
Abstract
Stem cells possess the unique capacity to differentiate into many clinically relevant somatic cell types, making them a promising cell source for tissue engineering applications and regenerative medicine therapies. However, in order for the therapeutic promise of stem cells to be fully realized, scalable approaches to efficiently direct differentiation must be developed. Traditionally, suspension culture systems are employed for the scale-up manufacturing of biologics via bioprocessing systems that heavily rely upon various types of bioreactors. However, in contrast to conventional bench-scale static cultures, large-scale suspension cultures impart complex hydrodynamic forces on cells and aggregates due to fluid mixing conditions. Stem cells are exquisitely sensitive to environmental perturbations, thus motivating the need for a more systematic understanding of the effects of hydrodynamic environments on stem cell expansion and differentiation. This article discusses the interdependent relationships between stem cell aggregation, metabolism, and phenotype in the context of hydrodynamic culture environments. Ultimately, an improved understanding of the multifactorial response of stem cells to mixed culture conditions will enable the design of bioreactors and bioprocessing systems for scalable directed differentiation approaches.
Introduction
Stem cells respond to a variety of environmental cues in vitro to either maintain potency or regulate differentiation; these cues include biochemical factors (both exogenous and endogenous), cell–cell interactions, cell–matrix interactions, and mechanical stimuli. Mechanotransduction of fluid shear stress has been studied in developmental and pathological contexts due to the induction of physical, biochemical, and epigenetic cellular responses. Hemodynamic forces are important for the regulation of cardiac morphogenesis in developing embryos, where altered flow patterns result in cardiac defects.3,4 High wall shear stresses (75 dyn/cm2 at 4.5 days postfertilization) have been measured within developing cardiac structures in vivo, 3 and can be correlated to specific patterns of gene regulation in endocardial tissue. 5 Conversely, the development of atherosclerosis in adult vessels has been linked to regions of decreased shear stress (<4 dyn/cm2). 6 As a result, the impact of fluid shear on endothelial cells has been studied extensively (extensively reviewed elsewhere 7 ). In response to physiological magnitudes (5–15 dyn/cm2) of laminar shear stress, endothelial cells in vitro exhibit morphological changes, orient along the axis of applied flow, and remodel stress fibers.8,9 Altered gene expression, as well as release of nitric oxide and other substances involved in vasoregulation, also result from endothelial cell exposure to fluid flow.10,11 Cell metabolism is altered in the presence of turbulent flow patterns, which induce cell turnover and proliferation due to loss of contact inhibition. 12 Endothelial progenitor cells similarly exhibit increased proliferation and differentiation in response to flow. 13 More recent work has indicated the potential for fluid shear stress to directly alter stem cell differentiation pathways. Mesenchymal stem cells (MSCs) differentiation along the endothelial lineage can be promoted in response to fluid flow in vitro within a parallel plate system. 14 Similarly, embryonic stem cells (ESCs) cultured in the presence of fluid shear stress in monolayer exhibited increased expression of endothelial and hematopoietic markers.15–17 The mounting evidence for fluid shear stress induced modulation of stem cell phenotype and function in adherent monolayer format strongly motivates investigation of the response of three-dimensional (3D) stem cell culture to hydrodynamic environments.
