
Abstract
Cell adhesion to scaffolds has remained one of the challenges in tissue engineering. Although protein surface modification has been proven to enhance cell adhesion and retention, its specificity depending on cell and biomaterial types means that the best protein and concentration must be established for each specific application. This review focuses on the improvement of cell adhesion for human mesenchymal stem cells with an osteogenesis approach. A brief outline of the cell adhesion process and extracellular matrix proteins precedes an overview of works focused on the adhesion of mesenchymal stem cells and osteoblasts to biomaterials and this effect in their differentiation into osteoblasts.
Proteins Involved in Cell Adhesion
Cellular adhesion to these molecules is mediated by integrins, surface receptors that recognize peptide sequences. This process regulates not only cell attachment to surfaces but also cell-to-cell and intracellular signals. The integrin structure includes two distinct subunits: an α chain and a β chain. Multiple integrin subunits have been characterized, and although this means countless combinations, only around 24 functional integrins are found in mammals. Integrins target specific amino acid sequences; however, as these are constituents of different extracellular matrix (ECM) proteins, one integrin will bind to all the proteins containing the target sequence.3–7
To mimic the biological processes during wound healing and tissue remodeling, tissue engineering considers the use of ECM components as a key factor in tissue repair. ECM provides not only the structural support for cells but also the biochemical cues to moderate cell physiology and phenotype (Fig. 1). Integrins recognize and translate these signals into the indicated response. Some receptors mediate adhesion to other cells (leading to assembly of three-dimensional [3D] structures), and others mediate adhesion to a physical surface to establish cellular orientation and spatial organization. 8 Cell adhesion onto a biomaterial can be either specific, with cell receptor recognition and binding to proteins or peptides, or nonspecific, by noncovalent attractive forces between cells and the biomaterial. Both improve cell adhesion and proliferation, but in the case of nonspecific adsorption, higher concentrations of coating are needed to achieve the same effect that is achieved with specific adhesion.
Main roles of the ECM in cell biology. 74 ECM, extracellular matrix.
Brighton and Albelda proved that ECM proteins promote bone cell adhesion; thus, orthopaedic biomaterials should either include or promote ECM production with the aim to mimic cellular environment. 9 By integrating a material with ECM components, cells are able to maintain a stronger interaction with the biomaterial and with adjacent cells.
The first strategy is to biochemically modify material surfaces by adsorption of ECM proteins. A chemical approach involves ion-exchange reactions, dissolutions, and degradation to immobilize and attach compounds to surfaces. Covalent coupling is also possible either by enzymatic attachment or during matrix formation. Bioactive surfaces can incorporate ECM proteins to elicit particular cell responses. Peptides are also used, as these are recognizable sequences of the ECM proteins. The peptide arginine–glycine–aspartic acid (RGD; mainly present in fibronectin, but also in collagen, laminin, pronectin, and vitronectin) is the major recognition sequence for integrins. 3 These integrins, with different subunits αβ, bind to ECM proteins recognizing RGD-dependent and RGD-independent ligands, overlapping in functions between integrins and giving a broad ligand specificity. 10 As a result, RGD has been widely used to create biomimetic environments to promote cell adhesion, retention, and differentiation. These biofunctional material approaches range from simple adsorption and patterning of the molecule in surfaces, to synthesizing hybrid materials, including metals, polymers, and hydrogels.11–17 There have been other sequences used to promote cell adhesion such as the recombinant human fibronectin peptide (hFNIII9-10) that induce the same cell adhesion levels as fibronectin but a higher osteogenic effect.18,19
In addition to ECM proteins and peptides, complex compounds such as Matrigel and serum are also used in vitro to modify cell adhesion efficacy. Although most of them are well characterized, these compounds have the disadvantage of being unsuitable because of its animal origin or because of the unpredictability of unknown components. 20
After fibronectin, collagen is the most used compound to modify surfaces as it represents nearly one-third of all proteins in the human body and bone is not the exemption. Bone structure is made up of 70% of inorganic material and hydroxyapatite, while the rest is an organic matrix. From this, collagen constitutes 95% and is responsible for its flexibility and strength.21,22 Collagen type I supports the mineral scaffold and it has been confirmed that it binds to integrins α1β1 and α2β1. 23 Other noncollagenous proteins also used for bone cell adhesion and/or differentiation improvement are shown in Figure 2.

