factors, Sox5 and Sox6, are upregulated. Sox9 is required for the expression of Sox5 and Sox6, and these three Sox transcription factors (the Sox trio) are then expressed until chondrocytes enter hypertrophy. 18 The Sox trio regulates many important events throughout chondrogenesis, thanks to the different spatiotemporal expression of the transcriptional cofactors (CBP [CREB binding protein], p300, TRAP230, PGC-1α, and TCF) that bind to the Sox proteins (Fig. 2). Other members of the Sox genes, like Sox10 in skull development, can also be involved in chondrogenesis, but they are less frequent. 29
Members of the Sox family of transcription factors directly or indirectly rule the complexity and richness of the cartilaginous ECM. ECM molecules like collagens, COMP, or proteoglycans are only synthesized after the expression of Sox transcription factors, but few of the regulatory mechanisms are understood to date. One of them is the control of the expression of Col2a1, the gene encoding collagen II, which is regulated by Sox9, Sox5, and Sox10 in avian neural crest cells. 49 Other studies also show that Sox9 stimulates the synthesis of collagen II,19,50,51 collagen IX, 52 and collagen XI proteins. 53 Finally, the synthesis of COMP is also regulated by the Sox trio. 54
After the ECM of cartilage is synthesized, eventually chondrocytes enter hypertrophy to render either into a calcified cartilage tissue or into bone, via endochondral ossification as described above. Hypertrophy is enhanced by the thyroid hormone triiodothyronine, 55 but its regulation relies on Indian Hedgehog/parathyroid hormone–related protein (Ihh/PTHrP) signaling. These two proteins create negative feedback gradients that control the fate and proliferation of prehypertrophic and hypertrophic chondrocytes within the cartilage. 56 On the other hand, Sox9 inhibits chondrocytic hypertrophy, explaining why Sox9 and PTHrP regulate each other.57,58
ECM Influence on Mesenchymal Condensation
The tightly regulated transition of MCs to chondrocytes involves not only the time and space–dependent expression of transcription factors, but also cell–cell and cell–matrix interactions. Cells interact with the ECM or with other cells using membrane receptors or different types of adhesion molecules, and these recognitions translate into intracellular signals affecting gene expression, cell proliferation, and migration.
During the mesenchymal condensation and differentiation, cell–matrix and cell–cell contacts have special significance. For instance, many approaches have shown that N-cadherin–mediated cell–cell interactions play a very important role in mesenchymal condensation. 7 Activation of N-cadherin is important for the expression of many chondrogenic markers like Sox9, Sox6, Sox5, aggrecan, and collagen II. 59 Nevertheless, chondrogenesis is also possible in N-cadherin–deficient limbs, thanks to other cadherins, for example, cadherin-11. 60 In any case, cadherins and cell–cell contact are necessary for the condensation of MCs. Therefore, the expression of N-cadherin is regulated by similar mechanisms to those controlling Sox9: TGF-β, BMP-2, and different Wnts influence in the overall N-cadherin expression.43,61
Cell–matrix interactions play an important role in establishing the mechanical strength necessary to pull the MC together in a condensed cell mass. Cells condense and subsequently migrate out from the condensation areas, which requires substantial ECM remodeling. Thus, it is not surprising that the ECM is tightly regulated throughout chondrogenesis to meet the specific mechanical needs in each step of this morphogenetic process. Before the onset of chondrogenesis, MCs are surrounded by an ECM rich in hyaluronan and collagen I.8,62 First of all, during mesenchymal condensation, the ECM is partially degraded to enhance cell mobility and cell–cell interactions. The concentration of hyaluronan in the ECM drops at this initial step. 62 Moreover, matrix metalloproteinases (MMPs) are upregulated to reduce the protein content of the ECM. 63 Although collagen I is detected along all the mesenchymal condensation, 11 MMPs reduce the presence of structural proteins like collagen I. These changes in the protein and polysaccharide content soften the ECM and facilitate a more intimate cell–cell contact that is critical for the mesenchymal condensation to occur. The different MMPs seem to have different roles in chondrogenic development that are dependent on the developmental stage of the chondrocyte. In particular, MMP1 and MMP2 have the capacity to degrade cartilage matrix, and they are characterized as the MMPs that are involved in earlier chondrogenesis. Specifically, blockage of MMP2 function supports precartilage condensation and chondrogenesis, 64 and MMP1 knockout mice show decreased chondrocyte proliferation in the proliferative zone of the growth plates of long bones. 65 In contrast, MMP13 appears to be one of the most important MMPs in late-stage chondrogenesis in terms of cartilage remodeling and mineralization because it is characterized by a substrate specificity for collagen II, the cartilage-specific collagen. 66 Specifically, MMP13 seems to make the space for cell enlargement that goes hand in hand with hypertrophy by degradation of the cartilage matrix.
