
Abstract
Current treatment modalities for soft tissue defects caused by various pathologies and trauma include autologous grafting and commercially available fillers. However, these treatment methods present a number of challenges and limitations, such as donor-site morbidity and volume loss over time. As such, improved therapeutic modalities need to be developed. Tissue engineering techniques offer novel solutions to these problems through development of bioactive tissue constructs that can regenerate adipose tissue in both structure and function. Recently, a number of studies have been designed to explore various methods to engineer human adipose tissue. This review will focus on these developments in the area of adipose tissue engineering for soft tissue replacement. The physiology of adipose tissue and current surgical therapies used to replace lost tissue volume, specifically in breast tissue, are introduced, and current biomaterials, cell sources, and tissue culture strategies are discussed. We discuss future areas of study in adipose tissue engineering.
Introduction
Adipose Tissue Engineering
To develop an adipose tissue engineering strategy, it is important to understand the complex physiology of the tissue in vivo. This will allow for proper integration of the engineered components so that the regenerated tissue will be structurally, mechanically, and functionally appropriate to the implantation site. This section will focus on adipose tissue physiology, key signaling mechanisms, and current surgical therapies that are clinically implemented.
Adipose tissue physiology and signaling
There are two major types of adipose tissue in humans: brown adipose tissue (BAT) and white adipose tissue (WAT). BAT is most prominent at birth, when it is responsible for the heat generation required to survive the stress associated with the rapid drop in temperature that occurs at birth. 2 Specifically, BAT is capable of nonshivering thermogenesis, in which triglycerides are broken down to generate heat, and this heat is then transferred through the tissue vasculature to warm the body.1,2 At birth, BAT is distributed throughout the body, and can even be found in small depots within WAT. 2 However, as the body matures, BAT content decreases, and WAT content increases. 3 Not only does WAT function as an energy storage depot, but it is also considered an endocrine organ involved with metabolic function and several pathological states. 4 In mature mammalian species, WAT comprises roughly 3% of total body weight in healthy athletes and up to 70% in obese individuals. 5 WAT is distributed throughout the body and can be categorized by type and depot: subcutaneous (including arm, abdominal, and gluteal), intraabdominal (omental, retroperitoneal, and visceral), and other sites (retroorbital, periarticular regions, bone marrow, intramuscular, and pericardial). 