
Abstract
Complete axon regeneration after trauma or disease is largely unsuccessful in the central nervous system. With the fast developing advances in tissue engineering and biomaterials, many investigations have identified promising approaches for guiding axonal extension. This review highlights a variety of these approaches and describes the biomaterial properties and signaling mechanisms involved in the fabrication of optimal guidance platforms. The vast majority of axonal regeneration approaches limit themselves to observe how axons elongate and migrate in response to signaling molecules presented on the substrate materials, or more recently, in response to different chemical and mechanical substrate properties. Many of these studies are encouraging in the hope of regenerating axons after disease or injury; however, numerous barriers remain. Here we illustrate the need to optimize a permissive heterogeneous environment for axon elongation using tissue engineering approaches and a thorough understanding of the mechanical properties of the substrate, mechanotaxis, and both attractive and repulsive signaling mechanisms.
Introduction
Regeneration capabilities in the peripheral and central nervous systems
CNS trauma and disease
CNS damage can be caused by injury or disease. Brain trauma and other acute disorders involve injury-induced apoptosis, while chronic disorders, such as Alzheimer's and Parkinson's, involve a slow degeneration of the cellular components of the CNS. 6 In contrast, trauma and injury to the CNS results in tissue damage, and the subsequent formation of a glial scar. Findings involving reactive astrocytes involved in the glial scar suggest that it may have beneficial functions in helping to preserve and protect tissue function and to aid in the repair of the blood–brain barrier after mild or moderate spinal cord injuries,7,8 as well as focal brain injuries. 9 However, the glial scar is notorious for creating a barrier to regeneration (see Fig. 1). A substantial amount of research has been placed on understanding how this response to injury prevents axon regeneration past this site. 10

Schematic representation of spinal cord injury and the formation of a glial scar. (
The glial scar forms within weeks after injury and is largely composed of astrocytes.10–13 The scar formation begins within the first hours after injury with the appearance of macrophages and blood-borne cells, followed by a response in microglia, which begin to divide and upregulate signaling molecules and receptors. 11 Along with microglia, macroglia infiltrate the lesion site, with astrocytes and oligodendrocytes being the major cell types. 13 Although the role of astroglia cells in the CNS has not been fully described, they have been identified to play a role in multiple cell functions, including CNS development, the CNS immune system, the uptake of neurotransmitters, and the preservation of tissue after injury. 13 Astrocytes become reactive when aiding in tissue preservation responses, typically after 1 to 2 days posttrauma, and it is often thought that this reactive gliosis is the major barrier to axonal regeneration after trauma.10,11,13,14 Astrocytes produce multiple types of proteoglycans, including heparan sulfate proteoglycan, which encourages neurite extension, as well as chondroitin and keratan sulfate proteoglycans, which have demonstrated inhibition of neurite outgrowth.10,13,14
Chondroitin sulfate proteoglycans (CSPGs) have been identified in many in vivo injury studies, including localization in the brain after stab wounds, in explants of wounded striatum, in the spinal cord after dorsal root injury and crush injury, as well as in the fornix after transection. 14 These increases in CSPG localization lead many to believe that these molecules may be the most significant components of regeneration–inhibition after CNS trauma. Many studies identify the role of CSPGs in plasticity control,15–18 as well as their barrier function during neuronal development.19,20 The support of many in vivo and in vitro studies that illustrate the inhibitory effect of CSPGs has begun to focus current research designs on abolishing these properties. In particular, it has been identified that digestion of the glycosaminoglycan chains with chondroitinase ABC (a chondroitinase specific for chondroitin sulfates A, B, and C) decreases regeneration inhibition.11,21,22 Recent attention has been placed on these molecules and their methods of inhibiting or encouraging axonal growth. A very simplistic schematic of a CNS lesion and the subsequent glial scar is illustrated in Figure 1.
Because of the described complexity of the CNS, and the intricate formation of the glial scar after injury, it is doubtful that one specific strategy can reverse these traumatic damages. To incorporate numerous strategies into a single treatment, the field of tissue engineering emerges as a highly attractive approach to repair CNS injury and/or disease. The field of tissue engineering can be subdivided into multiple applications, including as a means of controlled drug delivery, as an extension of medical and surgical therapy, as a method to repopulate cellular content of tissues, and as a method to control the normal healing response of tissue. 23 The intent of tissue engineering as a type of wound-healing treatment involves numerous general principles, including the regulation of the normal healing process, the attempt to replace cellular components of disease tissues, and the controlled release of chemical or drug treatments. 23 These principles have allowed the tissue engineering field to emerge as an exciting avenue in the pursuit of CNS repair, in attempt to regulate axon regeneration.
Tissue engineering approaches to axon regeneration
Tissue engineering approaches generally involve the combination of a biomaterial scaffold that supplies optimal environmental cues, with the incorporation of cells and biologically active molecules. A large variety of scaffolds have been evaluated in nerve regeneration approaches and include, but are not limited to, Matrigel,24,25 magnetically oriented type I collagen,26,27 chitosan channels,28,29 and various modifications of agarose,30–32 or fibrin.33,34 Within these studies, modifications of numerous variables have been performed, including changes in porosity, surface area, stiffness, and protein availability. The multifaceted aspects of tissue engineering designs, which incorporate numerous physical and chemical characteristics, emphasize its suitability for axon regeneration models, which proves to require a heterogeneous approach.
Many biomaterial studies have produced encouraging results for axonal regeneration such as the use of bovine aponeurosis to guide neurite outgrowth in vitro, 35 and chitosan channels that promote neural stem and progenitor cell survival during transplantation 28 and increase the number of myelinated axons when implanted in conjunction with peripheral nerve grafts. 29 Unfortunately, these experiments have not resulted in functional recovery, emphasizing the complexity of axon regeneration, and that numerous concepts need to be considered when planning an approach to studying such an intricate process. The majority of tissue engineering approaches for studying axonal regeneration involve the manipulation of either the environment, by preventing inhibitory signaling or encouraging beneficial signaling, or the material by tailoring its properties to encourage maximal outgrowth.
