
Abstract
Articular cartilage (AC) has no or very low ability of self-repair, and untreated lesions may lead to the development of osteoarthritis. One method that has been proven to result in long-term repair or isolated lesions is autologous chondrocyte transplantation. However, first generation of these cells' implantation has limitations, and introducing new effective cell sources can improve cartilage repair. AC provides a resilient and compliant articulating surface to the bones in diarthrodial joints. It protects the joint by distributing loads applied to it, so preventing potentially damaging stress concentrations on the bone. At the same time it provides a low-friction-bearing surface to enable free movement of the joint. AC may be considered as a visco- or poro-elastic fiber-composite material. Fibrils of predominantly type II collagen provide tensile reinforcing to a highly hydrated proteoglycan gel. The tissue typically comprises 70% water and it is the structuring and retention of this water by the proteoglycans and collagen that is largely responsible for the remarkable ability of the tissue to support compressive loads.
Introduction
Repair is the rapid process to resolve an injury. The reparative tissue is not identical to the original tissue and there is no integration of repair tissue with the original tissue. Regeneration is a relatively slow process that recapitulates development and morphogenesis and restores completely structure and function, including integration of the new tissue seamlessly to the original.
Regenerative medicine is the emerging discipline of medicine based on advances in basic science of development and morphogenesis and science of biomaterials and stem cell biology. The three key ingredients for regenerative medicine and surgery are the inductive signals such as bone morphogenetics proteins (BMPs), responding cells, and the scaffolding of extracellular matrix (ECM).5,6 Regenerative medicine is governed by biology, bioengineering, and biomechanics.
The embryonic development and morphogenesis of cartilage is initiated and regulated by BMPs. 5 The cartilage-derived morphogenetic proteins are critical for AC differentiation and the complete joint morphogenesis.7,8 BMPs and cartilage-derived morphogenetic proteins were isolated from bone and cartilage, respectively. The rules of architecture for regenerative medicine and surgery are an adaptation and imitation of the rules of development biology and morphogenesis and may be generally universal for many tissues. 5
The AC is adjacent to the subchondral bone. Yet, there is a fundamental difference between the regenerative response of bone and AC. Bone has supreme regenerative capacity. On the other hand, AC is recalcitrant and feeble in its capacity for regeneration and even repair. What is the cellular and molecular basis for these distinct differences of regenerative potential in these two adjacent and related tissues? First, unlike bone, cartilage is avascular and is devoid of nerve supply. Second, as they lack vascular supply there are no immediate early repair responses with monocytes and macrophages to injury. Recent work on mesenchymal stem cells (MSCs) demonstrated that blood vessels and associated pericytes support tissue regeneration and homeostasis. 9 Perhaps the absence of vasculature and attendant lack of pericytes may explain in part lack of regeneration.
The AC has very distinct anisotropy and distinct polarity. The superficial zone has flattened chondrocytes. The surface zone secretes the superficial zone protein (SZP) also known as lubricin.10–15 The middle zone chondrocytes secrete collagen II and the proteoglycan aggrecan. The deep zone of AC has a mineralized ECM with the distinct tidemark, on the subchondral bone. The distinct functional zones of surface, middle, and deep has to be faithfully regenerated with complete fidelity. This is the main challenge in AC regeneration. The next section will discuss the structure of the AC with special emphasis on collagen fibrils orientation in each of the three zones of AC.
Structure of AC Collagen Fibrils: Arrangement, Polarization, and Distribution
The ECM of the AC is a specialized connective tissue consisting of a hydrated proteoglycan gel that resists compression reinforced by a network of collagen fibrils. Collagen is an important component of the AC, representing around 50% of its dry weight and being the most important constituent to provide tensile strength. Nevertheless, its distribution and organization remains a matter of controversy. Perhaps the strong interaction with proteoglycans and the high level of hydration presents challenge to observe the matrix by classical optical microscopy. Recent technological improvements have contributed to our increasing understanding of cartilage organization.