6 WAT is predominantly composed of mature adipocytes containing large lipid droplets, in addition to stromal-vascular cells (preadipocytes, interstitial cells, endothelial cells [ECs], and pericytes). WAT contains an extensive system of blood vessels, lymph nodes, and nerves 5 supported within an extracellular matrix (ECM) composed of collagen types I, III, IV, V, and VI and other ECM proteins. 7 Although there is increasing interest in the presence and role of BAT in the metabolism and function of adipose tissue,1,4 this review will focus on tissue engineering strategies that have been developed to address the needs of WAT replacement, as it is the predominant form of adipose tissue in humans.
Adipogenesis is regulated in part by the canonical Wnt signaling cascade (Fig. 1).8–11 Wnt signaling is initiated by the binding of WNT ligands to the frizzled (FZD) transmembrane receptors and low-density lipoprotein-receptor-related protein-5 or -6 (LRP5/6). 12 Upon binding, Disheveled (Dvl) is activated, which then prevents the phosphorylation of β-catenin, resulting in its accumulation in the cytoplasm and translocation to the nucleus. 12 At the nucleus, β-catenin binds to the lymphoid-enhancer-binding factor/T-cell-specific transcription factor family of transcription factors and activates Wnt target genes. 12 In the absence of Wnt signaling, the complex containing glycogen synthase kinase 3, axin, β-catenin, and other proteins exhibits kinase activity that targets β-catenin. In this case, β-catenin is phosphorylated on key serine residues, leading to rapid degradation by the ubiquitin-proteasome system. 12 There are several known antagonists of the WNT pathway, including frizzled-related proteins (sFRPs), Wnt inhibitory factor (WIF-1), Cerberus and Dickkopfs (Dkk). sFRPs, WIF-1, and Cerberus inhibit Wnt signaling by sequestering Wnt factors and acting as inhibitors of the Frizzled receptors. 8 Dkk1, however, blocks signaling by binding with the Wnt coreceptors LRP5/6. 8
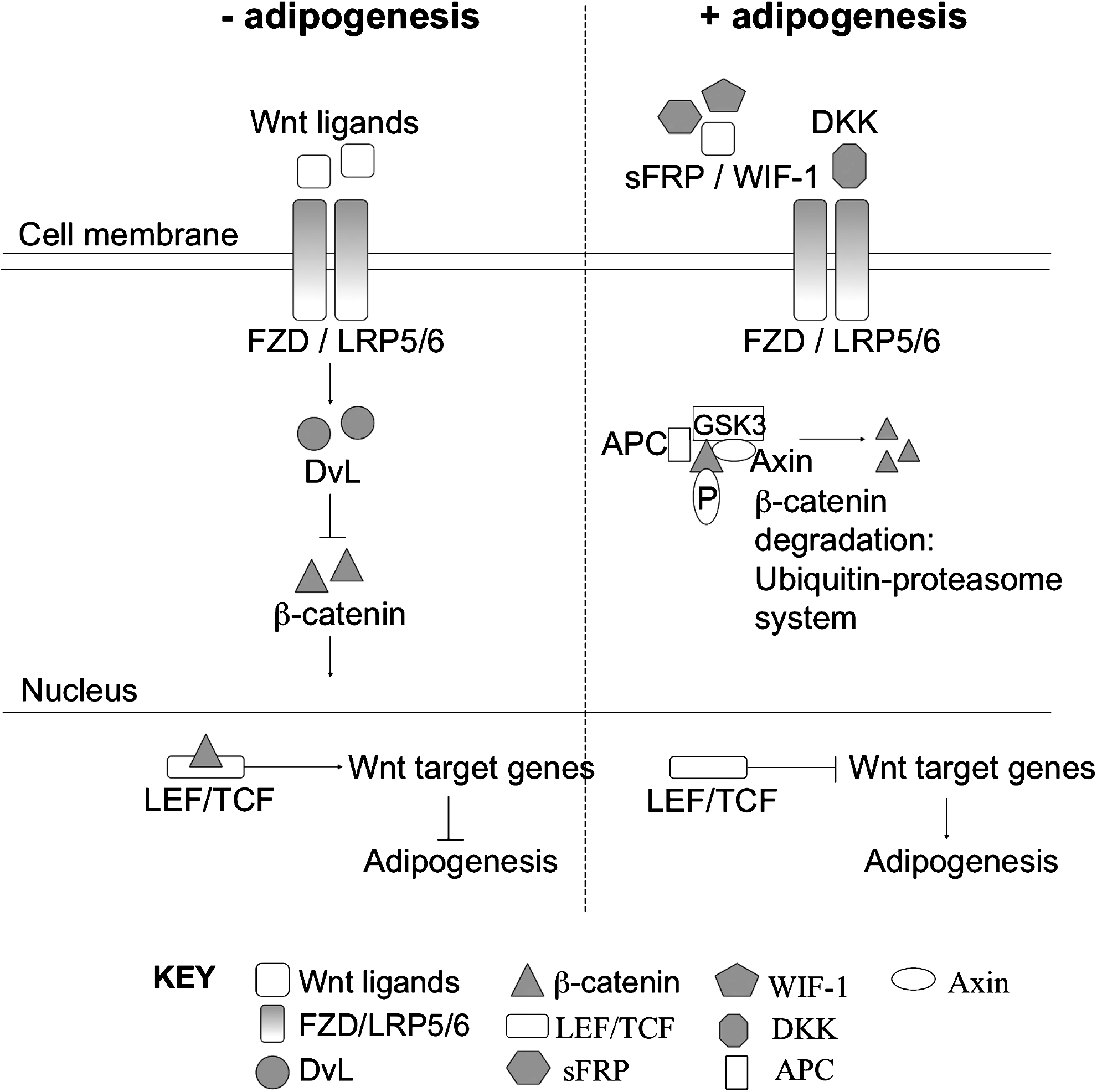
Regulation of adipogenesis by canonical Wnt signaling cascade. In the presence of Wnt signaling, adipogenesis is inhibited through β-catenin-mediated expression of Wnt target genes. Wnt pathway antagonists, including frizzled-related proteins (sFRPs), Wnt inhibitory factor (WIF-1), and Cerberus and Dickkopfs (Dkk), inhibit Wnt signaling and subsequently stimulate adipogenesis.
Recently, Park et al. explored the roles of both Dkk1 and sFRP4 during adipogenesis of human adipose tissue–derived mesenchymal stem cells (hASCs). 8 The mRNA levels of Dkk1, sFRP4, FZD1 and 7, and β-catenin in differentiating hASCs were determined. Upon addition of small interfering RNA that targeted DKK1 and sFRP4, partial inhibition of hASC differentiation and restored levels of β-catenin were observed, indicating that Wnt signaling has a negative effect on adipogenesis. 8 The effects of mutations in LRP5, 13 sFRP1, 14 and Wnt10B 15 have also been investigated to further support evidence of negative Wnt signaling in adipogenesis.