Finally, mesenchymal condensation is also affected by small proteoglycans such as versican, perlecan, or syndecan. Versican is a chondroitin sulfate proteoglycan that enhances mesenchymal condensation, 15 but the mechanism by which versican is able to trigger cellular aggregation is not fully understood. Versican can bind many structures that are present in the ECM of precartilage MCs such as fibronectin, collagen I, hyaluronan, or tenascin, and it has been shown that versican is necessary for chondrogenic gene expression. 15 Similarly, perlecan is present in very early stages of chondrogenesis (day 12.5 of gestation) during mouse embryo development, and is also capable of inducing cell aggregation, condensation, and chondrogenic differentiation of C3H10T1/2 fibroblasts in vitro. 67 Perlecan is a heparan sulfate proteoglycan that is known to bind other ECM molecules and basement membrane proteins, such as laminin and collagen IV, as well as growth factors, such as FGFs, especially FGF-2 or FGF-9, 68 and BMPs like BMP-2, -4, and -6.69–71 All the interactions established by perlecan explain the importance of this proteoglycan within chondrogenesis, as shown in perlecan-deficient mouse experiments. 72 Especially strong expression of perlecan is observed in the prehypertrophic and hypertrophic zones, 72 but the exact mechanism by which perlecan regulates mesenchymal condensation and chondrogenesis remains elusive.
Finally, syndecans are transmembrane heparan sulfate proteoglycans that have different isoforms and are reported to be involved in cancers, wound healing, angiogenesis, and chondrogenesis. 73 Syndecan-3 is present in tissues undergoing chondrogenesis and shares with perlecan many binding affinities, such as BMP-2. 74
In summary, ECM greatly affects mesenchymal condensation in two different ways: it establishes the adequate mechanical environment and moreover modulates the concentration or activity of important growth factors like BMPs or FGFs. This complex and important role of the ECM during mesenchymal condensation still remains unclear and needs further investigation.
Evolution of the ECM During Cartilage Maturation
Mature cartilage is a tissue with an ECM composed of collagenous and noncollagenous elements. The exact composition of the matrix depends on the type of cartilage, but they all share that collagen II is the main collagenous element (about 90% of the collagenous fraction) within the cartilage. 75 Collagens I, VI, IX, X, and XI are also present in the collagen fibrils with different percentages depending on the particular cartilage. 8 The noncollagenous fraction of the cartilage is basically composed of proteoglycans, hyaluronan, link protein, and interfibrillar proteins like COMP or decorin. Proteoglycans are important components of the noncollagenous cartilage matrix because they are responsible for the mechanical properties of cartilage. Aggrecan accounts for about 90% of the proteoglycan content, and the rest contains decorin, fibromodulin, lumican, biglycan, and perlecan. All the collagenous and noncollagenous components together form an interconnected elastic network surrounding chondrocytes in mature cartilaginous tissues 76 accounting for its unique rigidity and flexibility.