Although axonal regeneration has its challenges, it has been shown that CNS neurons can limitedly regenerate axons after an acute injury.36,37 This process is not spontaneous, but regeneration can be stimulated and guided by appropriate chemical and mechanical environmental cues.38–40 There are numerous potential strategies for designing implants that provide neurons with a permissive environment for regeneration, including the use of autografts made from PNS Schwann cells, the development of cell-free bridges made from biomaterials, and the combination of autografts and biomaterials. However, the disadvantages of autographs make them highly unfavorable for use in CNS injuries. Usually, an acute injury requires immediate treatment, and it takes too long to purify and cultivate the patient's own cells for use in autographs. If instead human donor or animal cells were to be used in the autografts, the transplant could cause harmful immune responses in the patient. 41 Stem cells have recently been used in the incorporation of neural stem cells and Schwann cell–loaded biodegradable scaffolds, 42 in the generation of functional neural artificial tissue, 43 as well as in the promotion of spinal cord tissue restoration. 44 Biocompatibility of various biodegradable synthetic polymers with neural stem cells has been studied 45 ; however, cell-free biomaterials that are biocompatible and allow for control of the chemical and mechanical signals could also be engineered to provide the optimal conditions for neural regeneration. Presently, tissue engineering is a critical field for development of strategies to repair acute CNS injuries. Here, we focus on tissue engineering approaches involved in the development of biomaterials for axon elongation after CNS trauma or disease. We believe that a crucial aspect in developing a tissue construct should rely on a thorough assessment of the biophysical aspects of cell–surface interactions, material requirements, and guidance signaling.
Mechanical Properties of Potential Biomaterials for CNS Repair
Effects of environmental stiffness
One approach in understanding the regeneration of neurons is to examine how the environment of the cells affects their behavior. This understanding is extremely important for the development of synthetic biomaterials that will guide neuronal growth. There has been much recent attention to the effects of environmental stiffness on how cells adhere, spread, and migrate over surfaces.46–55 For example, NIH 3T3 fibroblasts spread more50,52 and develop stronger focal adhesions on stiffer substrates. 50 Endothelial cells also show increased spreading areas and spreading rates, as well as upregulation of α5 integrins on stiffer substrates. 52 The rigidity of the substrate also affects stem cell differentiation. Soft substrates (i.e., brain) favor differentiation into neurogenic cells, medium stiffness substrates (i.e., muscle) favor differentiation into myogenic cells, and very rigid substrates (i.e., collagenous bone) favor differentiation into osteogenic cells. 56 Neural stem cells in particular have displayed the capability to modify differentiation and self-renewal capabilities based on the substrate modulus and biochemical environment. 57 Thus, environmental stiffness can be considered an important cue for spreading, migration, and differentiation of many cell types.
Neuron-like cells have also been shown to be affected by substrate stiffness, but not all neuronal subtypes behave similarly, and not all biomaterials produce the same effects, as shown in Table 1. When grown on polyacrylamide (PA) gels coated with fibronectin, pheochromocytoma cells (PC12s) display longer and more branched neurites, as well as a greater percentage of cells with neurites, on stiffer gels (102–104 Pa). 55 Meanwhile, PC12s on PA gels as soft as 10 Pa possess few, short, and unbranched neurites. 55 In contrast, a separate study evaluating neurite extension on polyethylene glycol (PEG) hydrogels ranging in stiffness between 75 and 400 kPa illustrates that PC12s have the greatest degree of neurite extension on less stiff gels. 54 Note, although in the presence of neurotrophic factors PC12s are capable of differentiating into sympathetic-like neurons,58,59 it would be interesting to identify whether similar behavior occurs using primary neurons.
Y, Young's modulus; G*, complex modulus; G′, shear modulus; 2D, two-dimensional; 3D, three-dimensional; PA, polyacrylamide; PC12, pheochromocytoma cells; PEG, polyethylene glycol; DRG, dorsal root ganglia; PDL, poly-D-lysine; PLGA, poly(lactide-co-glycolide acid); PDMS, polydimethylsiloxane; PLLA, poly(L-lactic acid); PLCL, poly(L-lactide-co-ɛ-caprolactone); RGDS, Arg-Gly-Asp-Ser; YIGSR, Tyr-Ite-Gly-Ser-Arg; IKVAV, Ile-Lys-Val-Ala-Val.
Another study investigated both PA gels and fibrin gels of varying stiffness, and optimal neuronal growth of rat cortices occurs on flexible fibrin gels of stiffness in the range of intact rat brain, which is several hundred pascals. 53 Dorsal root ganglia (DRG) in three-dimensional (3D) collagen gels 49 and 3D agarose matrices 60 possess maximum length of neurites at lower concentrations of collagen (softer gels), while cortical neurons on very soft hydrogels have low viability. 46 It has also been shown that while primary spinal cord neurons thrive on soft PA gels coated with poly-D-lysine (PDL) and Matrigel, glia are unable to survive on the same soft PA gels. 51 Thus, it may be possible to use hydrogels to reduce scar tissue, and substrate stiffness could be used as a method to encourage growth of neurons while stifling growth of inhibitory glial cells.