Hedgehog (Hh) signaling has also been shown to regulate adipogenesis in mesenchymal stem cells (MSCs).16,17 Recently, it was demonstrated that activated Hh signaling led to reduced lipid accumulation and decreased expression of adipocyte-specific markers in these cells. 16 Specifically, Fontaine et al. investigated Hh signaling in human MSCs (hMSCs) and showed that a decrease in Hh signaling is necessary during adipogenic differentiation, as Hh was shown to interfere with the function of both C/EBPα and peroxisome-proliferator-activated receptor-gamma (PPARγ). 16
Current surgical therapies
Soft tissue replacement therapies are often required after tumor resection or trauma, or as the result of congenital abnormalities. 1 Currently, there are many reported techniques for the utilization of autologous adipose tissue sources for soft tissue replacement. As adipose tissue substitutes for, specifically, breast augmentation and reconstruction have undergone clinical studies, 18 in this review, we will discuss current surgical therapies that have been implemented for breast reconstruction (Table 1). In the case of breast reconstruction surgeries, these can be categorized into either vascularized flaps or nonvascularized approaches. Common flaps utilized for breast reconstruction are described in depth in a review by Tachi and Yamada. 19 Briefly, vascularized flap therapies typically consist of a large mass of adipose tissue, which is transferred with its blood supply by pedicled flap transposition. 19 Adipose tissue transfer may also include transfer of both muscle and fascia tissue as well.20,21 The superficial inferior epigastric artery flap, transverse rectus abdominis myocutaneous, and deep inferior epigastric artery perforator flaps are among the most common vascularized autologous tissues used for breast reconstruction. Although effective in reconstructing the soft tissue defect, these are major surgical procedures that can carry significant donor-site morbidity.19,22–25 Recipient-site morbidity is an important issue, and is often the result of fat necrosis of the implant, due to the lack of vascular supply, leading to subsequent volume loss with time. 25
ECM, extracellular matrix.
Nonvascularized alternatives are also being utilized. The prosthetic alternative, silicone implants, has long-term complications that include device rupture and capsular contracture. 26 In a search to identify other materials for soft tissue reconstruction besides U.S. Food and Drug Administration–approved silicone implants, experimental studies have explored various ECM and tissue components, mineral/vegetable oils, paraffin, and various synthetic polymers. 1 These materials have limitations such as lack of mechanical integrity compared to native tissue, volume loss over time leading to changes in shape and structure, and the formation of a fibrous capsule, all of which can lead to poor esthetic results.1,27,28 Although current therapies for soft tissue replacement have a reasonable degree of clinical success, additional improvements are warranted.
Key requirements and analysis of engineered adipose tissue
The development of a soft tissue replacement therapy utilizing adipose tissue engineering techniques has generated significant interest in recent years. The development of three-dimensional (3D) in vitro human tissue systems for soft tissue replacement would offer a significant advancement in soft tissue reconstruction strategies. Boudreau and Weaver demonstrated the importance of 3D matrix-mediated remodeling for adipogenic differentiation. 29 The 3D environment not only provides a scaffold structure that functions as a more physiologically relevant microenvironment, but also promotes cell attachment and migration, and facilitates cell–cell and cell–matrix interactions. 30
Based on the weaknesses of current clinical options, a set of key requirements for a tissue-engineered alternative is presented (Table 2). Relevant to any tissue-engineered construct, host compatibility is imperative so that the implanted construct does not elicit host immunorejection. 28 Specifically, site-specific soft tissue reconstruction would optimize host compatibility. Various studies have shown that soft tissue site influences both degree of adipose tissue regeneration and metabolic function of regenerated tissue.31–34 Schipper et al. demonstrated that human adipose-derived stem cells from various subcutaneous depots exhibited significant differences with respect to degree of adipogenic differentiation, sensitivity to apoptosis, and lipolytic function. 34 Additionally, a review by Giorgino et al. describes the metabolic and hormonal differences between fat depots, as well as the different signaling properties of distinct fat depots. 31 Therefore, a thorough understanding of the specific site of interest will be necessary in the design of an engineered adipose construct. Likewise, it is necessary to thoroughly characterize the degradation and mechanical characteristics of the implanted biomaterial, because the material is typically broken down as new tissue is remodeled. 27 In addition, bioactivity of the regenerated adipose tissue needs to mimic that of native adipose tissue, not only in function, but also in structure, which includes the incorporation of a functional vasculature comprising blood vessels, nerves, and lymph supply. 5 Incorporation of the construct upon transplantation is also critical, as the ability of the tissue to completely regenerate over the long term without associated donor-site morbidity or loss of function with time is essential for full restoration of native tissue function.
Addressing these requirements, several assays are commonly used to evaluate the biochemical functions of adipose tissue (Table 3 and Fig. 2). Mature adipocytes, the predominant cell type in the adipose tissue, contain microscopic intracellular lipid droplets, which are the first visible signs of adipose tissue formation. Oil Red O staining and enzymatic assays are conducted to observe and quantify the intracellular lipid, respectively.35–40 In addition, real-time RT-PCR is applied to detect specific transcripts related to adipogenic differentiation and lipogenic activity. Key transcripts that are often measured include the transcription factor PPARγ, glucose transporter-4, fatty acid binding protein-4, and acyl-CoA synthetase.41–43 Glycerol-3-phosphate dehydrogenase, a key enzyme in triacylglycerol biosynthesis, is frequently detected at the levels of both transcript and activity. 44

Schematic of adipose tissue outcome measures. Current adipose tissue engineering strategies examine various aspects of the following adipose tissue outcomes. Adipogenic differentiation is initiated upon inhibition of both Wnt and Hedgehog (Hh) signaling. Peroxisome-proliferator-activated receptor-gamma (PPARγ) and CCAAT/enhancer binding protein alpha (C/EBPα) are key transcriptional regulators that promote expression of adipogenic genes such as glycerol-3-phosphate dehydrogenase (GPDH), glucose transporter-4 (Glut4), fatty acid binding protein-4 (FABP4), and acyl-CoA synthetase (ACS). Triglyceride (TG) synthesis subsequently occurs, causing mature adipokines to be secreted (leptin and adiponectin) and extracellular matrix (ECM) production. TG breakdown, or lipolysis, is mediated by two key lipases (adipose triglyceride lipase [ATGL] and hormone sensing lipase [HSL]), producing glycerol and fatty acids.