The complexity of the ECM of cartilage arises after the mesenchymal condensation, when cells activate all the chondrogenic machinery and gradually transform the ECM into that of a mature cartilage. Collagen I gradually disappears and is substituted by collagen II, VI, IX, and XI. Collagen II is, by far, the most abundant, and its synthesis is regulated by three members of the Sox family of transcription factors, Sox9, Sox6, and Sox5. 77 More interesting is that collagens I and II can induce chondrogenesis in bone marrow–derived MSCs, which suggests that ECM molecules also regulate gene expression. 78 Collagen X is expressed later on, when chondrocytes acquire the hypertrophic phenotype, and either form calcified cartilage or enter endochondral ossification to become bone tissue.
The short proteoglycans that are present in the mesenchymal condensation (versican, perlecan, or syndecan-3) are gradually substituted by larger and more complex proteoglycans, mainly aggrecan. 15 Link protein, COMP, and all other components of the cartilage ECM are also synthesized to obtain mature cartilage. The regulatory mechanisms that control the synthesis of the noncollagenous elements of the cartilage remain unknown, but are probably dependent on the expression of Sox9.
Finally, if mature cartilage is to become bone or calcified cartilage, chondrocytes acquire the hypertrophic phenotype and express vascular endothelium growth factor. 79 When ossification occurs, the cartilaginous ECM is gradually degraded and invaded by blood vessels, while chondrocytes become apoptotic. Osteoblasts coming from the blood stream invade the tissue, and cartilage gradually transforms into bone. Then, collagen II is replaced by collagen I, the predominant collagen in bone. 80
Sources of Chondroprogenitor Cells
Cartilage has two main ontogenetic origins: the mesoderm and the neural crest. In both cases, MCs are generated from less differentiated Sox9-expressing cells that start skeletogenesis with the mesenchymal condensation. 24 The NCMCs that give rise to the craniofacial skeletal structure are formed from ectodermal cells, which suggests that ectodermal and mesodermal cells are embryonic origins for chondrocytes. Because two out of the three germ layers produce chondroprogenitor cells, it is quite probable that many cells of mesodermal or ectodermal origin could turn into chondrocytes with the proper signaling mechanisms.
In vitro chondrogenesis has been described with adult multipotent cells from bone marrow,26,81,82 muscle,83,84 synovial or periosteal cartilage, 84 and adipose tissue.26,82,84 To date, the results show that the best chondroprogenitors of adult MSC origin are those of the synovial or periosteal cartilage, followed by muscular MSCs and bone marrow stromal cells, which have better chondrogenic commitment than progenitor cells from adipose tissue. 84 MSCs possess a multipotent capacity that is characterized by their ability to differentiate into chondrocytes, osteoblasts, and adipocytes. Further, in their function as stem cells they are able to self-renew, however, only for a limited number of passages in vitro. 85 The best way to isolate MSCs remains elusive. Originally, isolation protocols have been described that rely on the fact that MSCs readily adhere to plastic. 86 Intermittently, the multilineage potential of MSCs has been characterized after isolation by FACS (Fluorescence-activated cell sorting) using a monoclonal antibody against STRO-187,88 and the low-affinity nerve growth factor receptor, also named CD271. 89 Only recently, Pittenger et al. 90 published a landmark paper describing surface marker expression characteristic of MSCs as being CD29+, CD44+, CD71+, CD90+, CD106+, CD120+, and CD124+.
Chondrogenesis from embryonic stem cells (ESCs) has also been reported.91,92 In contrast to MSCs, ESCs have to go through additional developmental stages to reach the fully differentiated chondrocyte state because they represent a pluripotent population. This is achieved through induction with BMP-2 or BMP-4 in murine and human ESCs.91,93 Similar to the in vivo embryonic chondrogenic events, additional induction with TGFs can augment chondrogenesis, 92 underlining the importance of resembling stem cell differentiation systems according to in vivo embryonic development (Fig. 3A, B). Interestingly, these chondrocytic cultures can undergo hypertrophy, mineralize, and express bone-specific markers such as collagen I when challenged with osteogenic inducers like vitamin D3. 92 It could therefore be hypothesized that this particular culture model represents endochondral bone formation (Fig. 3C, F, and Fig. 4). Further, overexpression of human Sox9 in murine ESCs (mESCs) leads to upregulated expression of the cartilage markers collagen IIA, aggrecan, and pax1 even in undifferentiated ESCs. 94 Upon induced differentiation, the adult form of collagen II (the isoform collagen IIB) was expressed by the Sox9 overexpressing ESCs, again emphasizing the importance of this transcriptional regulator.