The pioneering work of Benninghoff established the concept that collagen fibrils are oriented vertically in the deeper layers of AC, 13 twisting into arches at the intermediate layers, and assuming a horizontal disposition in the superficial layer. 15 Benninghoff's concept has ever since received wide support especially by investigators using polarization microscopy. In the last century, several authors, using different microscopic methods, postulated specific orientation for collagen fibrils in AC. However, the methodological limitations and the intricate organization of collagen fibrils continue to be challenging.16–19
The organization of several cartilages, including AC, were investigated using tissue sections, Picrosirius red staining, enzymatic digestions, and polarized microscopy. 20 Picrosirius-polarization method has an advantage over other methods because it increases the resolution of light microscopy due to the increase in natural birefringency of collagen fibrils when they bind the Picrosirius Red dye. Colored birefringency is enhanced against a dark background, allowing the distinction of usually undetected thin fibrils. 21 Enzymatic digestion using hyaluronidase removes proteoglycans and unmasks collagen fibrils rendering them more accessible and therefore better stained. 22 Using such methods several authors found for AC in several species similar fibril distribution to the original Benninghoff proposal: single gothic arches made by vertically oriented fibrils in the deeper zone reaching osteochondral boundary and horizontal orientation in the superficial zone. Similar collagen fibril distribution has been reported by Nieminen et al. using quantitative magnetic resonsnce imaging and polarized light microscopic study in bovine AC. 23
The three-dimensional structure of collagen in bovine AC was discovered with scanning electron microscopy using a modification of a technique.24,25 Enzymatic digestion of the proteoglycans defined the underlying collagen structure but was incomplete to maintain tissue integrity. In the middle and superficial zones, collagen was organized in a layered or leaf-like manner. The orientation was vertical in the intermediate zone, curving to become horizontal and parallel to the articular surface in the superficial zone. Each leaf consisted of a fine network of collagen fibrils. Adjacent leaves merged or were closely linked by bridging fibrils and were arranged according to the split-line pattern. The surface layer (lamina splendens) was morphologically distinct. As collagen is an integral component of cartilage matrix its organization is critical for cartilage formation, growth, repair and regeneration. 24 Following a similar pattern of collagen distribution in rabbit and in human were described novel aspects from cryo and modified chemical preparation techniques for scanning electron microscopy.26,27 They focused the studies on the radial zone where a special distribution of collagen fibrils forming columns in the regions surrounding the rows of chondrocytes, the so-called chondrons. These columns of 1–3 μm diameter each, have densely packed collagen fibrils. These fibrils were arranged radially; some were straight of 30 nm and others in an opposed spiral arrangement of 10 nm, with regularly repeating patterns. The aggrecan component of the ECM could be contained in such columns. The load bearing property of the tissue was explained by the directed flow and containment of the interstitial fluid, modulated by the protein–carbohydrate complexes, along collagen bound tubular structures. The possible reason why such structure was not described earlier may be that it is not preserved by aldehyde fixation followed by dehydration, the method commonly used for tissue preparation for electron microscopy.
There is no unanimity about the diameter of the collagen fibrils; they ranged in diameter between 30 and 110 nm. 20 Type IX collagen interacts with collagen II and other IX fibers. 25 In addition, there is potential crosslinking between collagens II and IX. 27 The collagen IX binds to collagen II.28–30 A similar D-periodic banding has been observed in chick embryo sternal and bovine AC.29–31
Rieppo et al. observed birefringence, orientation, and parallel collagen fibril network after autologous chondrocyte transplantation. 31 The repair tissue lacks the typical collagen network organization of AC. Collagen fibril orientation is parallel to the surface throughout the entire thickness of cartilage and the normal phenotype of AC is not achieved. Changes in the collagen architecture and spatial collagen content during AC growth and maturation in pig showed classic Benninghoff architecture. Perhaps one possible explanation for the appearance of cartilage is adaptation to joint loading. The finding in pig may have significance to the human. Shirazi and Shirazi-Adl 32 have investigated in human knee joint the role of deeper vertical fibrils of collagen in AC mechanics through finite element methods. They hypothesize that those fibrils play a crucial role in cartilage mechanics by supporting and protecting the tissue under physiological loading conditions.