Effects of substrate topography and dimensionality
Stiffness of the surrounding cellular environment has been suggested to be a key determinant of cell structure. 47 However, other physical features of the cellular environment also influence cellular behavior (Table 1). For neurons, these features include topography,46,61–66 fractional porosity and pore size,51,67 hydrophobicity or hydrophilicity,45,68,69 as well as swelling behavior, interfacial tension, surface-to-volume ratio, and viscoelasticity. 67 Further, cellular behavior depends on whether the neuronal environment is two-dimensional (2D) (such as artificial substrates) or 3D (such as tissue-engineered scaffolds).70,71 It has been well established that 3D matrices are much more effective in stimulating neuron growth than 2D substrates,70,71 most likely because they more closely mimic the native environment of the cells, allowing for innate behavior and function. Recent results, however, have suggested that this preference may vary when neurons are presented with multiple options. 72 Kofron et al. 72 developed an in vitro system designed for the observation of outgrowth behavior in the simultaneous presence of a 2D environment and a 3D environment. For DRG neurons cultured at an interface between a 2D glass slide and collagen gel, the majority of neurite growth was visible at the glass–gel interface. Although there was penetration into the 3D gel, neurites extending from the interface cultures were not as long as those that extended on patterned 2D laminin surfaces. It was suggested that neurite extension may actually be slowed because of the complexity of the interface environment and the multiple options provided to the neurites. 72 In addition to dimensionality, further considerations in the design of biomaterial scaffolds for axon regeneration include biocompatibility, biodegradability, sterilization, and the ability of the scaffold to conform to the dimension of the implantation site. 73
Physical topographical guidance has been shown to be an effective method to enhance axon outgrowth. Many studies on the effects of topography use gels of varying chemical composition, which have been micropatterned to contain grooves of different height, thickness, and depth. Those grooves can act as channels to guide the growth of axons in a preferred direction. Smaller channels with widths of 100 μm in poly(lactide-co-glycolide) are more effective than larger channels in neuronal guidance and result in longer extension of primary neurites, as well as less secondary branching. 64 Murine embryonic cortical neurons, when faced with a step in height on PDL-coated polydimethylsiloxane (PDMS), minimize their turning angle when faced with deeper grooves, while in shallower grooves they are able to extend into or out of the channel. 65
It is also possible for rat DRG neurons to extend neurites over grooves on PDMS, with no underlying solid support. 61 This study also reports that neurons are able to migrate into a groove, extend neurites to either side of the groove and up onto the plateau, and pull the soma of the cell up onto the plateau. The experimental conditions that favor the highest number of these neuron bridges are a cell density of 125,000 cells per sample, groove depth of 50 μm, groove width of 30 μm, and plateau width of 200 μm. 61 These studies indicate that small surface changes can affect axon behavior; therefore, topography can be utilized in biomaterial development in designing pathways for which axons extend in more directed manners.
Combinatorial studies of biomaterials
Several studies also combine topographical features with other mechanical or biochemical features. One such study investigated murine embryonic cortical neurons on PDMS with grooves of varying depth and with coatings of either PDL or Matrigel. 65 PDL versus Matrigel was preferred for adhesion and neurite outgrowth, and a preference for the straightest path possible during axon extension was also shown. When neurons are presented with a conflict of these preferences, axons chose to extend into the Matrigel, and when the only straight pathway is nonpermissive, axons take the path that minimizes the turning angle. 65 Another study of PC12s on PDMS combined effects of microtexture (consisting of microislands in a square array) with mechanical strain. 63 For both smooth and microtextured substrates, lower mechanical conditions (i.e., low strain at high frequency or high strain at low frequency) result in neurons with more and longer neurites, while higher mechanical conditions (i.e., high strain at high frequency) result in fewer and shorter neurites and lower cell density. Meanwhile, the lowest mechanical condition (i.e., low strain at low frequency) shows a significant increase in neurite length on the microtextured gel. 63 Thus, the effects of topography seem to be most influential to neurite outgrowth under low mechanical conditions. Topographical features have also been combined with DNA drug delivery, and controlled release of neutrophic factors into guidance channels has shown to increase the effects of optimal topographical conditions. 64
Varying mechanical and topographical properties have also been combined with biochemical cues, in particular, neurotrophic factors, to better understand axon outgrowth in heterogeneous structures. Evaluating the benefit of neurotrophic factors within a matrix is done either by linking the growth factor directly to the matrix,74–77 or by simply adding the factors into the cell medium. The combination of multiple neurotrophic factors supplied simultaneously has shown interesting results, which should be considered when designing heterogeneous materials. In particular, an investigation on the synergistic effects of combined nerve growth factor (NGF), neurotrophin-3 (NT-3), and brain-derived neurotrophic factor illustrated a lack of directional growth when NT-3 was applied alone; however, when NGF and NT-3 were simultaneously supplied, DRG axons were guided a greater distance (12.5 mm) than that observed for NGF alone (7.5 mm). 32 Additional synergistic effects on DRG outgrowth have been observed with the combination of glial-derived neurotrophic factor, ciliary neurotrophic factor, and NGF. 78 Sharma further describes the neuroprotective role of many NTs and their synergistic effects as potential candidates in spinal cord injury studies using a rat model. 79 In addition to neurotrophic factors, the addition of extracellular matrix (ECM) molecules, cell adhesion molecules (CAMs), and cells native to the injury site is important in attempts to recreate the most optimal environment for axon regrowth. Further details of these signaling cues and their role in axon guidance are discussed below.