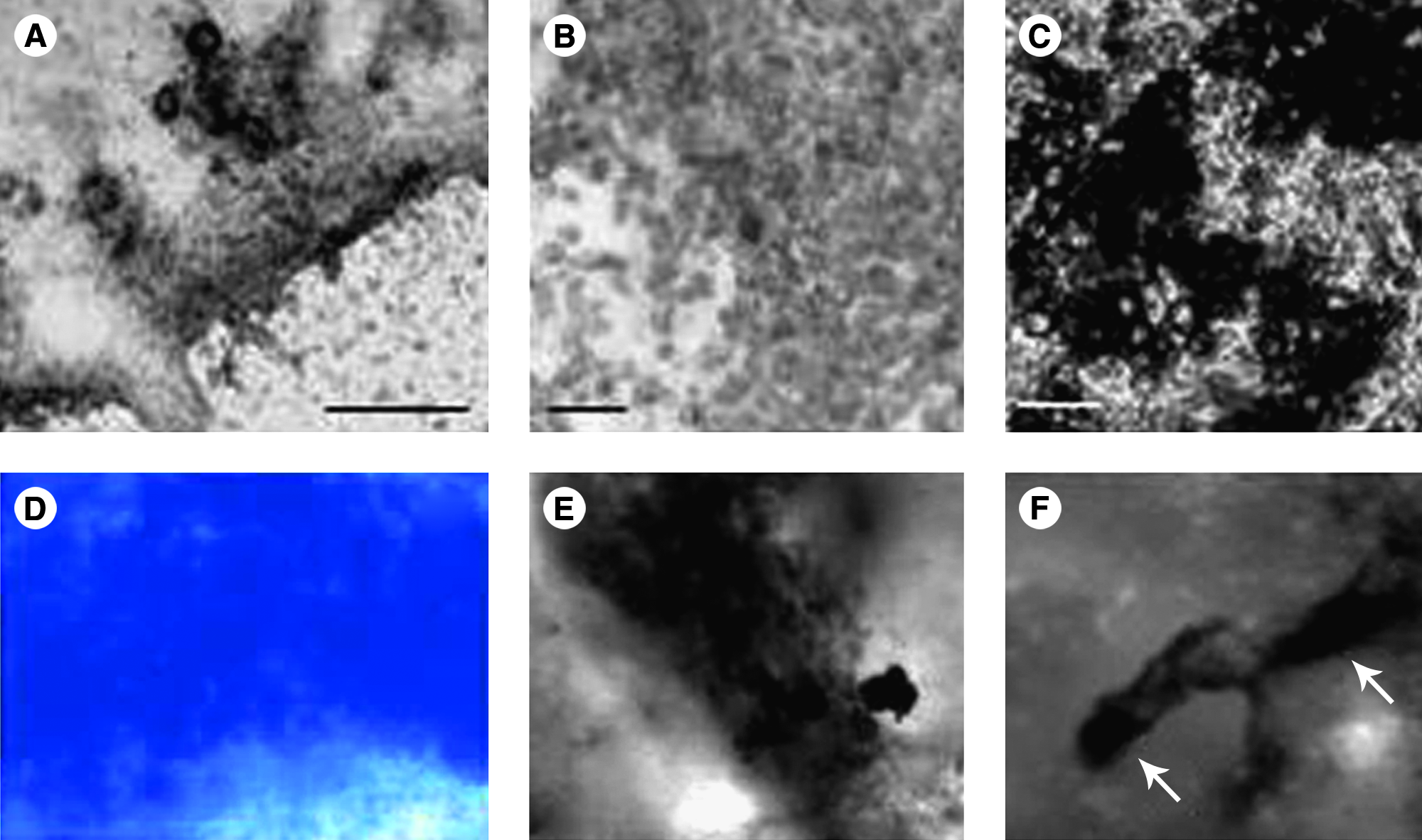
In vitro models for chondrogenesis and osteogenesis with embryonic primitive cells (mESCs and mouse embryonic fibroblasts [MEFs]). (
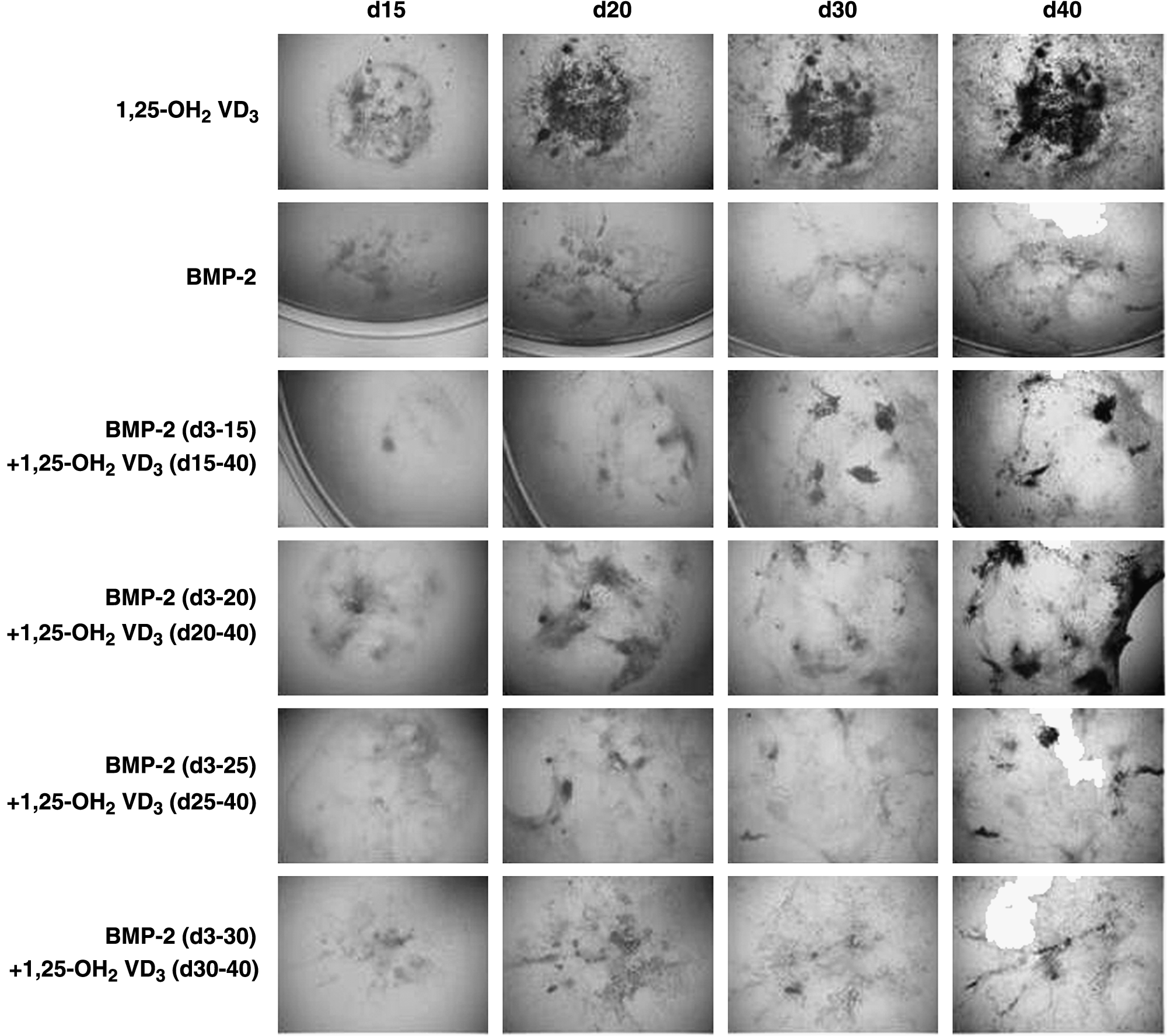
Endochondral bone formation from mESCs in vitro. mESCs were differentiated with 1,25-OH2 vitamin D3 toward osteoblasts, which appear black in phase contrast microscopy. Chondrogenic cultures were treated with BMP-2 from day 3 of differentiation onward. Interestingly, chondrogenic cultures can undergo mineralization when additionally triggered with the osteogenic inducer 1,25-OH2 vitamin D3. However, a window of opportunity seems to exist that falls between day 15 and day 20 of differentiation. 92
Further evidence for how critical it is to modulate stem cell differentiation according to developmental events found in vivo comes from two papers by Hwang et al., who have utilized the supportive effects of ECM molecules to enhance chondrogenic differentiation from ESCs in vitro. In the presence of glucosamine, a component of glycosaminoglycans such as heparin sulfate and hyaluronan, or encapsulated in RGD-modified (arginine-glycine-aspartic acid integrim binding motif ) hydrogels, neocartilage formation from ESCs could be detected accompanied by an increase in cartilage-specific gene expression.95,96