The surface of the superficial zone is stained by Picrosirius Red more strongly than the interior (Fig. 1). The surface region fibrils are oriented parallel to the surface and the parallel fibers appear to intersect. The polarization optics permits the observation of birefringence in gradation in both positions of polarization going from less in one position to more when the specimen is turned 45° left or right. The inner areas of superficial zone are initially dark and increases birefringency with turn of the slide. After papain treatment to remove proteoglycans there is a generalized increase in birefringency as a consequence of unmasking collagen fibrils. In the middle zone increased birefringency is observed with polarized light. The territorial matrix in the pericellular areas demonstrated weak birefringence under polarized light. The appearance of the cells after papain treatment decreases metachromasia and the territorial matrix shows the appearance of a Maltese cross, which move around the cells when the slide turned, which demonstrating a heterogenous collagenous organization. The interterritorial zone exhibits thicker collagenous fibrils with an oblique pattern criss-crossing at right angles to each other. The schematic depiction (Fig. 2a) demonstrates that the fibers are parallel in the surface and in the layer close to middle layer the oblique collagen fibers interspersed within vertical regions. Similar fibril organization also is observed in the deep zone. In the course of repair one observes a disorganized fibril organization (Fig. 2b). Ideally, one would like to observe complete regeneration with optimal fibril orientation (Fig. 2c).
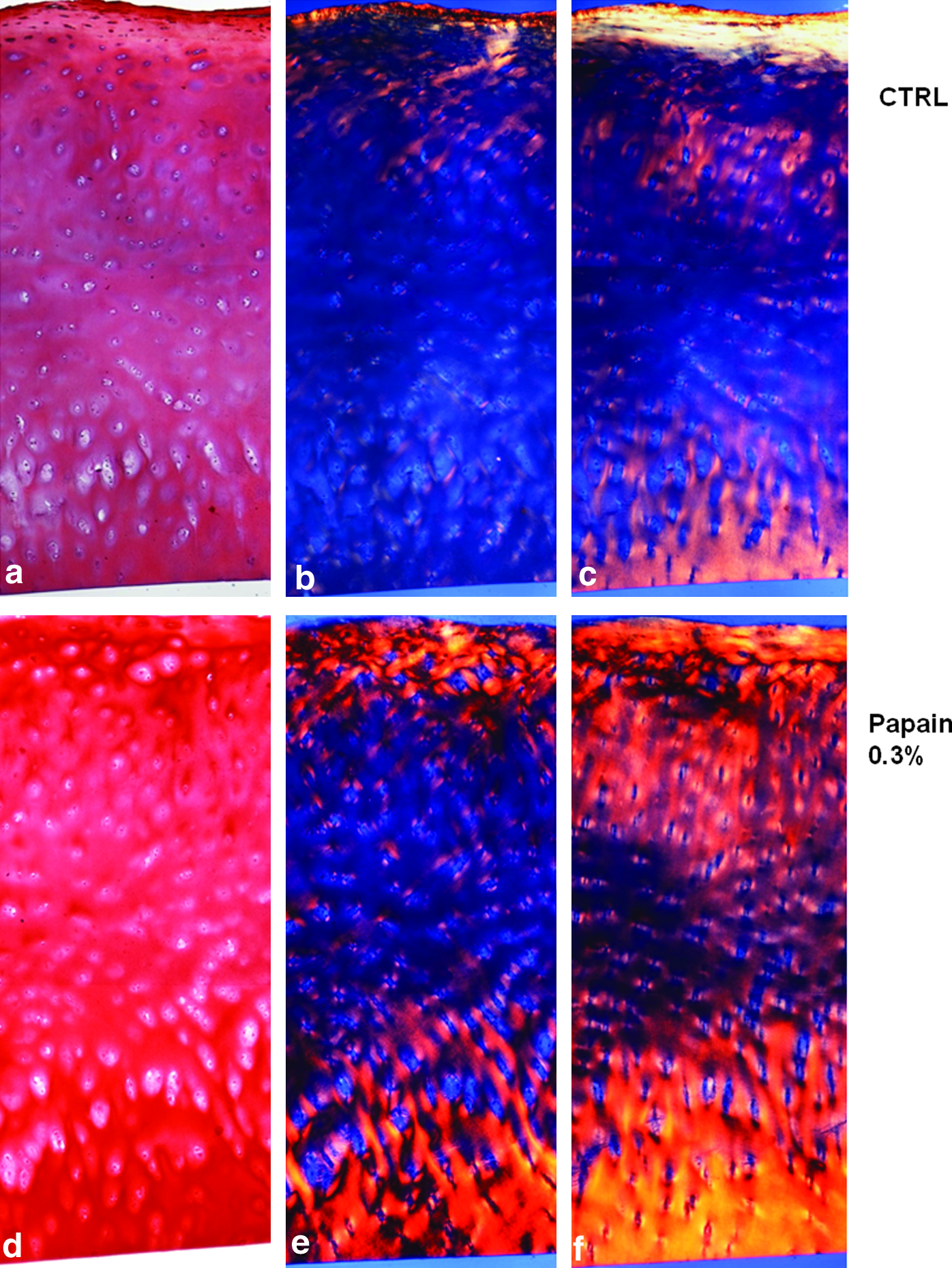
Structure of human articular cartilage. Human articular cartilage sections were stained with Picrosirius and hematoxylin.