Laminin, vitronectin, fibrin glues, hyaluronic acid, and alginates are also used for clinical applications, including the coating of implants. Table 1 shows the functions of the three main ECM proteins used in surface modification. Coatings are also used to direct cell growth into specific patterns. Precise configurations are achieved by polymer lithography, photo-lithography, deposition, or printing of concentration gradients. These differences in surface structure prompt distinctive responses at different times. These techniques also allow for specific cell distribution and mixed cultures. 24
Cell Adhesion Rate
How rapidly cells can attach to a biomaterial can determine the success of clinical applications. An early adhesion signifies that most of the cells are concentrated into the graft with a strong attachment and therefore cells will start sooner to spread. On the other hand, late attachment implies that cells have been around the system for longer before adhering to the surface, which could decrease viability. This is of special importance when considering seeding time for constructs going into bioreactors. Seeding time should be enough to ensure maximum cell adhesion without compromising viability or the bioreactor operation.
Fibronectin enhancing effect on cell adhesion has been proved in several cell types.25,26 Initial approaches of coating dishes and analyzing cell adhesion have proved that fibronectin enhances rapid cell adhesion and spreading of mesenchymal stem cells (MSCs) with most of the works stating that fibronectin promotes early adhesion. 27 However, Athanassiou and Deligiani reported that fibronectin induces significantly stronger adhesion after long adhesion times (30–85 min), showing that this increase in adhesion strength was proportional to the adhesion time. 28 Laminin does not induce early adhesion in MSCs because early integrin α3β1 is not present in these cells and can only promote specific adhesion after 2 h. 29 Geibler et al. demonstrated that collagen type I speeds up initial adhesion of osteoblasts compared with fetal calf serum after 1 h cells flat and spread. 23 Figure 3 demonstrates the preferential affinity of MSC adherence to different proteins that are commonly used in tissue engineering.
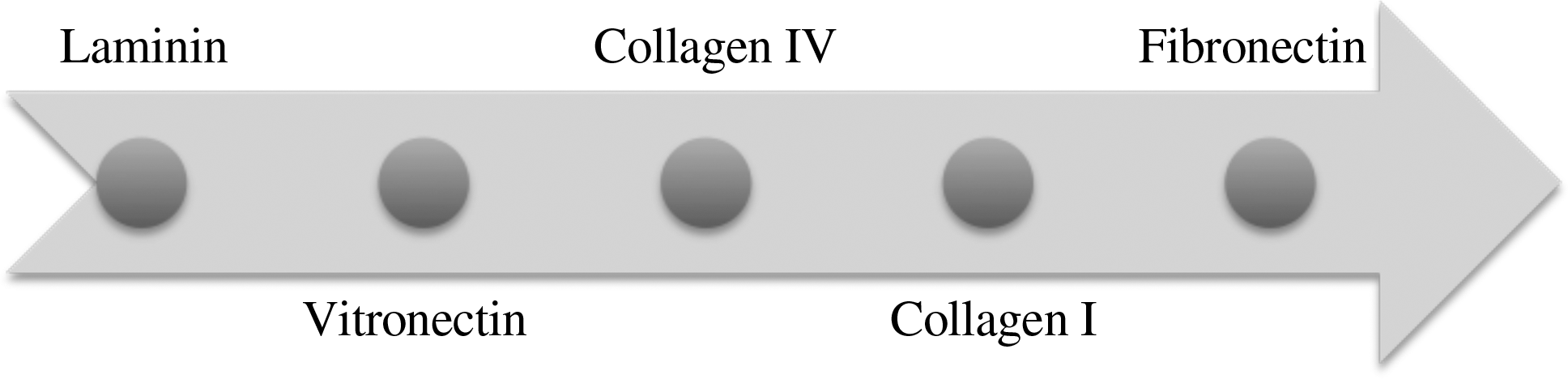
ECM affinity for cell adhesion in mesenchymal stem cells. The arrow direction indicates the best results in cell attachment when surfaces are modified with these proteins.