Usage of both ESCs and MSCs in tissue engineering and clinical application is characterized by certain caveats. Although being the stem cells with the highest differentiation potential, ESCs represent a population of cells encircled by moral and ethical dilemma. MSCs, in turn, can be harvested from the patient, however, not without invasive surgery. The search for alternative cell sources is therefore still ongoing. Other possible sources of chondroprogenitor cells appear from the natural process of the limb regeneration in amphibians. There, the first step in limb regeneration is the formation of a cell mass called blastema containing dedifferentiated chondroblasts, myoblasts, and fibroblasts. 97 Although there is evidence that myotubes can enter cellularization in mammals, 98 no chondrogenesis has been achieved with adult myotubes. On the other hand, fibroblasts have extensively shown multipotential capacity99–101 and specifically chondrogenic capacity.67,69 Moreover, fibroblasts can undergo spontaneous chondrogenesis in simple three-dimensional culture conditions (Fig. 3D) showing a high synthesis of proteoglycans. 102 The same chondrogenic cultures can be induced to differentiate into bone-like tissue (Fig. 3E) by inducing the system with osteogenic media. 99 This makes fibroblasts (where embryonic or adult) good and reliable candidates in future therapeutic approaches for cartilage and bone repair.
Mimicking Cell–ECM Interactions with Biomaterials
The broad incidence of the diseases related to cartilage and bone degeneration explains the emergence of a great amount of engineered tissues for cartilage repair. All of them are formed with biomaterials, cells, or a combination of both. The cells to be used vary from mature chondrocytes to any type of the previously described chondroprogenitor cells.
Polylactic acid (PLA), polyglycolic acid (PGA), and polylactic-coglycolic acid (PLGA) are U.S. Food and Drug Administration–approved materials for medical use that are extensively studied for cartilage repair applications.103,104 PLA and PLGA have also been tested in vivo in combination with progenitor cells from the bone marrow 105 and also mixed with articular chondrocytes. 106 The good performance of PLA and PLGA in vivo makes of them one of the reference tools for the medical reconstruction of cartilage and bone defects such as Bankart's reconstruction, which is the reattachment of the capsule and glenoid labrum to the glenoid lip.107,108
Apart from PLA and PGA, other polymers to regenerate cartilage or bone are currently being studied, including polycaprolactones,109,110 polyfumarates, 111 polyethyleneglycol, 112 polyvinyl alcohol,113,114 and polyethylene oxide. 115 All of them are nontoxic, nonimmunogenic, and bioresorbable and have proven to be effective in allowing chondrocyte growth.