A schematic description of the collagen fiber orientation in the human articular cartilage. The normal structure
As collagen is an integral component of cartilage matrix, its organization is critical for cartilage formation, growth, repair, and regeneration.
Transforming Growth Factor-βs' Family and Cartilage Zonal Organization and Differentiation and Metabolism
The AC has a distinct zonal organization that is intimately linked to function. The superficial zone cartilage (SZC), the middle zone cartilage (MZC), and the deep zone cartilage (DZC) are generally identified in AC. 33 The AC in addition has a calcified cartilage zone that has mineralized ECM and is distinguished at the interface with DZC by the so-called tide mark, which in most histological stains such as toluidine blue has different staining properties from the metachromatic middle and deep zones (MZC and DZC). The entire AC is intimately associated with subchondral bone by the most characteristic interdigitation of the calcified cartilage zone into the bone.
The middle and DZC has the collagen II and the proteoglycan aggrecan and several noncollagenous proteins, including but not limited to cartilage oligomeric matrix protein and cartilage intermediate larger protein. 34 It is important to keep in mind that the superficial, middle, and deep zones are a continuum, and therefore there are an expected transitional areas between the SZC, MZC, and DZC. The functional aspects of the various zones in the AC are an important consideration in exploring the AC regeneration.
SZP is secreted by the SZC and is a mucinous glycoprotein with a covalently attached proteoglycan chain. The SZP is homologous to lubricin, a glycoprotein secreted by synovium and first purified from synovial fluid. SZP/lubricin is secreted by both the superficial zone chondrocytes and synovium, and therefore the relative regulation of these cellular sources by morphogens and growth factors is critical. 35 Lubricin/SZP is known to function as a boundary lubricant in diathroidal joints and plays a role in reducing the coefficient of friction in the opposing gliding surfaces or AC in all joints. SZP and lubricin are encoded by the same gene, proteoglycan 4 (prg4). Mutation in the prg4 gene has been attributed to the Camtodactyly-arthropathy-coxa-vara-pericarditis syndrome.34,35 A key feature of this syndrome is alterations in articular surface and attendant degradation of AC and noninflammatory early onset joint failure. Thus, the functional importance of SZP/lubricin is demonstrated by the pathophysiology of the joints in the Camtodactyly-arthropathy-coxa-vara-pericarditis syndrome.
Why focus on the surface of AC in arthritis? The first changes at the onset of OA occur at the superficial zone of AC.36–41 There is loss of cellular SZP immunostaining in degenerative AC in menisectomized sheep model of early OA. 42 In addition, there is a strong association between the loss of boundary lubrication and damage of AC in an experimental model of OA in rabbits induced by transection of anterior and posterior cruciate ligaments. 43 Therefore, SZP plays a critical role in joint physiology and is an attractive target for systematic investigations especially of the regulatory biology of the superficial zone of AC.