Current biomaterials
Tissue engineering approaches to axon regeneration are highly dependent on the types of biomaterials used in scaffold development. Many 2D studies have been done using PA gels, because of their well-characterized mechanical properties and the ability to tailor the gels to a preferred stiffness.80,81 Most frequently, PA gels are used to evaluate cell attachment, growth rates, and morphology changes based on matrix compliance and microenvironment. Neurogel, a 3D polymer hydrogel, has mechanical behavior, viscoelasticity, and porosity similar to the native neural environment and thus has potential applications in neural reconstruction. 67 Another copolymer hydrogel, N-(2-hydroxypropyl) methacrylamide, can be polymerized around dispersed cells and supports growth of axons within pores of the gel; this may potentially act as a scaffold for regenerative growth. 82
PEG hydrogels have also been considered as a potential conduit biomaterial alternative to autografts. 54 Cell adhesion ligands such as Arg-Gly-Asp-Ser (RGDS), Ile-Lys-Val-Ala-Val (IKVAV), and Tyr-Ite-Gly-Ser-Arg (YIGSR) can be covalently linked to PEG hydrogels to increase cell adhesion. In particular, it has been shown that RGDS is a better ligand than IKVAV for extension of PC12 neurites. YIGSR allows for adherence of PC12s but no neurite extension, while Arg-Gly-Glu-Ser (RGES) results in no adherence at all. 54 It would be interesting to repeat these experiments with primary neurons, and to observe whether this observed adhesion and outgrowth behavior is maintained. The stiffness of PEG hydrogels can also be varied in combination with the cell adhesion ligand, creating a matrix scaffold that is easily tailored to enhance neural growth. The wettability (i.e., hydrophobicity or hydrophilicity) can also be varied on polyethylene surfaces through a corona discharge treatment. In these studies, medium wettability (which is mid-range between very hydrophobic and very hydrophilic) results in increased neurite formation. 69
Meanwhile, in a comparison of agarose gels, collagen gels, and fibrin glue, it has been shown that PC12s cannot attach or differentiate on agarose gels, and that the properties of fibrin glue are most favorable for neuron survival. 71 Also, 2D fibrin glue substrates have no effect on neurite length, while PC12s in 3D fibrin glue environments display increased neurite length. 71 In particular, salmon fibrin (as compared to fibrin made from human or bovine blood proteins) promotes the greatest degree of axon and dendrite outgrowth in mouse spinal cord and cortical neurons. 83
Other researchers have studied neuron growth on poly(lactide-co-glycolide acid) (PLGA). PLGA coated with PDL and laminin or PDL and fibronectin results in higher neurite outgrowth than PLGA with PDL and collagen. 68 Additionally, 3D polystyrene matrices that are inert and highly porous allow for differentiation and intricate network formation of neural precursors. 70 Cross-linked hyaluronic acid scaffolds also promote neurite outgrowth in chick DRG neurons. 84
In a recent biocompatibility study, 45 PLGA, poly(L-lactic acid) (PLLA), and poly(L-lactide-co-ɛ-caprolactone) were compared. Of these biomaterials, PLGA was shown to be the best for cell viability, mitochondrial metabolic activity, and apoptotic activity of neurons, and was also favorable for cell differentiation and neurite outgrowth. The hydrophilic property of PLGA allows the actin fibers of cells to be well developed, resulting in increased focal adhesions. PLLA was shown to be the worst in all categories, while poly(L-lactide-co-ɛ-caprolactone) and PLLA are hydrophobic, resulting in poor spreading and underdeveloped stress fibers. 45
Recent attention has been given to carbon nanotubes for many biomedical applications owing to their favorable and unique structural, electrical, and mechanical properties. They can support long-term growth of postnatal cortical and cerebellar neurons, 85 stimulate electrical brain activity, 86 and support neurite outgrowth and branching, 87 especially on those modified with bioactive molecules such as 4-hydroxynonenal. 88 Conducting nanotubular polymers electrospun with PLLA and PLGA are potential mechanisms for precisely controlled drug release, 89 an attractive component in biomaterial fabrication. Further, carbon nanotubes coated with neurotrophic factors such as NGF can be used to differentiate and enhance survival of DRG neurons. 90 However, because of the nonbiodegradability of carbon nanotubes, they would be most beneficial only when long-term drug delivery is necessary.
Many advantageous substrate properties have been discussed above; however, it still remains difficult to establish which biomaterial is considered most effective for applications in CNS injury repair, since variation has been shown among different cell types and environmental conditions. Development of effective synthetic biomaterials for neural regeneration involves, among many other issues, a complete understanding of how the mechanical properties of the material affect cellular behavior and response. In particular, significant attention has been focused on how the stiffness and topography of the biomaterials affect neurite outgrowth. However, it is also essential to consider signaling mechanisms that are occurring at the injury site. Thus, in addition to studying the physical properties of potential biomaterials for neural regeneration, it is also necessary to look at how the biochemical composition of the environment affects axonal outgrowth. These signaling mechanisms will be discussed further in the next section.
Signaling Mechanisms
Although a plentitude of research illustrates the importance of cell adhesion, and substrate and matrix interactions with axonal growth, it has been argued that signaling mechanisms may actually play the primary role. 41 This has been suggested based on research that illustrates poor correlations between axon growth and the adhesiveness of matrix molecules.91,92 Neurites showing little selectivity between N-cadherin, laminin, and L1 in vitro suggests that even though these permissive substrates may define an axon pathway, they are not necessarily instructive (i.e., they do not tell the axon which way to go at an intersection). 91 Although these results bring up the suggestion that substrates may not exert their effects primarily through relative adhesiveness, it is important to note that cell–substrate contact with preferred substrate molecules can dictate which neurite becomes an axon for hippocampal neurons. 93 Recently, it has also been shown that hippocampal neurons are capable of sensing gradual changes in laminin presented on the surface, which may have high physiological relevance, since substrate concentrations in vivo are likely to be gradual. 94 These findings emphasize the importance that neurons recognize their biochemical environment, and can modify their behavior based on the ECM present.
When studying axonal regeneration, there are two types of signaling one must consider: (1) repulsive signaling, which prevents re-growth, and (2) attractive signaling, which encourages neurite and axonal extension. It has been accepted that the reason CNS axons do not regenerate is not because of a lack of growth-promoting molecules, but rather the dominating barrier properties of the glial scar and the inhibitory molecules that overpower the beneficial cues. It is therefore no surprise that signaling molecules and pathways must be understood to engineer materials and create environments necessary for encouraging axonal regeneration.