Osteoblast Adhesion
Surface modification techniques for osteoblasts consider growth factors (transforming growth factor-beta), bone morphogenetic protein 2, bone morphogenetic protein 7, and adhesion proteins. However, the superior adhesion is not followed up by higher differentiation or cellular functions. 30 RGD may not always improve cell adhesion because it also depends on how RGD is presented. Moreover, Garcia and Reyes describe RGD as not ideal for osteoblasts because the peptide is not as stable as the protein and osteoblasts need non-RGD integrins for differentiation.31–33
When osteosarcoma lines have been cultured in collagen type I or tissue culture plastic, cells in collagen not only reported enhanced adhesion and spreading but also showed a superior activity of the osteogenic marker alkaline phosphatase (ALP). 34 Differences between bone cell clones, osteoblastic rat osteosarcoma cell line (ROS) and nonosteoblastic RCT, both derived from the same tumor, proved that fibronectin and collagen type I were predominantly superior to laminin, collagen type IV, thrombospondin, osteopontin, osteocalcin, and bovine serum albumin in immortalized rat calvarial cell lines (RCTs). In contrast, osteopontin, fibronectin, and laminin showed higher adhesion levels in ROS. 35 Castoldi et al. reported no significant difference in acute cell adhesion between fibronectin, laminin, and collagen type I and type III in osteoblasts. 36 Moreover, Kornu et al. demonstrated that fibrinogen, vitronectin, and laminin did not have any significant effect on cell adhesion, whereas fibronectin showed a threefold increase after 2 h and no decrease in this effect after 1 day. 37
Common materials for orthopedics applications are metal alloys and ceramics. Although these nondegradable materials are not typical choices for tissue engineering scaffolds, it is important to be aware of the information relating to cell attachment on these substrates when considering cell adhesion to degradable materials. Often, similar approaches in improving the cell response to a degrading material (in particular, slow degrading materials) can be used, as have been previously identified with nondegrading materials. Initial works on cell adhesion of osteoblasts onto uncoated metallic surfaces suggested that although bone cells adhere to titanium alloys, functional bioactive surface molecules were needed to promote cell adhesion for orthopedic and dental grafts. 38 Several research groups have used ECM proteins to increase cell adhesion onto these inorganic surfaces. Dennis and Caplan used fibronectin and laminin to coat ceramics and analyzed them for 7 days. After comparing the morphology of the modified ceramics, fibronectin demonstrated a rapid spreading effect on the mesenchymal cells after just 10 min. This spreading was similar to laminin coatings only after 4 h, when most of the cells have flattened. When retention was compared, initial analysis showed no difference between the control and the two proteins. Moreover, after 4 and 7 days cell number decreased in all the cases, although protein-coated ceramics maintained twice as many cells as control ceramics. 39
When collagen type I coating was used in titanium alloys, there was no difference on cell adhesion after 1 day when compared to controls; however, osteoblastic differentiation, assessed by ALP activity and calcium content, was superior when the ECM protein was present. 40 Collagen type I effects have also been evaluated on polylactic acid (PLA) surfaces either on its own or combined with apatite to mimic the bone matrix. Enhanced MSC adhesion and morphology was showed when the PLA surface was modified, although there was no significant difference between the results of the protein alone and the combined coating. 41 Higher matrix mineralization levels were also induced by collagen type I surfaces but not with fibronectin or vitronectin. 42 These results have been used as a rationale to the use of collagen type I fibers as a biomaterial to grow MSCs. Shih et al. observed higher cell viability and improved morphology of cells cultured in collagen type I fibers than in those cultured in tissue culture plastic, whereas osteogenic differentiation was not significantly different between the two groups. 43
Effect of these proteins has also been evaluated in glass surfaces. Garcia et al. treated bioactive glass, hydroxyapatite, and borosilicate glass with fibronectin to evaluate the effect of a surface reaction treatment in the fibronectin-mediated adhesion effect. This work confirmed that fibronectin surfaces resulted in stronger cell adhesion proportional to fibronectin concentration. In addition to this, the surface treatment proved to improve the cell receptor–fibronectin affinity, suggesting that fibronectin conformational changes can affect its effect on cell adhesion.44,45
MSC Adhesion
Skeletal tissue growth and repair are determined by the MSCs, multipotent progenitor cells from bone marrow. MSCs have been differentiated in vitro into osteoblasts, chondrocytes, adipocytes, and myoblasts and now are part of the paradigm of tissue engineering to repair musculoskeletal tissues. Current front-edge therapies on cartilage, tendon, and bone investigate on the application of MSCs. 46