Scalable Culture of Stem Cells
Stem cells
ESCs, derived from the inner cell mass of blastocyst stage embryos, were first isolated from mouse embryos,18–20 followed by the establishment of ESC lines from primate21,22 and eventually human23,24 sources. ESCs are characterized by unlimited self-renewal and pluripotent differentiation potential into all three germ layers—mesoderm, endoderm, and ectoderm—as well as into germ cells. Cells derived from murine (mESC) and human (hESC) sources share many transcriptional programs characteristic of pluripotency and differentiation, but can respond differently to extrinsic stimuli, such as leukemia inhibitory factor, a cytokine required for maintenance of mESC pluripotency.25,26 Recently, investigators have demonstrated the ability to alternatively produce pluripotent cells from various mammalian somatic cell sources by introduction of exogenous transcription factors capable of fully reprogramming the cell state.27–30 Induced pluripotent stem cells, as they are now called, exhibit many similar characteristics to ESCs with regard to differentiation and self-renewal properties, although subtle differences appear to exist between the genomic and epigenomic signature of the two pluripotent cell types.31,32
Adult stem and progenitor cells have limited self-renewal capacity and possess multi- or uni-potent differentiation potential, such that they are restricted in the lineages to which they can give rise. MSCs and hematopoietic stem cells (HSCs) are commonly studied adult stem cells that can be derived from the bone marrow by sorting based on surface marker expression.33,34 The relative ease of isolation of adult stem and progenitor cells from somatic tissues facilitates the availability of such cell types and permits autologous transplantation. The limited potency of adult stem cells may simplify differentiation and produce a more homogeneous resulting population; however, primary cells can be difficult to obtain and/or impossible to expand for some lineages of interest, such as cardiac, hepatic, and neural cells. Additionally, within the same lineage, the resultant populations from adult and ESCs may differ, as cells derived from ESCs are believed to differentiate into more embryonic-like cells, and therefore possess more immature phenotypes, but potentially enhanced capacity for expansion and regeneration.20,35
Suspension cultures
Monolayer culture of stem cells provides a defined substrate for cellular attachment and uniform application of external stimuli (Fig. 1). However, from a bioprocessing standpoint, monolayer culture is not readily amenable to the production of increased cellular yields necessary for regenerative therapies (upward of 109 cells, depending on the application).36–38 Although scalable adherent culture systems have been developed, these techniques rely on “scaling out” to provide more surface area for growth. In contrast to high density “scale up” techniques, which maintain cells in suspension, scalable monolayer cultures rely on relatively large volumes of media and cytokines, and therefore do not significantly reduce the cost required to maintain the cultures. Adhesion of stem cells to spherical microcarriers increases the surface area available for culture (approximately fivefold) without dramatically altering the exposure to nutrients or waste removal. However, microcarriers rely on stem cell adhesion to exogenous materials, which may alter differentiation.39,40 Culture of stem cells as 3D aggregates is thought to more accurately recapitulate cellular adhesions and signaling exhibited by stem cells found in native tissues.41,42 Microtissues have been created by the spontaneous aggregation of various cell types, resulting in homotypic and heterotypic spheroids, which retain aspects of metabolic, functional, and mechanical properties of living tissues.43–47 Neural stem cells (NSCs) are commonly cultured in suspension format as spheroids, referred to as neurospheres. ESCs are often differentiated in the form of cellular aggregates, termed embryoid bodies (EBs), which parallel many of the phenotypic changes that accompany gastrulation during embryogenesis in vivo, 48 including the epithelial–mesenchymal transition, which is coordinately regulated by the balance of interactions between cells and ECM molecules. 49 Taken together, stem cells can be cultured in a variety of formats and at a range of scales, from adherent monolayers to microcarriers to cellular aggregates in suspension; however, there is an inherent tradeoff between the transport of soluble factors and the scalability of the culture.