PPARγ, peroxisome-proliferator-activated receptor-gamma; Glut4, glucose transporter-4; FABP4, fatty acid binding protein-4; ACS, acyl-CoA synthetase; GPDH, glycerol-3-phosphate dehydrogenase; vWF, von Willebrand factor.
As adipose tissue is recognized as a vital endocrine organ, the engineered tissue's ability to secrete appropriate adipokines, such as leptin and adiponectin, has also been widely investigated.35,45,46 Table 4 summarizes peptide hormones and other endocrine factors synthesized in the adipose tissue.4,47,48 The adipose tissue niche, comprising adipogenic stimulants, growth factors, adipokines, inflammatory cytokines, and thrombosis-associated cytokines, make up the complex microenvironment that stimulate and maintain native adipose tissue.4,37,47–49 There are several known mediators of adipose tissue formation that are commonly utilized. These include the phosphodiesterase inhibitor 3-isobutyl-1-methyl-xanthine; the glucocorticoid dexamethasone; insulin, which is active in glucose uptake and triglyceride formation; and indomethacin, an inhibitor of cyclooxygenase. 30 Basic fibroblast growth factor (bFGF), a key factor present in native adipose tissue, has been widely utilized in adipose tissue engineering strategies, and will be discussed later in this review. Additionally, the microenvironment consists of specific pH and oxygen levels, appropriate ECM, and support cells. 1 This is important as the design and analysis of engineered constructs should reflect closely the native tissue environment. Another key measure of adipose tissue function is its sensitivity to hormone stimulation. Standard assays involve measurements on glucose uptake or glycerol and fatty acid release after stimulation with lipogenic (e.g., insulin) or lipolytic (e.g., catecholamine) hormones.37,46,50,51
In achieving functions reflecting the in vivo architecture of the adipose tissue, demonstrating the ability of the tissue to incorporate blood vasculature is important. Vasculature has been incorporated either in vitro through the addition of vascular cells or in vivo through recruitment of host cells, and is often evaluated through histological analyses for specific endothelial markers such as CD31 and von Willebrand factor.36,38,52,53 Complete regeneration of functional adipose tissue and incorporation of a functional vasculature will only be attained through thorough long-term analyses of both in vitro and in vivo constructs. This includes measurement of the viability and function of the engineered/regenerated tissue, as well as ensuring minimal donor-site morbidity over months and years. In addition, new ECM production is another indicator of mature adipogenesis. Specifically, new ECM deposition, composed of collagen type IV, laminin, and fibronectin, plays a role in the maintenance and remodeling of adipose tissue. 38 Studies have also been reported in which secreted ECM from hASCs has been utilized as a biological scaffold for adipose formation.46,54 Regarding sustainability, there have been very few long-term studies thus far, and this issue will be discussed further in this review.
Biomaterials for Adipose Tissue Engineering
Many biomaterials have been explored for adipose tissue engineering. Both synthetic and natural polymers have been utilized extensively for adipose tissue engineering studies. Considerable advantages and disadvantages with respect to material biocompatibility, mechanical and chemical properties, and degradability for soft tissue applications have been reported. In this review, we will focus on recent applications of common synthetic and natural polymers for soft tissue regeneration.