Most of the cell adhesion studies in MSCs agree on the superior effect of fibronectin compare with other ECM proteins (Table 2). This increased attachment ratio of cells to fibronectin is explained by fibronectin being recognized by 20 different binding receptor integrins.19,47 Zhang et al. have also investigated the effect of modifying surfaces with fibronectin for improved MSC attachment and function. 48 In their study, a novel recombinant protein of fibronectin and cadherin 11 was created and used to coat a calcium phosphate base before primary human MSC adhesion. It was found that coating surfaces with the novel FNII17-10/CDH 11 EC 1-2 fragment and MSC adhesion, proliferation, and ossification were significantly improved compared with the pure calcium phosphate surface and calcium phosphate surfaces that had been functionalized with fibronectin and cadherin 11 alone. Stem cells derived from adipose tissue have also been investigated with regard to fibronectin. 49 Results from this particular study showed that these types of stem cells have an adhesion mechanism for the heparin-binding domain of fibronectin and that their morphology is controlled by their adhesion mechanism. These data also showed a strong correlation of adhesion mechanism with adipogenic differentiation. Collagen type I provides a good substratum for cell attachment after 2 h but with no further signs of differentiation. 50 Collagen type II can promote MSCs into chondrogenesis as it triggers the synthesis of cartilage proteins. 51
Best statistically significant cell adhesion results are highlighted. Major osteogenic differentiation effect is in boldface. Cell type used is included in parentheses.
hMSC, human mesenchymal stem cell; NA, not applicable
Chen et al. have investigated the effect of type I collagen and fibronectin on MSC (C3H10T1/2) cell adherence, proliferation, and differentiation. 52 Polyacrylamide hydrogen was used to support different protein surfaces. The authors suggested from their findings that there is a “complex interplay between matrix mechanics and biochemistry in the regulation of MSC function.”
Hoshiba et al. have taken a less “protein-specific” approach to investigate and improve MSC adhesion onto substrates. 53 In their study, MSCs were first grown in tissue culture dishes to produce ECM. At various weekly time points, these samples were then decellularized. The remaining ECM then provided biological substrates for a new fresh batch of MSCs to be cultured onto. This study concluded that MSCs cultured on “osteogenesis mimicking matrices” (ECM that was produced by MSCs undergoing osteogenic differentiation) produced more runt domain factor 2 protein (RUNX2) than those cells cultured on ECM produced by undifferentiated MSCs.
The effects of ECM-modified surface are not limited to improved cell adhesion. It has been demonstrated that fibronectin, vitronectin, and collagen type I also induced a dose-dependant chemotaxis in rabbit and human MSCs, with fibronectin inducing a higher response than vitronectin and collagen type I. Haptotaxis, growth directed by substrate gradients, and motogenic activity were showed as well with MSCs producing fibronectin and collagen type I to enhance MSC migration to wound site. 54 This shows that ECM proteins used as a substrate are related to collective cell migration. 55
Klees et al. confirmed recently that fibronectin and laminin enhance adhesion in MSCs, but most importantly focused on laminin 332. This molecule elicited high activity for ALP, osteocalcin, bone sialoproein-2, and core binding factor alpha-1 but did not improve the levels of calcium when compared to the effect of osteogenic supplements in the media (dexamethasone, ascorbic acid, and β-glycerophosphate). 56
Protein Adhesion Roles in Cell Differentiation
Osteogenic differentiation markers are based on the main proteins expressed and used by osteoblasts (Fig. 2). It is a standard to use ALP (involved in the mineralization process of bone matrix), osteopontin, osteonectin, and sialoproteins to evaluate the differentiation level of a bone cell. ECM-integrin-activated signals are responsible for this osteoblastic differentiation through the initiation of the mitogen-activated protein kinase signal transduction pathway.57,58 Fibronectin and collagen have been proved to promote higher activity in this pathway contrary to vitronectin, whose osteogenic effect must be related to a different mechanism. 59 Klees et al. published that laminin induces osteogenic differentiation through the activation of the extracellular-related kinase, whereas fibronectin has no effect on this. 60 RGD has also been proved to promote osteoblast proliferation, osteo-specific gene and protein expression, and mineralization when used in solid surfaces.61,62 Although collagen type I accelerates mature osteoblast phenotype development, enhances expression of osteoprotein markers, and produces a higher ALP activity of MSCs cultured in fibronectin and fibronectin/collagen surfaces, fibronectin and collagen differentiation effects do not compare to their superior results on adhesion.23,63 Chun et al. have investigated the use of an RGD-conjugated thermosensitive polymer to promote the osteogenesis of MSCs. 64 In their study, poly(organophosphazene)-RGD conjugate was used to suspend rabbit MSCs before injection into a nude mouse, upon which the hydrogel formed as a result of exposure to body temperature. At week 4, significantly higher calcium, osteocalcin, and collagen type I were detected in the hydrogel/cell samples than in the controls.