Cell adhesion molecules
Beneficial signaling includes growth factors,95,96 cell surface and adhesion molecules, and integrin signaling of the growth cone. 97 One of the first factors to consider when engineering materials for axonal regeneration is cell surface signaling, which involves not only mechanotransduction effects but integrin signaling as well. Integrin signaling is critical during axonal development, with CAMs playing a predominant role. Neural cell adhesion molecule (NCAM), L1, and N-cadherin comprise the three CAMs that serve as neuronal receptors encouraging axonal growth during mammalian development. 98 The magnitude of CAMs is evident in cases of L1 gene mutations, which result in neurological deficits, including the absence of corticospinal tracts,98–100 as well as in NCAM deficiencies that result in decreased body weight and spatial learning, and abnormal circadian clocks. 100
Cellular mechanotransduction
The method by which cells sense and respond to their physicochemical environment is not yet well understood. However, mechanical stimuli influence many aspects of cellular function, including growth, migration, and gene expression. 101 It is believed that several cellular components, including integrins, focal adhesions, membrane ion channels, cytoskeletal filaments, and the nucleus, are molecular players in mechanotransduction events. Integrins are the transmembrane links that connect the ECM with the inside of the cell, and they are therefore believed to act in a mechanosensitive way, through inside-out signaling, outside-in signaling, or an interplay between the two. 102 In neurons, the integrins responsible for adhesion to the ECM are NCAM, L1, and N-cadherin. 98 In the extracellular domain, these integrins bind to matrix proteins such as fibronectin, collagen, laminin, and vitronectin. A recent in vitro approach to investigating cellular mechanotransduction has been to observe cells on flexible substrates, which mimic the elasticity of the innate tissue. It is possible that the substrate or ECM acts as a spring whose stiffness influences displacements of integrins on the cell surface; in this model, calcium influx into stress-activated membrane channels may lead to a biochemical cascade inside the cell that results in the displayed cellular behavior.46,47
In the intracellular region, integrins bind to molecules such as focal adhesion kinase, paxillin, and vinculin, forming adhesion structures known as focal adhesions. Focal adhesion assembly increases when external stresses are applied to integrins,103,104 providing evidence for the mechanosensitive nature of these complexes. The focal adhesions link to the F-actin cytoskeletal elements via molecules such as tensin, talin, and α-actinin. Then, the F-actin stress fibers maintain tension between focal adhesions, leading the cell to exert traction forces on its underlying substrate, which can be measured using advanced techniques such as traction force microscopy.103,105–108 However, while some of the molecular mechanosensors have been identified, it is still unclear how mechanical signals might be conveyed to the nucleus of the cell, whether it is through cytoskeletal force transmission, biochemical pathways, or a combination of both. Further, because most of the studies on mechanotransduction and substrate stiffness have been for nonneuronal cells, the method by which neurons in particular sense the stiffness of their surroundings is only recently being understood.109,110 Chan and Odde 109 recently proposed a stochastic model for motility on compliant substrates based on a motor-clutch system, where cells physically link F-actin to the ECM through the use of molecular clutches. Their experimental results using embryonic chick forebrain neurons on PA gels illustrate model-predicted behavior, showing that F-actin/substrate coupling and clutch behavior is dependent on the substrate stiffness. On stiff substrates neurons exhibit frictional slippage, where the F-actin bundle continuously slips from the point of contact; on softer gels, neurons have increased traction forces and prolonged F-actin–clutch interaction times. 109 Cells may use these contrasting substrate–dependent clutch mechanisms as a way to sense the compliance of the surrounding environment, which in turn can signal internal mechanosensing elements, causing potential changes in cell morphology or motility. 109 Many of these mechanosensing elements and pathways are still unclear in neurons; however, receptor-like protein tyrosine phosphatase α (RPTPα), an enzyme involved in the fibronectin-based rigidity response mechanisms of fibroblasts,111,112 has been of recent interest in its relationship to neurite outgrowth and rigidity sensing. 113
Kostic et al. 113 illustrate with RPTPα knock-out mice that although hippocampal neurons differ in response to stiff FN surfaces compared to fibroblasts (neuron outgrowth and differentiation is inhibited, while fibroblasts spread more), the rigidity-detecting mechanisms used appear similar. In their model, they propose that the rigidity-sensing mechanism requires the integrin-dependent activation of RPTPα, which in turn recruits and activates the Src family kinase Fyn at the leading edge. The activation of Fyn then phosphorylates the docking protein, p130Cas, in a ridgidty-dependent manner, in a similar way to that described for the recruitment of these proteins at the leading edge of fibroblasts. 112 These results emphasize that although neurons often have different responses to mechanical stimuli, the signaling pathways that they use for mechanosensing may in fact be similar to those described for other cell types. This includes additional signaling molecules such as paxillin, 114 small GTPases such as Rac 115 and Rho, 116 and focal adhesion kinase, 117 which have all been shown to become activated by mechanical stress in an integrin-dependent manner. Additional understanding on the cytoskeletal changes and mechanosensing elements involved with neuron extension on compliant substrates will greatly benefit tissue engineering designs for axon regeneration, and will likely contribute to more precisely controlled scaffolds.
Guidance cues
Once the importance of neural receptors was identified, specific detail has been placed on how binding to these receptors triggers the internal signaling necessary for axonal extension; however, no concrete understanding has been defined. Recent evidence has supported that NCAM, N-cadherin, and L1 stimulate neurite outgrowth through fibroblast growth factor receptor (FGFR) activation in neurons. In particular, N-cadherin–stimulated neurite outgrowth is inhibited from FGFR blocking agents,118–120 and all the major NCAM isoforms have been illustrated to interact with the FGFR complex. 121
Along with CAMs, detailed studies have been placed on the signaling involved with extracellular guidance cues identified to guide the neuronal growth and axonal extension. Guidance cues have been identified to play a role during embryonic development when neuronal migration is necessary to establish signaling pathways and neuron connectivity. The guidance cues may be either attractive or repulsive for axons and neurons. The current picture of guidance cues and their receptors is not complete. However, 11 molecules and receptors have been at least partially characterized. 122 Most understood guidance cue families include the semaphorins, netrins, slits, and ephrins. Attractive guidance cues come from some netrins and ephrins, yet semaphorins, netrins, slits, and ephrins have all illustrated repulsive mechanisms for steering axonal and neuronal migration.97,122