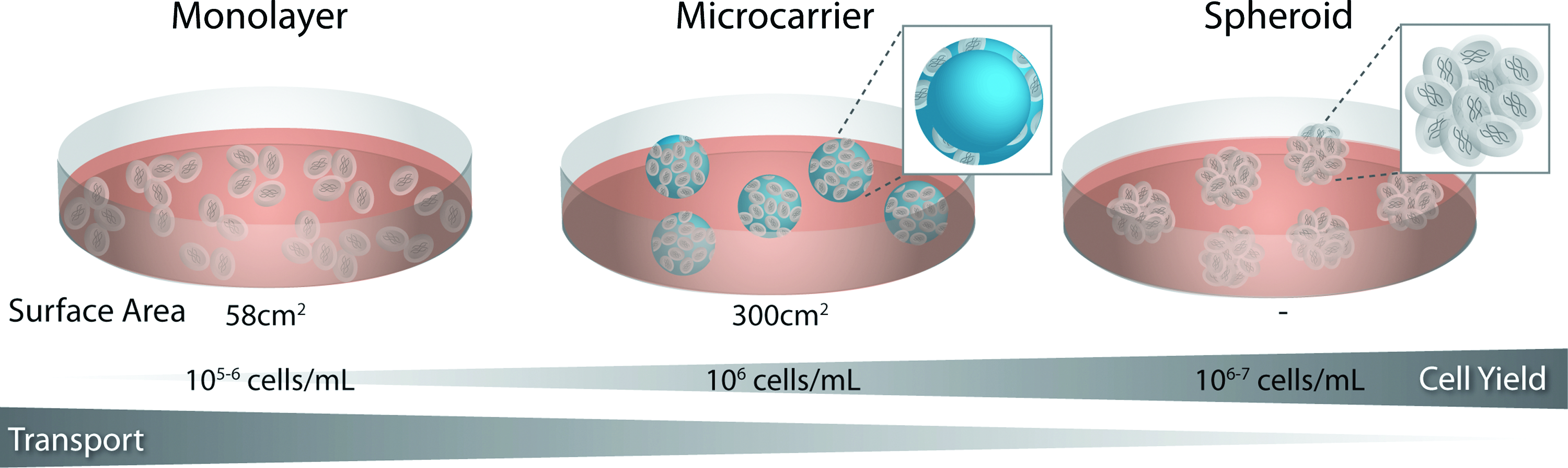
Stem cell differentiation formats. Stem cells can be cultured in monolayer or in suspension, either adherent to spherical microcarriers or as aggregates of cells. Suspension cultures generally increase the density of cells, and thus increase the overall cell yield per volume or media. Although suspension cultures are more scalable, the three-dimensional aggregate structure increases the diffusive distance between the media and cells at the center, which may result in decreased transport and the development of gradients of nutrients and metabolites throughout the spheroid. Color images available online at www.liebertonline.com/teb
Several techniques have been developed to facilitate stem cell spheroid formation and differentiation, including methods that physically separate individual aggregates, which yield homogeneous populations, and batch methods that result in larger yields of EBs but with increased heterogeneity (Fig. 2A). Spheroid formation is especially prevalent in ESC culture because ESCs express E-cadherin during pluripotency and early differentiation, thus enabling the cells to readily, spontaneously aggregate on substrates or in suspension.50,51 Hanging drop cultures are initiated by suspending small volumes of culture media (10–20 μL) containing a defined number of cells (200–1000 cells) from the lid of a Petri dish. Although hanging drops yield uniformly-sized aggregates, the need to physically separate individual drops limits the number of spheroids formed per dish (∼100 per 10-cm plate); thus, hanging drops are not readily amenable to scale-up methods.52–54 It is also traditionally difficult to exchange media in the hanging drop format, though this has recently been accomplished through a modified 384-well hanging drop plate, which contains access ports for manipulation of the media in individual drops. 55 Alternatively, static suspension culture is often accomplished by inoculating cells (103–106 cells) in a 35–100 mm Petri dish, in order to promote formation of spheroids via random aggregation of cells.56–58 Static suspension cultures, however, are often subject to agglomeration of individual spheroids, resulting in large cell masses, which are widely variable in size and shape, and thereby contribute to the overall heterogeneity of differentiation that is commonly observed.

Methods of embryoid body formation and propagation.