Regulation of SZP accumulation is critical not only for homeostasis and maintenance of AC, but also for the tissue engineering and regenerative medicine of functional AC. SZP accumulation by superficial zone chondrocytes was enhanced by transforming growth factor-β (TGF-β) in bovine cartilage. BMP-7 stimulated the accumulation of SZP in both explant cultures of SZC from bovine AC and in monolayer cultures of chondrocytes derived from SZC. 44 In view of this, a systematic study of TGF-β/BMP superfamily members was conducted on the responses of cells from SZC and synoviocytes in calf cartilage. 45 The BMP/TGF-β superfamily includes BMPs, TGF-β, growth/differentiation factors, and activins. TGF-β isoforms 1, 2, and 3 were potent in stimulating SZP secretion by both superficial zone chondrocytes and synoviocytes. 46 SZP is mainly secreted by SZC but not middle and deep zones even in the presence of TGF-β. The response of superficial zone chondrocytes and synovium to TGF-β isoforms is dose dependent and is biphasic. High dose (30 ng/mL) was inhibitory compared to the optimal dose (1–3 ng/mL). The biological actions of TGF-β isoforms are dependent on TGF-β receptor I kinase as specific inhibitors of the kinase blocked the response. 45 Members of the BMP family (BMP-2, 4, and 7, and growth/differentiation factor-5) demonstrated a distinct differential response in SZC chondrocytes and synoviocytes in the bovine joint. The synoviocytes were much more responsive to BMPs. Similarly, synoviocytes were more sensitive to activins A, B, and AB compared to chondrocytes from the superficial zone. These observations demonstrate the differential regulation between superficial zone and synoviocytes in the joint, indicating the differences among the various compartments of the joint such as AC and synovium. In addition, there may be a division of labor among the BMP/TGF-β superfamily; the superficial zone is more responsive to TGF-β isoforms, whereas the middle zone is mainly regulated by BMPs.
The function of AC is intimately linked to biomechanics. Mechanical loading is critical for homeostasis of musculoskeletal tissues, including AC. AC performs the biomechanical function of load support and lubrication with minimal wear and little or no damage in animals and humans. Dynamic shear stimulates SZP. The molecular basis of mechanotransduction in AC is beginning to be understood. The SZP expression pattern is dependent on the geometry of the femoral condyle surface and anatomical location. The anterior sites secreted more SZP in both lateral and medial femoral condyles. The biomechanical assays of SZP accumulation were confirmed and corroborated by immunolocalization of SZP in the various anatomical locations of the lateral and medial femoral condyles. 47 Again, the highest SZP were in anterior region compared to the posterior sites. The distribution of maximum contact pressures was associated with high SZP content. Anatomical regions of high contact pressure were consistently located in the anterior region. In addition, shear loading of the anterior medial condyle increased significantly SZP secretion compared to posterior sites of the same femoral condyle. The SZP response to shear-loading was abolished in the presence of TGF-β type I receptor kinase inhibitor, demonstrating the critical role of TGF-β signaling in mediating mechanotransduction. 48 Thus, these experiments demonstrate a new role for TGF-β signaling pathway in joint lubrication and is mediated by cellular mechanotransduction in superficial zone of AC. The middle and deep zones were refractory to shear loading, further illustrating that there apparently a gradient of response originating from the surface to deep zones. The SZC is therefore critical in mechanotransduction and in joint lubrication. The science of tribology deals with lubrication and wear in various surface boundary lubrication regimens. A new field of biotribology has emerged dealing with biological surfaces in sliding contact combining concepts of friction, wear, and lubrication of opposing interacting surfaces such as the gliding AC surfaces in the joint. 49 Thus, the emerging new findings in AC are also critical for regeneration and restoration of damaged AC in OA by tissue-engineered cartilage. It is critical that attention be focused on the characteristics of low coefficient of friction and high resistance to wear in regenerative medicine and tissue engineering of AC.
Cell Therapy for Cartilage Regeneration
During the last decade there has been an exponential increase in research activity in the field of cartilage tissue engineering. AC is seen as an ideal candidate for a tissue engineering approach to tissue regeneration.