Semaphorins have illustrated repulsive properties in multiple cell types including facial, olfactory, motor, hippocampal, and dorsal root neurons and more.97,122 The semaphorin family of proteins can be either secreted or membrane associated 123 and were first identified in studies of growth cone collapse activity.122,124 Like semaphorins, slits also act as repulsive guidance cues. These extracellular proteins were initially discovered in Drosophila, by their ability to repel commissural axons from the midline. 125 Slits have the ability to repress the attractive action of netrins when bound to the slit receptor, roundabout (Robo). 97 Netrins are unique in that they can act over both long and short distances, have both repulsive and attractive actions through different receptor mediation, and were the first family of guidance cues to be identified in both vertebrate and invertebrate nervous systems.97,122,126,127 Netrins exhibit their bifunctional qualities during development, when motor neurons are repelled from the midline, yet commissural neurons are attracted to this area.97,122 Ephrins, the fourth class of guidance molecules, also exhibit attractive responses but are primarily seen as repulsive. There are two types of ephrins, A and B, with EphrinA being bound to the ECM, and EphrinB serving as a transmembrane protein.97,122 Ephrins play critical roles during neuronal development by assisting axonal guidance into particular pathways and, more specifically, in retrieving positional information used during retinotectal development.128,129
The ability to understand the signaling molecules, ligands, and receptors active during neuronal development allows for a more complete comprehension of how to encourage axonal regeneration in the most native way. Both the beneficial and inhibitory signaling mechanisms must be considered when engineering approaches and materials for use in CNS regeneration studies. Much interest has been placed on the pathways of axonal guidance and growth cone behavior, and it has thus been suggested that concentrating on these pathways may be the best approach for incorporating growth factors into biomaterials. An example of this is including soluble factors within a matrix, in which the signaling pathway response for neuronal growth to these factors is understood. 41 NGF released from polymeric implants has been shown to stimulate neurite sprouting in PC12s, 130 while the controlled release of NT-3 from fibrin scaffolds increased neuronal fiber sprouting and migration into the legion after ablation injury in rats. 131 (For further understanding on drug delivery methods in neural tissue engineering, see the review by Willerth and Sakiyama-Elbert. 73 )
Growth cone signal transduction
A vast amount of axonal regeneration literature involves the signaling molecules and pathways that activate and encourage the growth cone to migrate in specific directions. As described above, the repulsive and attractive characteristics of the four families of guidance cues have been illustrated for multiple neuron cell types; however, one also needs to understand the steps that follow from guidance molecules to growth cone signal transduction. The key steps in the signal transduction process have been described as (1) binding of an extracellular molecule to receptor on the growth cone membrane, (2) activation of effectors through receptor–cue complexes, (3) activation or inactivation of actin-binding proteins, and (4) modulation of actin dynamics. 97 The growth cone was first described by Cajal in 1890 132 with most recent studies beginning to understand the cytoskeletal dynamics of actin and microtubules during motility (for review, see Ref. 133 ), largely owing to modern high-resolution microscopy. Once the growth cone binds to an extracellular molecule and the effector is activated, these guidance cues can modulate actin dynamics during the migration process. It is necessary to understand the signaling behind this process to manipulate directed axonal extension after trauma. The signaling transmitted intracellularly after receptors on the growth cone interact with the described guidance cues involves the Rho family of GTPases. Rac, Rho, and Cdc42 are three of these adaptor proteins that have illustrated close association with actin cytoskeleton dynamics not only in neurons 122 but in other cell types as well.134,135 Rac is responsible for actin-rich lamellipodia and promotes actin polymerization, along with Cdc42, at the leading edge of the growth cone. Similar to Rac, Cdc42 induces actin-rich filopodia, used for sensing the nearby environment. Unlike Rac and Cdc42, Rho is in charge of mediating repulsive cues, and is believed to contribute to retraction through actinomyosin contraction at the trailing end.97,122 The complicated process of signal transduction pathways downstream of the Rho GTPases that regulates cytoskeletal organization is described in detail by Guan and Rao, 122 yet there is the likely possibility of more activity pathways that remain unknown.
Axon specification signaling
Not only are extracellular and intracellular signaling mechanisms critical in the migration and extension of the growth cone, but they also play a significant role in determining which neurite becomes the axon. Many studies aim to identify proteins that localize within the growing axon, but are absent from the remaining neurites. These proteins are termed “polarity proteins” and are generally suspected to be involved in axon specification, in part, through signaling to the cytoskeletal components such as actin and microtubules. 136 Polarity proteins include growth-associated protein 43, synaptic vesicle antigens, tau, and L1, 137 as well as partitioning-defective (PAR) proteins PAR-3 and PAR-6, which localize at the tips of all neurites before differentiation, yet localize only in the axon of developing growth cones during polarization.136,138 A tentative model for axon polarization and intrinsic signaling is described by Arimura and Kaibuchi 139 in terms of positive and negative regulation. It is suggested that during the transition from neurite to axon there is a disturbance in the balance between positive regulation (membrane recruitment, protein transport, increase F-actin dynamics, and microtubule assembly) and negative regulation (membrane elimination, degradation of proteins, decrease in F-actin dynamics, and microtubule catastrophe). 139 A neurite can elongate quickly and become an axon because of an imbalance of signaling caused by the activation of adhesion molecules and receptors, and the recruitment of signaling molecules. 139 Arimura and Kaibuchi also describe a current list of neuronal polarity-regulating molecules and models of positive and negative signaling. 139
Guiding axonal regeneration with signaling
Overall, signal-passed regeneration approaches involve one of three methods: (1) introducing new attractive molecules or increasing the amount currently present in the environment, (2) preventing inhibitory molecules, or (3) influencing axons to regenerate in a previously inhibitory environment. 11 The attractive and repulsive characteristics of the above-mentioned adhesion molecules, guidance cues, and transduction pathways have lately become incorporated into optimizing many engineering approaches for axonal regeneration. In particular, attempts at delivering both soluble and bound growth and neurotrophic factors have been of important consideration for axonal regeneration in the CNS.131,140,141 Attempts have been made at supplying organized and even patterned quantities of neutrophic factors, and growth and adhesion molecules. Direct supply of guidance cues has been used in collagen implants containing NT-3, which attracts regrowth of neurons and illustrates partial functional recovery in the adult rat spinal cord. 140 With the recent attention of microprinting techniques, patterned arrays of neurotrophic factors have been frequently used on experimental regeneration substrates. Microprinting has been used to direct axons in specific directions by stamping attractive proteins, ligands, and growth factors, in a pathway desired for axonal extension.54,62,68,142 The reverse can also be performed, where microprinting can be used to create repulsive areas, including repellent backgrounds, which discourage cell attachment and movement in specified areas. 143
Other approaches have been taken to modify the intrinsic properties of the growth cone by inactivating Rho signaling pathways, in attempt to ignore the inhibitory proteins encountered in the path of regeneration. 144 Recently, experiments using small interfering RNA and the specific myosin II-ATPase inhibitor, blebbistatin, revealed that repulsive guidance molecule (RGMa) inhibition of neurite outgrowth requires myosin IIA. 145 These findings are helping to shed light on the complex relationships among pathways and contributors that are activated (or inactivated) by neurite outgrowth inhibitors.