Hydrodynamics in large-volume culture systems
Although suspension cultures are amenable to the culture of spheroids statically in bench-scale (<10 mL) volumes, translational approaches are increasingly moving toward larger scale culture systems (>100 mL), in which fluid mixing is introduced to reduce nutrient gradients and promote enhanced gas exchange within the culture volume. 59 Mixing is accomplished either via external agitation of the entire vessel, as in rotating wall vessels, such as the slow turning lateral vessel and high aspect rotating vessel, or by internal agitation of an impeller, as in stirred flasks and pitched-blade bioreactors (Fig. 2B).60,61 The mixing within reactors introduces hydrodynamic conditions, which can generate high shear stresses depending on the rotation speed and vessel design. Quantifying the hydrodynamic environments within bioreactors and bioprocessing systems has been accomplished by combinations of computational fluid dynamic simulations and experimental particle-image velocimetry measurements.62,63 Such analyses aid in defining hydrodynamic properties, including shear stress and velocity profiles as well as frequency characteristics of the mixing system. Overall, results from such analyses indicate that fluid shear varies as a function of spatial position within the vessel, as well as rotational speeds and fluid viscosity.62,64,65 Shear computations for spinner flasks with centrally located constructs indicate that a maximum shear stress of 2.83 dyn/cm2 is imparted on cells within the scaffolds; however, this value is expected to increase near the walls of the bioreactor, and under increased mixing speeds, which may impart high shear stresses on cells in suspension. 66 In contrast to larger volume systems (0.5–2 L), rocking/shaking suspension cultures are externally agitated lab-scale (10 mL) systems, with similar shear ranges (<2.5 dyn/cm2), and are amenable to screening multiple hydrodynamic conditions and additional variables in parallel.65,67,68 The complex shear environments created within 3D fluid mixing systems have motivated the need to better understand the impact of hydrodynamic parameters on cell responses (i.e., aggregation, metabolism, and phenotypes) in mixed culture conditions.
Hydrodynamic Culture of Stem Cells
The impact of hydrodynamics on stem cells manifests via alterations in several key cellular processes, including aggregation (kinetics, size), metabolism (viability, transport, proliferation), and phenotype (differentiation, function) (Table 1). Since all of these processes are inter-related within the context of stem cell biology, the cellular responses cannot easily be attributed to any one single parameter. Therefore, stem cell fate within a mixed fluid system is a result of the synergy between direct and indirect responses imparted by hydrodynamic environmental cues (Table 2).
mESC, murine embryonic stem cell; hESC, human embryonic stem cell; HSC, hematopoietic stem cell; MSC, mesenchymal stem cell; LDH, lactate dehydrogenase; PB MNC, peripheral blood mononuclear cell.
BHK, baby hampster kidney cell; STLV, slow turning lateral vessel; HARV, high aspect rotating vessel; NSC, neural stem cell.
Hydrodynamic effect on aggregation
Mixed culture systems have been used for a long time as a means to facilitate multicellular aggregate formation, whereby a population of single cells self-assembles via cell–cell adhesion receptors (Fig. 3A). As mentioned previously, ESCs aggregate spontaneously in suspension culture via homophilic binding of the cell surface adhesion molecule E-cadherin that is expressed ubiquitously by undifferentiated ESCs. Several other types of stem cells, including NSCs and even MSCs, exhibit a similar propensity to form multicellular aggregates under appropriate conditions via other cell adhesion molecules; however, the molecular variations governing the mechanisms of cellular aggregation may result in altered binding kinetics of different stem cells populations. Therefore, this section discusses the aggregation kinetics within stirred culture systems in the context of EB formation via E-cadherin interactions; however, the principles highlighted can be generally applied to understanding the aggregation of various types of stem cell populations.