Synthetic polymers
Synthetic polymers have been widely utilized in adipose tissue engineering. Significant advantages include the ability to specifically tailor a synthetic polymer with respect to its mechanical properties, chemical properties, and degradability (Table 5). 27 For soft tissue applications, considerable work has been performed using polymers such as poly(lactic acid) (PLA), poly(glycolic acid) (PGA), and the copolymer poly(lactic-co-glycolic acid) (PLGA). These polymers degrade by acid hydrolysis into lactic and glycolic acids, and their degradability can be controlled by altering the molecular weight, crystallinity, and the ratio of lactic to glycolic acid subunits. 27
PLA and PGA polymers are predominantly used for 3D meshes, scaffolds, and/or grafts for adipose tissue engineering.36,40,42,43,45,55,56 They have been used for both in vitro and in vivo studies, and have shown potential in supporting regenerated adipose tissue. Long-term studies have been performed in which PGA meshes degrade by 12 weeks in vivo. 40 Briefly, the long-term ability of PGA meshes to support 3T3-L1 cells, a preadipocyte cell line derived from disaggregated Swiss 3T3 mouse embryos, 57 for 24 weeks in nude mice was studied. 40 The long-term effects of varying degrees of adipogenic precultivation on adipogenesis and vascularization in vivo were demonstrated, while the PGA meshes had completely degraded by 12 weeks. Regenerated adipose tissue was evident after 24 weeks in vivo. 40 Likewise, PLA, while showing promise for supporting in vitro adipogenic differentiation, 43 also rapidly degrades in vivo. 42 The ability of PLA scaffolds, along with other common biomaterials such as collagen, to support adipogenesis of hASCs in vivo for 4 weeks was studied. However, the PLA and collagen scaffolds were undetectable by the final time point. 42
The copolymer, PLGA, is more commonly used for tissue engineering studies. Significant work has recently been performed both in vitro and in vivo to explore the ability of this copolymer to support adipogenesis.41,49,53,58–62 PLGA has been utilized as a 3D scaffold to incorporate adipocytes derived from human and rat cell sources as well as additional factors such as bFGF, as this has been shown to induce vascularization in vivo.49,58,60–64 In addition, PLGA microspheres have also been utilized to encapsulate cells, as they proliferate and differentiate in this matrix, as well as to encapsulate factors such as bFGF.41,53,59,65
Other synthetic materials have also been explored for adipose tissue engineering applications, and some show promise for potential soft tissue replacement. These include hydrogels formed from varying compositions of polyethylene glycol (PEG),39,66 3D scaffolds and meshes composed of polyethylene terephthalate and polypropylene,35,36 and various silicone tubing/housing structures for adipose tissue engineering support and delivery.60,67–69
Natural polymers
Natural polymers are materials found as part of the native ECM or those generated by biological systems. 27 The use of naturally occurring materials for tissue engineering applications poses advantages with respect to biocompatibility, and their mechanical and biological properties tend to match those found in vivo. There are numerous polymers derived from native ECM that have been utilized for adipose tissue engineering. Although not comprehensive, Table 6 explores the most common natural polymers used in recent adipose tissue engineering research. This review will focus on collagen, as it is the most prevalent natural polymer used in current adipose tissue engineering research, and silk fibroin, a material that has recently generated interest for this type of application. Additional natural polymers include adipose-derived ECM,38,46,54 decellularized human placenta,7,44,50,70 fibrin,58,63,71–74 gelatin,36,75–81 hyaluronan,44,50,70,82–86 and Matrigel.38,60,67,68,78,80,87,88
3D, three-dimensional.
Collagen is often used for soft tissue engineering owing to its biodegradability, biocompatibility, and weak antigenicity. 81 Collagen has been utilized in many forms, including 3D gels, fibers, scaffolds or sponges, and microbeads. There are a number of studies demonstrating the ability of 3D collagen sponges to support adipogenesis from various cell sources,75,79,89–91 as well as promote development of new adipose tissue in vivo after 12 weeks. 91 Some recent developments include the addition of short collagen fibers within collagen gels to reduce contraction, which increased cell viability and lipid accumulation in vitro. 92 Collagen hydrogels containing FGF-2 encapsulated within gelatin microspheres have been shown to facilitate the development of vascularized adipose tissue in vivo even without the addition of a cell source. 81 These studies show the tremendous potential of collagen in adipose tissue engineering.
We recently conducted comparative studies in which the ability of collagen and PLA matrices to support adipogenesis of bone-marrow- and adipose-derived stem cells was compared to silk fibroin scaffolds both in vitro and in vivo. 42 Although collagen scaffolds supported greater cell attachment upon seeding, and greater lipid accumulation in vitro, both collagen and PLA scaffolds were undetectable after a 4-week cultivation in vivo due to their relatively rapid rates of degradation. 42 Thus, although it is important to establish relevant adipose tissue models containing high lipid content, it is also critical to demonstrate sustainability of the engineered constructs for in vivo soft tissue replacement therapies. Silk has been shown to be a versatile biomaterial for various tissue engineering applications,93–96 with favorable characteristics such as low immunogenicity, slow degradation, absence of bioburdens, and strong mechanical properties.97–102 Additionally, we recently demonstrated the ability to engineer vascularized adipose tissue in vitro by cocultivating adipocytes derived from hASCs with ECs in an aqueous-based silk fibroin scaffold. 103 The ability to tailor silk fibroin as a biomaterial with respect to its biodegradability, mechanical strength, and ability to undergo surface modifications provides strong rationale for the use of silk as a scaffold for soft tissue regeneration. 104
Considerable evidence of adipose tissue formation has been reported in studies utilizing both synthetic and natural polymers. The ability to consistently control material properties of synthetic materials such as PLA, PGA, and PLGA provides considerable advantages over natural materials. On the other hand, natural materials such as collagen and silk have been shown to provide advantages with respect to compatibility, extent of adipose tissue formation, and degradation properties. We have demonstrated in this section that a number of studies have reported adipose tissue formation and maintenance in both synthetic and natural materials. However, additional studies are warranted to further demonstrate and compare specifically long-term in vivo functionality of each material type for clinical applications in soft tissue replacement.