Iba et al. explored the effect of concomitant soluble osteogenic factors and showed that the presence of dexamethasone enhances the promoting effect of fibronectin in ALP activity, whereas the use of 1α,25-dihydroxyvitamin D3 (1,25(OH)2D3) resulted in increased levels of osteocalcin in collagen type IV–coated materials. This demonstrated that ECM components have factor-dependent effect on differentiation. 65
New Approaches
As target regions are now fully sequenced, peptides have been explored to control cell adhesion. Cooke et al. compared the cell adhesion effect of peptide motifs most likely to enhance cell attachment of fibronectin, collagen types I and IV, and laminin to the results of complete proteins. Although this work did not use MSCs but PC12 cells, cell adhesion levels with specific peptides were between 12% and 80% of the adhesion levels reached by complete proteins. 66 These results are confirmed in MSCs by Hennessy et al., who compared cell adhesion on collagen type I surfaces and collagen type I peptides. Cell adhesion and spreading was lower on peptides' surfaces than on the complete protein surface. 67 Hennessy et al. also demonstrated in this article that MSCs adhere significantly better to hydroxyapatite surfaces that are coated with RGD, Asp–Gly–Glu–Ala sequence (DGEA), and P15 (integrin binding factors). The collagen-derived peptides, DGEA, and P15 enhanced the osseointegration of hyaluronic acid implants, but their roles for improvement were of cell differentiation promotion rather than cell adherence. There has been no report published regarding the identification of which structure portion (secondary vs. tertiary) is required for MSC adhesion to surfaces.
Another approach is the use of adhesion microarrays. These arrays for testing cell adhesion with specific peptides and proteins have the advantages of minimal use of protein, multiple cell types, and easy analysis under the microscope. First-generation or two-dimensional ECM arrays are further investigated to develop 3D microarrays resembling in vivo microenvironments.68–70 The goal of this technology is to identify the best molecule for cell adhesion and differentiation to use the spatial control of these factors through micropatterning to direct cell adhesion, distribution, and differentiation. 71 Next-generation biomaterials are predicted to be nano-engineered 3D structures able to support specific cell adhesion and differentiation.
Conclusion
ECM proteins' effect on MSC adhesion decreases in the following order: fibronectin, collagen type I, collagen type IV, vitronectin, and laminin, showing that fibronectin promotes more cell adhesion than any other ECM protein.29,72 Differences between osteogenic differentiation in fibronectin are attributed to cell type. Differences of ECM affinity on cell adhesion between osteoblasts and MSCs are caused by the use of αv and α2 integrins in MSCs, while osteoblasts use αv and α5 integrins. 73 Although fibronectin greatly induces osteogenesis in osteoblasts, it has minimal effect on MSCs, where osteogenesis is induced by collagen type I and laminin. 29
General consensus of the ECM affinity for cell adhesion has been reached (Table 2); however, the optimal ECM coating for osteogenic differentiation is still not clear. Further studies on this are conducted, focusing in combining two different ECM proteins in the coating, one for cell adhesion and another for differentiation improvement; enhancing bone cell adhesion and differentiation will allow tissue engineering to develop functional osteo-constructs with the ultimate goal of clinical skeletal regeneration.
Footnotes
Acknowledgments
Financial support was provided by the Biotechnology and Biological Sciences Research Council (BBSRC; grant reference code BB/F013892/1).
Disclosure Statement
No competing financial interests exist.