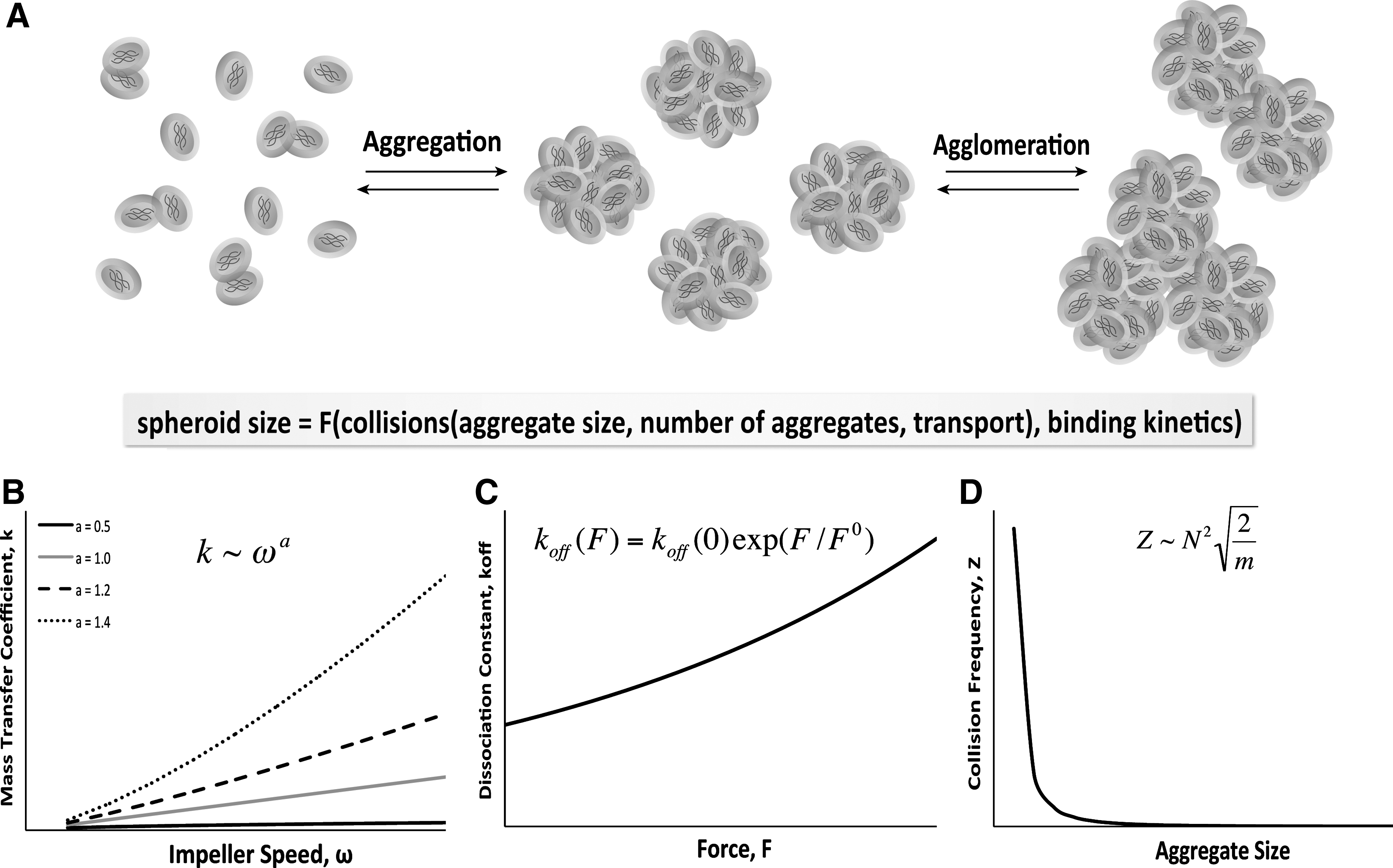
Stem cell aggregate formation.
Mixing within bioreactors has been extensively utilized, modeled, and validated within the field of chemical engineering and in the bioprocessing industry in particular. Therefore, it is well established that convective forces imparted through hydrodynamic mixing increase the mass transfer of the system. One by-product of enhanced mass transfer is more frequent collisions between particles, such as cells, suspended in the fluid. In general, the mass transfer coefficient increases as a power function of the impeller speed (Fig. 3B), and is subject to several parameters specific to the geometrical and physical characteristics of a bioreactor system.69,70 The increased transport in mixed bioreactors is correlated with increased spheroid formation efficiency and cellular incorporation in stirred suspension and rotary orbital suspension compared to static culture.67,71 The vessel size and shape also alter the aggregation dynamics, as indicated by modulation of spheroid formation rate within mixed various culture formats under similar agitation speeds.72,73 In addition to the hydrodynamic parameters, the cellular inoculation density also impacts the frequency of cell collisions in suspension; hence, understanding the changes in transport based on various parameters of the system (mixing, inoculation density) is important for rational design of systems to control stem cell aggregation.