Cell Sources for Adipose Tissue Engineering
Multiple cell sources have been utilized in adipose tissue engineering strategies. These include terminally differentiated adipocytes; preadipocytes of rat, mouse, and human origin, which are cells committed to the adipocyte lineage; adult stem cell sources; and embryonic stem cell (ESC) sources. Although varying degrees of success have been reported using these cells, this review will restrict its focus to human stem cell sources for clinical adipose tissue engineering. A review of stem cells utilized in adipose tissue engineering has been previously reported, 30 and the present review will focus on recent advances in the use of both human adult, or multipotent, and pluripotent cells for soft tissue replacement therapies. A summary of literature references regarding recent progress in human stem cell sources for adipose tissue engineering is presented in Table 7.
Human mesenchymal stem cells
hMSCs can be derived from adult bone marrow (hBMSCs) or adipose tissue (hASCs). Both hBMSCs and hASCs are multipotent cells. Many studies have demonstrated that these cells can differentiate into multiple tissue lineages. hBMSCs have been shown to differentiate into bone,105–107 cartilage,108,109 adipose,42,106 myogenic,110,111 neuronal,112,113 and tendon 114 lineages, whereas hASCs have been shown to differentiate to bone,115,116 cartilage,117–119 adipose,39,103 myogenic, 120 and neuronal 121 lineages.
The use of hBMSCs for adipose tissue engineering has been prevalent in recent years. As described earlier, hBMSCs are stimulated toward adipogenic differentiation with the addition of 3-isobutyl-1-methyl-xanthine, dexamethasone, insulin, and indomethacin to the standard growth medium Dulbecco's modified Eagle's medium (DMEM) + 10% fetal bovine serum. 122 Many studies have shown the utilization of adipogenic-differentiated hBMSCs for engineered adipose tissue both in vitro and in vivo on a variety of biomaterials. Recently, the ability to culture adipogenic differentiated hMSCs on 3D collagen scaffolds in vitro for 50 days was demonstrated. Lipid accumulated adipocytes were found within a bed of remodeled collagen after the culture period. 90 Other recent studies include the culture of adipogenic-differentiated hBMSCs on an electrospun poly(L-lactic acid) (PLLA) scaffold for over 21 days, 43 as well as within a Pluronic hydrogel. 39 In both studies, the biomaterial formed an appropriate microenvironment for adipogenesis from hBMSCs and lipid-accumulated adipocytes were evident. Although progress continues with respect to the use of hBMSCs for adipose tissue engineering under a variety of culture conditions and biomaterials, one disadvantage of these cells is that the relatively painful procedure of obtaining human bone marrow limits their clinical application.
The more recent discovery of hASCs has led to numerous studies addressing soft tissue reconstruction for a variety of reasons. hASCs have been demonstrated to be widely available, abundant, and easily accessible through procedures such as liposuction and biopsies. 123 Approximately 400,000 liposuction surgeries are performed each year, and a range of 100 mL to 3 L of aspirate tissue is collected, in which multipotent stem cells can be isolated. 124 This tissue is normally discarded, but new studies indicate that the use of this tissue as a stem cell source has provided enormous potential for tissue engineering applications. 125 A comparison of hBMSCs to hASCs showed that both sources can differentiate toward multiple lineages, and both also exhibit similar cytokine secretory profiles. 126 Likewise, hBMSCs and hASCs share surface immunophenotypic markers in common. Both populations are positive for tetraspanin (CD9), the aminopeptidases (CD10 and CD13), β1 integrin (CD29), hyaluronate receptor (CD44), 5′ ecto-nucleotidase (CD73), Thy-1 (CD90), endoglin (CD105), activated leukocyte cell adhesion molecule (CD166), and histocompatibility locus antigens ABC (Class I), while negative for the hematopoietic markers mac-1 (CD11b), the endotoxin receptor (CD14), common acute lymphocytic leukemia antigen (CALLA, CD45), and the histocompatibility locus antigen DR (Class II).127,128 Further, a comprehensive phenotypic characterization of hASCs as a function of passage contributed to our knowledge of hASCs and their behavior in culture. 129 The similarities that exist between hBMSCs and hASCs show promise for hASCs, as hBMSCs have developed as an important cell source for human adipose tissue engineering. Addressing the limitations in hBMSC isolation, these similarities present hASCs as a strong alternative. It is not surprising, therefore, that hASCs have been the predominant cell source for adipose tissue engineering studies in the past few years. Remarkable progress has been made with respect to the culture of hASCs on both natural and synthetic scaffolds, in vitro and in vivo, and for various culture time frames. In addition to exploring hASC viability and differentiation capacity on various materials,18,38,50,59,77,130–133 contributions of hASCs toward vascularization36,70,85,134 and wound healing 135 have been investigated.