Trauma to the AC surface of the joint represents a challenging clinical problem because of the very limited ability of this tissue to self-repair. A number of surgical protocols are currently in use for the treatment of AC defects. In abrasion arthroplasty, 50 for instance, the subchondral bone is perforated by drilling to promote bleeding into the defect, with the result that there is formation of bone and fibrous repair tissue. In the microfracture technique 51 the exposed subchondral bone is microfractured (“picked”) to promote localized bleeding. Moreover, repair techniques such as microfracture, which introduce cells into the joint, have unpredictable clinical outcomes as they produce a fibrocartilage tissue that degenerates with time. Another approach involves the use of allografts 52 where cartilage lesions are filled with grafts of donor-derived osteochondral fragments. However, these procedures are restricted by the availability of suitable donor tissue. In mosaicplasty 52 cylindrical osteochondral plugs are harvested from nonload-bearing sites in the affected joint and pressed into place within the osteochondral defect, creating an autograft “mosaic” to fill the lesion. Autologous chondrocyte implantation (ACI) therapy 52 represents another approach, where a chondral biopsy is taken from a donor site at the time of clinical examination. One of the major relevant issue in this field as been the recent acceptance of ChondroCelect® (by TiGenix) as a cell-based medicinal product consisting of chondrocytes that are taken from a healthy region of the patient's cartilage, grown outside the body, and then re-implanted during an ACI surgical procedure. The ACI procedure with a membrane of collagen (collagens I and III) is called MACI. Chondrocytes, enzymatically released from the retrieved tissue, are expanded in monolayer culture, and subsequently implanted in a second procedure beneath the periosteal membrane or the MACI membrane, which is sutured to the cartilage adjacent to the defect and sealed with fibrin glue. All of these approaches offer exciting opportunities for the regeneration of cartilage defects. However, the long-term outcome may be uncertain and there are may be other disadvantages associated with the harvest site, even when it is some distance from the lesion. In this regard, nowadays it is well known that lesion size, activity level, and age were the influencing parameters of the outcome of AC repair surgery. 53 Lesions >2.5 cm2 should be treated with ACI or osteochondral autologous transplantation, whereas microfracture is a good first-line treatment option for smaller (<2.5 cm2) lesions. Patients who are active show better results after ACI or osteochondral autologous transplantation when compared with microfracture. In this regard, studies discussed by Knutsen et al. conclude that both ACI and microfracture provide satisfactory results in 77% of the patients at 5 years with no significant difference in the clinical and radiographic results. 54
However, because there are limitations, these have been the driving force behind the emerging field of AC tissue engineering, whose approaches may provide an ideal alternative to the current surgical treatments for cartilage repair. Implantation of cells with both chondrogenic and osteogenic potential derived from the bone marrow for the treatment of osteochondral lesions represents another approach that might result in persistent, functional restoration of the AC. Chondrogenic cells that are in more abundant supply can be used for cartilage tissue engineering. Identification of a suitable cell population for cartilage tissue regeneration is the critical first step in the process. Articular chondrocytes are the most obvious choice. To obtain large numbers of allogeneic chondrocytes for culture of autologous cartilage constructs, it is necessary to be able to expand the cells in culture. Different groups have carried out experiments to assess whether ovine chondrocytes could be expanded in monolayer and retain their ability to produce cartilage in the culture system. 55
Aging of the cells is an issue when these autogenous cell-based techniques are applied to aged people. Chondrocytes obtained from the aged have much lower ability to repair cartilage than young ones. 56 For cartilage regeneration in aged people, using cell type with greater potential, such as bone marrow stromal progenitor cells, may be perhaps more promising, 54 as it has been demonstrated recently where age did not made a difference in outcomes in the use of bone marrow-derived MSCs in patients older than 45 years. 57 There are also a prevalence in the use of MSCs from synovial origin rather than bone marrow in terms of chondrogenesis as it has been demonstrated by several authors with porcine 58 and human multipotent MSCs isolated from the synovial membrane of knee joints.59,60 These cells may be superior as a potential source of MSCs and play a role in the regenerative response during OA. 61 More recently, attention has been focused on human embryonic stem cells as they have the ability to self-renew and differentiate into any cell lineage of the three germ layers, therefore holding great promise for regenerative medicine applications. In this term, Toh et al. describe a micromass culture system as a model system to study chondrogenic differentiation and modulate cartilage-specific matrix gene expression in a distinctive manner. 62