The kinetics of receptor–ligand binding between cells can also be altered as a result of varying convective forces. Previous studies have measured the dissociation constant of homophilic E-cadherin binding to be 0.45/s, which corresponds to a bond duration of approximately 2 s; this bond duration is similar to the transient bonds established between selectins during leukocyte rolling, indicating that the individual E-cadherin bonds can be readily broken. 74 Additionally, the dissociation constant exhibits behavior characteristic of the Bell model for kinetics of receptor ligand binding, resulting in an exponential increase as a function of force (Fig. 3C). Therefore, the shear forces imparted by hydrodynamics can manipulate the kinetics of E-cadherin interactions (or other cell adhesion molecules) in the context of cell aggregation. For example, the altered flow environment within the wavy-walled bioreactor supports increased chondrocyte aggregation compared to standard spinner flasks of comparable dimensions. 59 Similarly, EB formation kinetics are altered at various rotary orbital speeds, with increased rotary speeds delaying, or even inhibiting, EB formation.65,75,76 However, the dissociation kinetics may also change as a function of the aggregation state of the cells, because multivalent E-cadherin interactions likely stabilize intercellular adhesion, and may withstand increased hydrodynamic forces. Taken together, these studies suggest that the increased transport and shear introduced by hydrodynamic culture environments can differentially regulate cellular spheroid formation through alterations in cell–cell collisions and adhesion binding kinetics at the cellular and molecular level, respectively.
The cell–cell adhesions established during EB formation are critical to differentiation and morphogenesis; however, a fine balance exists between the aggregation necessary for initial spheroid formation and agglomeration of individual spheroids. Excessive agglomeration typically results in large, necrotic regions located centrally within multicellular aggregates, as is commonly observed in static suspension cultures of EBs. Encapsulation methods have been used successfully as a means to culture ESCs and MSCs in mixed hydrodynamic bioreactors without agglomeration.77–81 Although encapsulation of ESCs within nonadherent hydrogels prevents agglomeration initially, as EBs grow in size they can protrude beyond the edges of agarose or alginate beads and begin to agglomerate.82,83 As spheroids merge due to agglomeration, the mass of individual spheroids increases, and the total concentration of spheroids in suspension decreases, leading to an overall decrease in the collision frequency between multicellular aggregates (Fig. 3D). The total surface area available for interaction between spheroids in culture also decreases, which may alter binding kinetics. The kinetics of spheroid agglomeration, however, may also be complicated by remodeling of cell adhesions as morphological events proceed, including changes in cadherin expression profiles, formation of adherens and tight junctions between cells, and heterophilic interactions with components of the ECM. Since collision frequency and binding kinetics are both altered by hydrodynamic parameters, spheroid size in mixed suspension cultures can be significantly impacted by agglomeration. Stirred suspension cultures maintained over a range of hydrodynamic conditions often exhibit decreased agglomeration compared to static suspension, resulting more homogeneous populations of mESCs and hESCs.65,67,75,84 Alteration of the hydrodynamic environment by employing different impeller types or changing media viscosity also alters the homogeneity and size of spheroids.85–87 in general, an inverse relationship exists between agitation speed and spheroid size, as evidenced by work in stem and progenitor cells from different species cultured in various types of stirred and mixed bioreactors.65,85,88–90
The aforementioned results indicate that various parameters of the hydrodynamic environment, including the configuration of the culture vessels and mixing conditions, may alter the transport and binding characteristics of stem cells, ultimately impacting the terminal size of spheroids. The control of spheroid size within hydrodynamic environments is a fine balance between promoting initial multicellular aggregate formation while limiting secondary spheroid agglomeration.