Gene delivery approaches have also been employed, and should be greatly considered, since optimal axonal growth requires proper gene expression. 41 Trophic factors have illustrated increases in regenerative activity in multiple types of neurons by upregulating genes involved with growth-associated proteins.146,147 Experiments have also been performed with gene delivery by immobilizing DNA complexes onto substrates in attempt to deliver specific plasmids in separate channels for selectivity of nerve tracts. 64 Plasmid DNA has also been encapsulated for use in gene delivery, showing promising results of neurite outgrowth in a PC12 and primary DRG coculture. 148 Recently, these approaches have been taken one step further, and combined with microfluidic approaches to create spatial patterns of gene expression used to direct cellular extensions. 64
It is apparent that there are multiple ways one can test permissive environments and encourage regrowth by regulating both the inhibitory and attractive cues present to the injured axons. Although there is an abundance of signaling mechanisms known and even experiments performed, there remains no gold standard in regeneration approaches. This makes the ultimate goal of regenerating axons after CNS trauma even more challenging.
Future Directions for CNS Regeneration
Considering many of the described potential research directions and their associated advantages and disadvantages, there appears to be three areas that possess great potential for future consideration in regeneration approaches: (1) manipulating cellular components of the neuron, (2) modifying inhibitory signaling mechanisms, and (3) designing heterogeneous degradable biomaterials that create a more permissive environment.
Manipulating cellular components
One of the major challenges in creating an optimal environment for axonal regeneration lies in the fact that not all neurons express the same receptors for inhibitory molecules, and when subjected to the same environment, different types of nerve cells may deviate in behavior.5,149 This suggests that there is a possibility of different regeneration effects for the same molecules supplied to diverse classes of axons, making it difficult to identify a universal behavior. Some success has, however, been made through the overexpression of growth-associated genes in transgenic animals or through virus-mediated gene transfer. 149 In particular, overexpressing growth cone proteins, including growth-associated protein 43 and cortical cytoskeleton-associated protein of approximate molecular mass, 23 KDa (CAP-23), has been shown to induce spontaneous sprouting in populations of both PNS and CNS neurons in transgenic mice.149–151 The partial degree of axon regeneration reported in these gene overexpression studies is promising for the field; however, it is not fully understood how the axons are capable of overcoming the inhibitory signaling. It has been suggested that gene upregulation may shift the balance between positive and negative cues allowing for outgrowth, or the genes may alter the negative signaling pathways directly, which could reduce the sensitivity to these inhibitors. 149
In addition to genetic modifications and upregulation experiments, manipulation of neuron cytoskeletal components has illustrated encouraging results for potential regeneration applications. Snow et al. 152 recently suggested using cell-permeant peptidomimetics (CPPs) to alter growth cone function. Although preliminary, their work suggests using CPPs to directly manipulate the protein activity in the neuron, for example, through modification of cofilin, one of the cytoskeletal proteins involved in the inhibitory response of the growth cone to CSPGs. Snow et al. were able to illustrate the use of CPPs in identifying cofilin-related peptides as regulators of DRG neurite outgrowth. 152 Observations of murine cortical neuron extension on laminin-coated glass substrates show that the rate of axon extension increases after treatment with blebbistatin, a myosin II inhibitor (unpublished observations). Although others observe a decrease in extension after myosin II inhibition of DRG neurons, 153 the increase in extension that we observe may be specific to cortical neurons, or the concentration of inhibitors or substrate-bound laminin. These types of methods suggest potential applications in modifying a previously stalled axon to grow over a once nonpermissive location, such as the glial scar.
Modifying inhibitory signaling mechanisms
Since inhibitory signaling molecules, such as CSPGs, have illustrated such a powerful role in growth cone inhibition during both development and regeneration, there seems an obvious need to understand these inhibitory mechanisms when evaluating potential therapeutics and biomaterial development for nerve regeneration. Many have begun to describe approaches aimed at blocking CSPG inhibitory mechanisms, including the use of chondroitinase, which has proven successful in increasing axon elongation on scar surfaces.10,11,19,21 Also, attention has recently been placed on the mechanisms by which CSPGs inhibit axonal growth, in particular the glycosaminoglycan chains, which, when enzymatically removed from the core CSPG protein, have been illustrated successful in overcoming CSPG-mediated inhibition in vivo. 19 Laabs et al. 19 describe six potential modes of action by which CSPGs may inhibit axon growth and plasticity, including various CSPG receptor and binding possibilities. A new direction to consider for modifying inhibitory signaling may include the development of two-protein moieties, which allow the binding of an attractive protein to the inhibitory CSPG protein. Blocking this exposed receptor site may disguise the repulsive cues and allow the growth cone to extend forward over the scar region. A method that blocks CSPG inhibition without digesting the core protein may be valuable, since it has been suggested that regions of digested CSPGs left behind may remain slightly inhibitory to neurite outgrowth. 10
Designing heterogeneous degradable biomaterials
It has become apparent that continued research concerning neuron regeneration is going to require combinatorial strategies that utilize the advantageous characteristics of mechanical, chemical, and cellular responses. In terms of biomaterials, this would include a biomaterial that is capable of degrading after the axon has passed the trauma site and reached the synapse target. Degradable materials that are capable of releasing controlled amounts of neurotropic factors would also be beneficial, in particular for encouraging directed growth over a lesion site. Further, developing materials that are heterogeneous in nature may allow for more directed axonal extension. Since different stiffness can dictate varying cell responses, this could be used to create scaffolds with heterogeneous mechanical properties, tailored to suit growth in a particular direction. A heterogeneous nature of various pits and precise linear channels, such as those described by Stokols et al. 30 that have illustrated directed axonal extension, should also be considered when designing an extension pathway. Also, varying the concentration of ECM proteins and types of signaling molecules along the scaffold could potentially direct the neuron using chemoattractive or repulsive signaling mechanisms. Lastly, it is likely that materials will need to be tailored to specific class of axons, considering the wide variety of effects seen when using mechanically similar substrates with different neuron types. A schematic representation of advantageous characteristics and considerations for guidance platform fabrication is illustrated in Figure 2. The inclusion of a variety of these properties and tailoring these properties to specific neuron populations should allow for the most optimal development of axon regeneration biomaterials. It is important to note that even with complete axon extension across the distal portion of the biomaterial, it is still not well understood what type of functional recovery will be restored, and whether the remyelination of axons will need to be promoted with additional interventions. 41 Further focus on functional recovery and myelination will be important following the observation of complete axon regeneration using axon guidance materials.