Apart from polymers, peptide-based matrices are the other important family of scaffolds used in regenerative medicine. Many natural ECM, such as collagen ECM or the basement membrane, have a high-protein content.116,117 Consequently, many bioinspired engineered tissues have peptides or large proteins within their structure. A whole family of self-assembling peptides has been isolated or engineered over the last 15 years, and many of them have been tested promising for tissue engineering applications.26,118–120 The self-assembly of peptides is driven by noncovalent interactions, such as hydrogen bonds, and polar and hydrophobic interactions giving rise to interweaving nanofibers. 121
An example of a peptide matrix is RAD16-I, a 16–amino acid peptide that self-assembles to form a scaffold with a distribution of pore sizes between 50 and 100 nm. 122 This soft scaffold has proved to be permissive for the growth of different cell types and nontoxic to rats.119,123,124 Recently, RAD16-I was used to recreate mesenchymal condensation in vitro. We have developed an assay with which embryonic fibroblasts, encapsulated in RAD16-I, proliferate and condense to form a dense cell mass with bilateral symmetry. 102 The volume reduction can be as high as 50%, but, more interestingly, bilaterality is accompanied by spontaneous chondrogenesis of the fibroblasts. This phenomenon has many similarities compared to mesenchymal condensation and limb bud regeneration. It is interesting that the condensation of fibroblasts only happens when the peptide that forms the nanofiber matrix is diluted enough to render a soft three-dimensional environment to the cells. When fibroblasts are cultured in slightly higher concentrations of the peptide, which give rise to stiffer hydrogels, they fail to condense although they form cellular networks. This work supports the previously discussed fact that cell–cell contact and a soft ECM are necessary in mesenchymal condensation.
We consider that the use of new paradigms in tissue engineering and regenerative medicine is crucial to obtain in vitro constructs and in vivo therapies with the capacity of truly regenerate damage tissues and organs. Hence, future medical standards for tissue-engineered–derived products will require not only to achieve functionality but also to be safe (nongenotoxic and nontumorigenic), to present high degree of integration with the local tissue (nonrejection and noninflammatory), and to recover natural shape and volume (esthetics). This is way the material reviewed here aims to integrate new concepts and ideas learned by looking at the natural developmental process of chondrogenesis with the hope to create more efficient therapeutic modalities.
Conclusions and Future Directions
Cartilage development is a tightly regulated morphogenetic event where many balances define the fate of chondroprogenitor cells. Much has been studied on gene regulation by different types of signaling molecules (TGF-β, BMPs, FGFs, Wnt, Ihh, and others). Moreover, the onset of the key chondrogenesis transcription factors Sox5, Sox6, and specially Sox9 is well characterized, but less is known of their downstream regulations.
The structure and changes of the ECM during cartilage formation are very well described in the literature. It is also clear that chondrogenesis is affected by the ECM surrounding the MCs. But very little is known of this matter, and major efforts should be undertaken to achieve this goal. The control over cell fate by the ECM could be a very powerful tool for regenerative medicine. We can speculate that for a future therapeutic approach, freshly isolated progenitor cells could be implanted with an ECM analog capable of assisting differentiation and integration of these cells into the host tissue.
Finally, the last 20 years have been very prolific in obtaining mature chondrocytes out of other differentiated or multipotent cells and new biomaterials suitable for tissue regeneration. Briefly, cartilage-like tissues have been obtained from many cell types using different scaffolds. From all chondroprogenitors, fibroblasts have shown spontaneous and high chondrogenic commitment. The fact that fibroblasts can be easily extracted from the skin combined with their chondrogenic potential makes of them good candidates for cartilage repair, and eventually bone.
Footnotes
Acknowledgments
We gratefully acknowledge Alan Grodzinsky for his outstanding help and discussion during the writing of this manuscript. We thank the Catalan Government for the predoctoral fellowship 2005 FI 00864 to L.Q. and to Grant NIH 1-ROI-EB003805-01A1 and Translational Centre for Regenerative Medicine (Leipzig University) award TEMAT 1098SF to C.E.S. that supported this work.
Disclosure Statement
No competing financial interests exist.