Hydrodynamic effects on metabolism
Physiochemical properties of the culture environment that promote the highly proliferative state of stem cells have been established based on known cellular requirements and parameters from in vivo conditions (Fig. 4). For example, the high metabolic demand required for proliferation of stem cells usually necessitates culture in media containing high concentrations of glucose (25 mM). When oxygen is readily available, glucose is metabolized by conversion of pyruvate into ATP through the tricarboxylic acid cycle, whereas in low oxygen conditions, pyruvate is converted to lactate by the enzyme lactate dehydrogenase (LDH) during anaerobic glycolysis. Oxygen uptake rates for mammalian cells range from approximately 0.05–0.5 μmol/106 cells/h, 91 with hematopoietic cells from the bone marrow exhibiting some of the lowest rates (as low as 0.005 μmol/106 cells/h). 92 Recent studies indicate a link between the oxidative metabolic state and self-renewal in adult and ESC populations93,94; highly proliferative cells exhibit increased glycolysis and decreased oxygen consumption, similar to the Warburg effect in cancer cells. 95 The mixing within large volume bioreactors and bioprocessing systems aids in the transport of nutrients and metabolites, and thereby enables sampling, monitoring, and control of the environment. HSCs are a useful model for understanding the impact of convective transport on expansion, because the culture of HSCs as single cells in suspension avoids the diffusive transport limitations in spheroids (Fig. 4A). Computational modeling of nutrient and oxygen transport supports experimentally observed increases in hematopoietic cell expansion within mixed cultures compared to static suspensions.96–100 Additionally, the number of cells inoculated within bioreactors impacts hematopoietic cell metabolism, with a minimum lower limit (∼2×105 cells/mL) required to facilitate expansion. 97
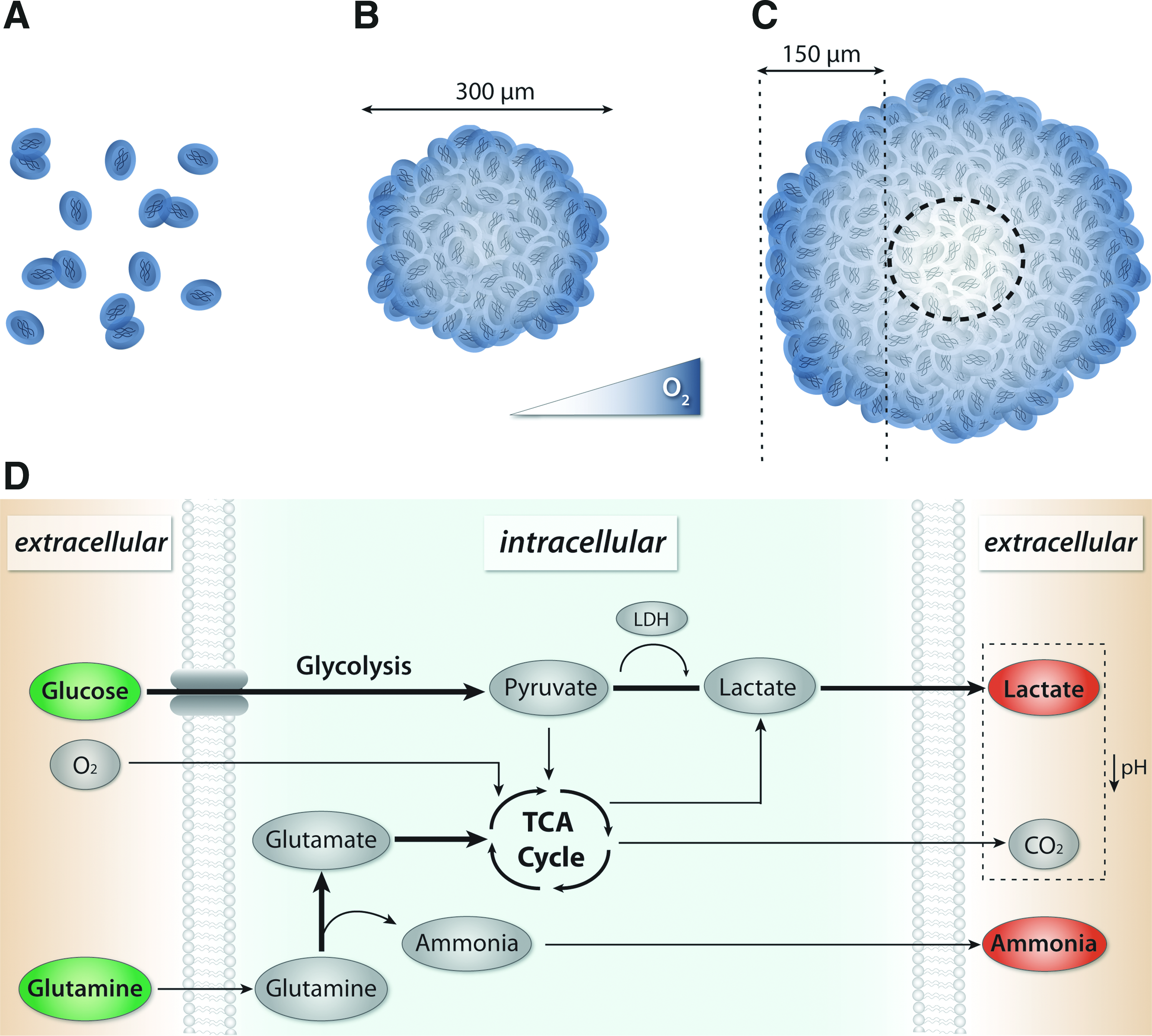
Macro- and microscale changes in transport and stem cell metabolism.
In contrast to single-cell suspensions, such as for HSCs, cells maintained as aggregates in culture confound metabolic analyses because gradients develop within the 3D structures (Fig. 4B, C). Ultimately the heterogeneous distribution of oxygen and nutrients within spheroids impact cell expansion and the doubling time of stem and progenitor cells. 