The development of effective therapies for OA has been slow, and today most recommended medications alleviate symptoms without altering the course of the disease. In many cases, joint replacement surgery is the best option to restore joint function. The observations made by several authors 63 on the characteristics of stem cells from OA patients indicated that stem cell therapy might be an effective approach in impeding the degenerative changes in the OA joint. A number of studies have been carried out involving the delivery of stem cells to the goat knee joint following medial menisectomy, resection of the anterior cruciate ligament, or a combination of the two. 64 This procedure, in combination with a defined exercise regime, leads to the development of lesions in the joint that are characteristic of OA. Transduced cells with green fluorescent protein have been injected and the general approach was to deliver the cells as a suspension by intra-articular injection, thus avoiding the need for arthrotomy and the placement of a scaffold. 65 Injected cells were retained within the joint and also recovered from synovial fluid in a viable form. The cells colonized soft tissue surfaces, primarily those of synovial origin rather than the AC. Injection of MSCs into destabilized, osteoarthritic joints resulted in marked remodeling of the medial meniscus, which had been totally removed during surgery. 66 The new tissue that formed has a hyaline-like appearance and focal areas of type II collagen similar to developing rabbit meniscus. Related to this tissue regeneration is a marked chondroprotective effect of the MSCs injection. These observations highlight the potential therapeutic benefit of injected MSCs in an osteoarthritic joint by regenerating neomeniscal tissue to stabilize the joint and protect the articular surfaces against progressive degeneration. 67 Wakitani and others show the first clinical trial using autologous culture-expanded bone marrow MSCs to repair AC. 68 Subsequently, they performed this procedure in about 40 patients demonstrating the safety of the procedure. Later on, they demonstrated improvement in clinical symptoms with a hyaline-like type of cartilage tissue in young, active patients, 69 although the repair cartilage was not hyaline cartilage in normal age individuals. To regenerate AC by cell transplantation, it is essential that cells proliferate without losing their capacity for differentiation. To find appropriate conditions, different culture conditions, mechanical stresses, growth factors, and gene transfection have all been explored, but these have not yet been applied clinically. 70
One might consider the phenotypic plasticity as an in vitro artefact as the chondrocytes are being exposed to and artificial environment and being extensively modified during their expansion in monolayer. However, plasticity between different phenotypes is a common phenomenon during embryonic development of tissues and organs. Using this feature, the same cells can be reprogrammed to serve several functions. Also, the plasticity is preserved and has critical functions in adult amphibians such as salamanders, as observed in the regeneration of an amputated limb. 71
In vivo, plasticity is mainly seen during the embryonic development of organs and tissues, for example, the shift from endothelial-like cells to mesenchymal cells during the embryonic formation of synovial joints. This change is made from the cells that will form the AC, the interzonal cells. Initially, these cells have characteristics similar to the endothelial cells lining blood vessels and, after condensation, will round up and start to secrete cartilage matrix proteins. The shift from endothelial to mesenchymal cells has further been shown to persist in the formation of adult heart valves. 72
It is a well-established fact that during monolayer expansion of chondrocytes in vitro, the cell population losses its phenotype and starts to express primitive embryonic markers.73,74 This dedifferentiation process is not unique for chondrocytes but a rather common phenomenon observed in many types of cells and tissues.75–77 Another possible explanation to the change in expression during the expansion is that the culture conditions are favoring certain cell subpopulations, for example, transit-amplifying cells, and that these cells are able to respond to the culture conditions more rapidly.
During redifferentiation of dedifferentiated adult chondrocytes expanded in human serum, chondrocytes do express genes that have been described in the initiation of chondrogenesis during limb formation. 77 This shows, in addition to the phenotypic plasticity,77,78 that the cells revert to a primitive stage during the expansion in vitro. Culture-expanded chondrocytes are able to redifferentiate when replaced into three-dimensional environment under appropriate culture conditions.79–81 This ability of redifferentiation is particularly important when the cells need to be transplanted into patients. To better define the chondrogenic phenotype and to try to optimize the cellular recovery for clinical cell transplantation, several attempts have been made to characterize culture-expanded chondrocytes at the molecular level.80–84
During the expansion of human chondrocytes, the degree of dedifferentiation has been correlated to the number of cell division or passages.73–75 This was a typical characteristic demonstrated in the study of myogenic differentiation of chondrocytes. 74 In that study, no differentiation into muscle was obtained when cells from a low passage were used, whereas with later passages, cartilage-derived muscle fibers were obtained, confirming the amplification of adult articular chondrocytes in vitro results in a population of cells with progenitor properties.
Biomaterial scaffolds provide the chondrogenic cells with a microenvironment, where they survive, multiply, and produce ECM to constitute regenerated cartilage. Although the cellular products are expected to replace the degradable biomaterial, the process is usually time-consuming and the scaffold should be implanted before completion of the process. The biomaterials thus play the role of a vehicle to transfer cells and therefore should be compatible with the native tissue around the recipient site. 76 Many natural substances are suitable as the cell-carrying scaffold for cartilage engineering, including fibrin, agarose, alginate, collagen, chitosan, and hyaluronan. Many of these are hydrogels and can be designed as injectable in their liquid form, which blends well with chondrogenic cells. 77 After being injected into the recipient site, they set by gelation to fill in any shape and size of cartilage defect.