Human pluripotent cells
Cells that have the capacity to differentiate into cell types from any of the three embryonic germ (EG) layers are defined as pluripotent cells. Two of the pluripotent cell types used today are ESCs, which are derived from the inner cell mass of the blastocyst, and EG cells, which are derived from primordial germ cells. 136 The advantage of these pluripotent cells over other sources such as MSCs is that these cells can differentiate into cells from all three layers and they can also be maintained in culture for a longer time without losing their differentiation potential. 137 The use of pluripotent cells for adipose tissue engineering has been limited to date. However, studies designed to understand the processes involved in adipogenic differentiation of these cells are underway.
In one study, adipogenesis was induced in murine ESCs derived from the inner cell mass of murine blastocysts. 138 Both lipogenic and lipolytic function were demonstrated, and expression of adipocyte-related genes was shown. 138 After this, the discovery of EG cells was reported and the pluripotency of these cells was demonstrated. 136 A number of studies have been conducted to investigate the differentiation potential of embryonic cells.139–143 In particular, the derivation of multipotent MSCs from human ESCs (hESCs) has been demonstrated, and these cells were able to differentiate into adipocytes. 137 Also, mutant hESCs have been developed to further explore the mechanisms involved in adipogenic differentiation. For example, hESCs have been transduced with Oct-4 shRNA; Oct-4 is a transcription factor that is involved in maintaining the undifferentiated, pluripotent state of hESCs. 144 Transduction of hESCs with Oct-4 shRNA led to an extremely high rate of spontaneous differentiation toward an adipogenic phenotype. 144
Investigations using pluripotent cells to generate 3D adipose tissue constructs are still limited. However, in recent years, adipogenic differentiation of EG cells in 3D PEG diacrylate hydrogels was demonstrated both in vitro and in vivo, and the outcomes were compared to those achieved using hMSCs. 145 Through Oil Red O staining and RT-PCR, it was demonstrated that EG cells accumulated lipid when cultured in 3D, and expression of adipogenic genes such as lipoprotein lipase and PPARγ was comparable to levels exhibited by hMSCs under similar conditions. 145 The continued development of adipose tissue engineering strategies involving pluripotent cells is a promising strategy for clinical application in soft tissue reconstruction.
As reported in this review, an abundance of studies have been reported with regard to the utilization of human adult MSCs, both bone marrow and adipose derived, for adipose tissue engineering. The ability to differentiate these cells toward adipocytes has been demonstrated, and the formation of tissue both in vitro and in vivo is widely accepted. However, the more recent use of human pluripotent cells for adipose tissue engineering, though early in development, shows tremendous promise for native adipose tissue formation. With the continued discovery and progression of adipose tissue engineering research, soft tissue replacement therapies utilizing human cell sources is increasingly becoming more feasible.
Culture Strategies for Adipose Tissue Engineering
There are numerous adipose tissue engineering culture strategies in which the core tissue engineering principles comprising appropriate cells, scaffold, and microenvironment are optimized. 1 This section will focus on various culture strategies to alter the microenvironment for adipose tissue outcomes. Specifically, we discuss static versus dynamic culture, cocultivation, addition of growth factors, vascularization, and long-term sustainability of engineered constructs.
Typically, cells are differentiated in vitro, assayed to ensure that they possess adipose-like characteristics (Table 2), and implanted to test the in vivo response. However, in vitro precultivation strategies can be complex and vary tremendously from one study to another. For example, most reported adipose tissue engineering studies have been conducted under static culture conditions. However, there have been several studies reported in which dynamic culture conditions were utilized (Table 8). There are significant advantages to dynamic cultivation of adipose constructs, including increased nutrient and oxygen delivery to cells within a 3D construct. 146 Improved bioreactor designs to address direct perfusion conditions are necessary to advance dynamic culture techniques.