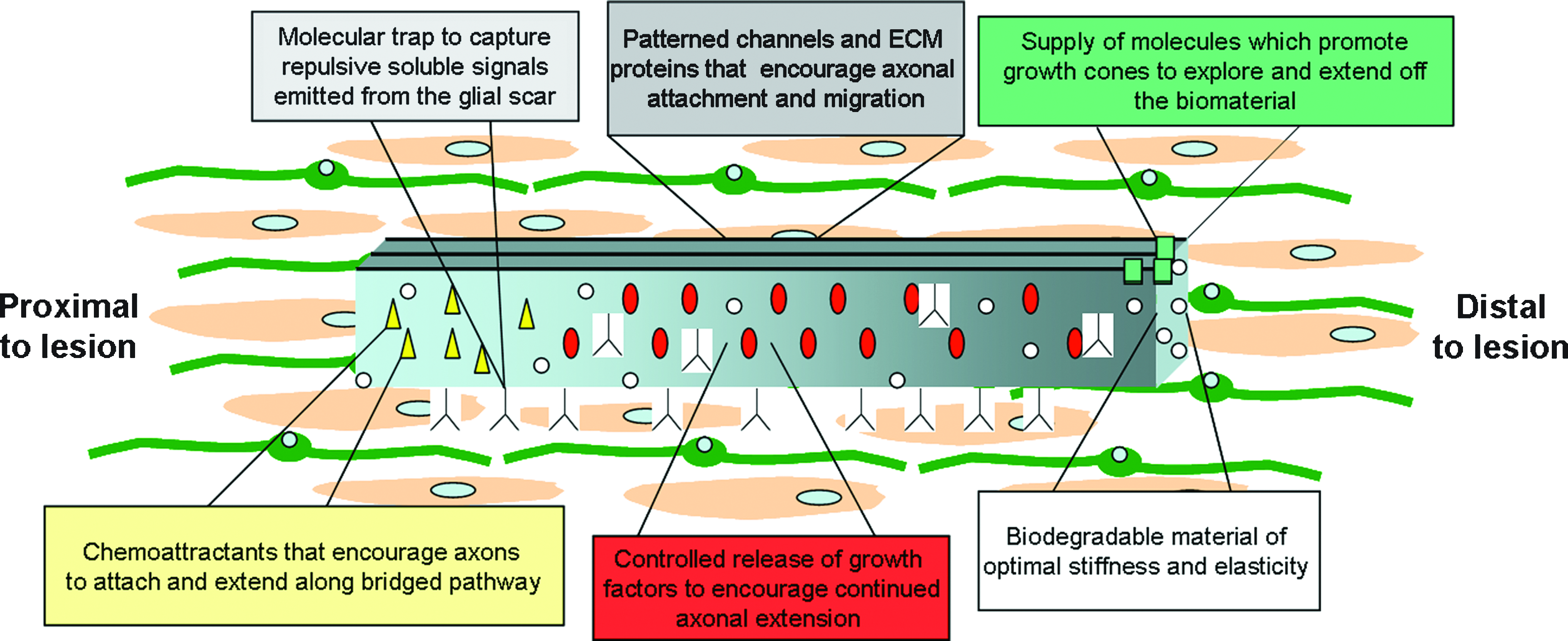
Optimizing biomaterial development for axon regeneration. To promote axon regeneration after spinal cord injury or disease, multiple parameters must be considered in biomaterial fabrication. The most promising biomaterials will incorporate attractive and repulsive signaling and utilize chemical and mechanical substrate properties to encourage directed axon regeneration. The parameters of interest include optimal stiffness, elasticity, topography, porosity, and patterned three-dimensional channels within the scaffold, which encourage directed extension. Importantly, the scaffold must also have a degradability that is appropriate for the time required for complete axon extension past the distal end of the implant. In addition to these mechanical and physical considerations, the most promising biomaterial fabrication will incorporate necessary chemoattractants and repellants, as well as molecular traps that can bind inhibitory signals such as proteoglycans. Molecular traps would be present through the biomaterial, and would help in disguising the once nonpermissive environment of the glial scar. Incorporating a collection of optimal parameters will allow for the development of heterogeneous biomaterials that can be designed for specific types of neurons and multiple injury or disease states. Color images available online at www.liebertonline.com/ten.
Conclusions
The field of tissue engineering has an unmistakable importance in the development of nerve regeneration materials. The mechanical properties, design, and chemical delivery methods will be critical components in guiding axons to their proper target sites. The final step in functional recovery after neurological trauma or disease requires appropriate synapse formations, which can only occur after proper growth cone migration. The most reliable biomaterials will require optimizing advantageous qualities of both mechanical and chemical components.
Footnotes
Acknowledgments
This work was supported by NSF Grant CMMI-0643783 to H.A.E., an NSF Graduate Research Fellowship to K.S., and a Department of Defense CREST Graduate Fellowship to L.L.N.
Disclosure Statement
No competing financial interests exist.