Either chondrocytes or stem cells are used to constitute engineered cartilage useful to regenerate damaged tissue, and in vitro manipulation of the cells is necessary in most of the currently available systems. When the constructed cartilage tissue is considered for clinical used, the safety of the whole process needs to be debated and the cost is high. The entire process has to be conducted with expensive laboratory facilities that meet the high standard of good tissue practice. In addition, all reagents involved in the process should be proven as safe for human use. More complicated manipulation of the cells will arouse more concern that the cells may be affected in unknown ways. When developing a system to regenerate cartilage for clinical application, one should always consider the safety and efficacy of the regenerated tissue.
As a result of the variable and unpredictable clinical experiences in cartilage regeneration in the past, biotechnology has been introduced to this field for evidence-based development of a solution. The knowledge to date supports that AC is best repaired with autologous engineered cartilage, and a considerable research has been carried out to improve cartilage regeneration. Although the efficacy of regeneration has much improved in the laboratory and animal studies, most findings have not been investigated for their clinical safety and performance. Further studies should highlight their clinical relevance to facilitate the development of products applicable to humans.
One needs to organize currently available knowledge to develop clinically applicable models of cartilage regeneration, on the basis of autogenous chondrogenic cell implantation. A clinically applicable model of cartilage regeneration should be safe, efficient, and simple. It can be completed in a single seed-and-implant surgery procedure, which decreases the surgical risks and complications from repetitive operations of conventional autologous chondrocytes implantation. If the site of repair allows an arthroscopic approach, the surgery can be done in a minimally invasive manner within a short time, estimated at 1 h. By avoiding the complex treatment of the autogenous cells in vitro, the safety of the procedure can be improved and the cost reduced.
The Future: Challenges and Opportunities
The foregoing discussion of the structure of the AC, the zonal organization, the regulation and mechanotransduction, and joint lubrication sets the stage for contemplating the future challenges and opportunities for regeneration and tissue engineering of AC.
This brief review has by design focused on structure, fibril-orientation and the cell shape, TGF-β signaling pathways, and mechanotransduction. It is well known to most students and practioners of tissue engineering that it requires triad of signals, stem cells, and scaffolds. 9 In future, there will be continuous refinement of the scaffolds for the tissue assembly during tissue engineering.
The native AC is a durable tissue that, unless ravaged by arthritis, lasts a life time in humans. The AC is an engineering marvel in the human body. It is truly outstanding that the ECM of cartilage has predominantly collagen II providing tensile strength and the electronegative proteoglycan aggrecan, which permits movement of water with the matrix to ensure electroneutrality. This represents an advantage for an efficient function with a minimal energy expense.
AC has significant biomechanical properties, including compressive modulus of 0.79 MPa, a shear modulus of 0.69 MPa, and a tensile modulus with a range from 0.3 to 10 MPa. 78 Thus, a dynamic hydrogel of cartilage ECM is critical for the biochemical, cellular, and biomechanical function. The recent advances in nanomaterials bodes well for nanoscaffolds for AC regenerative medicine. Therefore, we enter the realms of nanomaterials and nanomedicine for the benefit of patients with OA desperately looking for solutions for the painful disease. 79 Thus, regenerative medicine of AC and tissue engineering presents critical challenges for the future and presents outstanding opportunities.
Cells to be used for an efficient regenerative medicine should be chosen. Undifferentiated versus differentiated or predifferentiated chondrocytes will be the choice. A permanent solution will come when the new tissue built in the defect is of the same nature and is perfectly integrated in the whole structure in any pathology and in any age. Only in such s way structure and function will be fully recovered.
In conclusion, the recent advances in biotechnology, nanotechnology, and nanomaterials bode well for an optimistic and bright future for regenerative medicine of AC.
Footnotes
Acknowledgments
The authors thank P. Jiménez-Palomo for his excellent technical assistance. This work was supported by grants from the Banco Bilbao-Vizcaya-Argentaria Foundation (FBBVA, Chair in Biomedicine 2007 to A.H. Reddi), the Ministry of Science and Technology (BIO2009-13903-C02-01), the Ministry of Science and Innovation (FIS PI06/1855, PLE2009-0163, FIS PI10/2529), Red TerCel (Institute of Health Carlos III), and the Andalusian Autonomous Government (P07-CVI-2781, PAIDI, and BIO-217).
Disclosure Statement
No competing financial interests exist.