The static coculture of differentiated adipocytes with an additional cell type is an important strategy that has been explored to a limited extent.103,147 Recently, we conducted a study in which hASC-differentiated adipocytes were cocultured with ECs for 2 weeks in vitro to mimic in vivo conditions more closely. We found that ECs formed lumen-like structures in coculture and we obtained adipose-tissue-like constructs using this technique. 103 3T3-L1 cells were cocultured with ECs, and EC proliferation and 3T3-L1 differentiation were enhanced. 147 In addition to cocultivation, another strategy that shows promise for adipose tissue engineering is dynamic cultivation.45,146,148 Precultivated adipose constructs are prepared using spinner flasks45,148 or rotating bioreactors 146 to enhance transport of oxygen and nutrients. In these cases, dynamic culture of adipose constructs promoted enhanced adipose tissue formation. Specifically, lipid accumulation, expression of key adipogenic genes, and adipokine secretion were enhanced.45,146,148 Finally, the incorporation of a second biomaterial into a 3D scaffold is also a common strategy employed to either increase the mechanical integrity of the scaffold,53,58,70,79,81,92 or serve as a delivery vehicle for exogenous factors such as bFGF.37,49,68,75,80,88 There have been numerous studies in which bFGF has been adsorbed or delivered to adipose constructs both in vitro and in vivo, and in most cases, bFGF contributed to greater degrees of adipogenesis and vascularization.37,49,68,75,80,88
The problem of adequate vascularization is common to many tissue-engineering applications. Sufficient, functional vasculature is vital for the construction of larger constructs, in which nutrient and oxygen diffusion is limited by distance and metabolism, during in vitro cultivation and especially after in vivo transplantation, when the incorporation or integration of vessels needs to occur immediately. 28 There have been limited studies regarding the vascularization of adipose tissue. As mentioned previously, our recent coculture experiments allowed us to generate endothelial lumens within a bed of 3D adipose tissue in vitro. We cocultured hASC-derived adipocytes with human umbilical vein ECs for 2 weeks on silk fibroin scaffolds, and observed lumen formation within the constructs using CD31 immunostaining as well as observation of adipose tissue properties, including lipid accumulation and leptin secretion. 103 Stosich et al. also investigated vascularized adipose tissue formation. In their study, PEG hydrogels with and without microchannels were constructed, and these were implanted in severe combined immunodeficient mice with or without adipogenic differentiated hMSCs and/or bFGF. 37 After 12 weeks, lipid-accumulating adipocytes were evident, along with signs of newly formed vasculature from vascular endothelial growth factor and lectin immunostaining. 37
Finally, the long-term viability and sustainability of adipose tissue constructs must be determined. The ability of an implanted tissue-engineered construct to remain viable and active in vivo until complete tissue regeneration is obtained is of critical importance for clinical applications. As mentioned previously, silk fibroin is a biomaterial that possesses in vivo degradation properties that can be tailored to allow the material to degrade after as few as 2 months to as long as 1–2 years in vivo. 149 It has also been shown that silk fibroin scaffolds can support in vivo adipogenesis for a minimum of 4 weeks. 122 Weiser et al. performed a 24-week-long in vivo study in which they demonstrated the capacity of PGA fiber meshes, which were seeded with differentiated 3T3-L1 adipocytes (predifferentiated up to 35 days in vitro), to remain intact, and they obtained adipose tissue over the entire culture period. 40 Although this is a promising outcome, additional studies are necessary to demonstrate the development of sustainable human adipose tissue with complete viability, function, and regenerative potential.
Numerous culture strategies have been implemented, and have shown various degrees of adipose tissue formation. As mentioned, strategies involving cocultivation, dynamic culture conditions, vasculature incorporation, and sustainability all play a role in altering the microenvironment of cultured cells to impact tissue formation and function. Although a rationale exists for each culture strategy tested, clinical translation of adipose tissue engineering strategies for soft tissue replacement, however, will require thorough understanding and analysis of the complex tissue environment.
Concluding Remarks: Future Directions for Adipose Tissue Engineering
As the demand for soft tissue replacement techniques continues to increase, adipose tissue engineering strategies will continue to progress. A variety of biomaterials, cell sources, and culture strategies have been explored, and while it is both interesting and exciting to investigate the potential of these numerous options, it is important to find the appropriate avenue to advance for clinical applications. Clinical goals for adipose tissue engineering include regenerated tissue that both cosmetically and mechanically resembles that of native tissue. This includes mechanical integrity as well as sustainability and viability of the tissue over time. Additionally, adipose tissue that possesses active metabolic function would serve as a significant advancement over current soft tissue replacement strategies.
Currently, clinical trials have been conducted for soft tissue cosmesis applications in which a cell-assisted lipotransfer technique was developed for cosmetic breast augmentation.18,150 To our knowledge, however, human clinical studies utilizing 3D engineered adipose tissue constructs for soft tissue reconstruction have not yet been initiated. As current clinical defect tissue requirements are typically on the order of cubic centimeters, 19 engineered adipose constructs that are of significant dimension are necessary. Addressing this, the authors recommend that significant emphasis on vascularizing 3D adipose tissue and dynamic precultivation would allow for larger constructs, and enhance functional adipose tissue formation and maintenance. The ability to deliver appropriate nutrients and oxygen to an adipose construct no longer limited by diffusion, as well as improvements in long-term sustainability of the engineered tissue made through appropriate biomaterial selection, would significantly enhance the potential of engineered adipose constructs. Future work addressing each of these important strategies is required to move closer to a clinically relevant engineered adipose tissue for soft tissue replacement therapy.
Footnotes
Acknowledgments
The authors thank the NIH P41 Tissue Engineering Resource Center and Armed Forces Institute of Regenerative Medicine (AFIRM) for support of aspects of research on soft tissue engineering. The authors also wish to thank Dr. Jennifer Olson for editorial assistance.
Disclosure Statement
No competing financial interests exist.