101 For example, inoculation of spinner flasks with pre-formed spheroids of NSCs resulted in faster cell expansion, whereas a single cell NSC suspension produced a larger overall expansion, indicating that metabolic differences may be linked to spheroid size and formation kinetics. 57 In purely diffusive culture conditions, necrosis occurs in spheroids larger than approximately 300 μm in diameter, 88 presumably due to O2 transport limitations, which indicates that the maximum diffusive distance is on the order of approximately 100–150 μm; however, in bioreactors, metabolic gradients are influenced by the transport properties (diffusive and convective) of mixing conditions. Hollow fiber capillary membrane bioreactors are a promising technology for decreasing diffusive distances approximately 10-fold, by providing media perfusion via many parallel sources distributed evenly throughout the culture. 102 Additionally, inherent diffusional barriers of the spheroids may arise as a function of differentiation state; after several days in culture, EBs develop an exterior epithelial cell layer that limits diffusion of soluble factors, indicating that transport may be altered as a function of differentiation state.103,104
The impact of shear and transport within cell aggregates can be indirectly monitored through changes in the metabolic profile, specifically through extracellular LDH levels, indicative of cell lysis. Mixing conditions may be detrimental during early aggregation, as indicated by elevated LDH during early culture periods compared to static cultures; however, static cultures exhibit comparatively increased LDH at later stages of differentiation, likely due to cell death. 72 mESCs in rotary orbital suspension demonstrate increased viability compared to static culture, with a decreased instance of necrotic core formation, likely due to a combination of decreased EB size (limited agglomeration) and increased convective transport of nutrients to the center of EBs. 67 Spheroid cultures also exhibit a cumulative increase in released LDH (50–150 U/mL) 72 compared to microcarrier cultures (<5 U/mL), 105 indicating cell death as a result of differences in transport and shear forces between the different culture formats.
Monitoring of the physiochemical environment indicates that comparable ranges of nutrients and metabolites are maintained within suspension cultures of spheroids and in standard monolayer cultures. 99 Additionally, comparable levels of glucose (1.94–1.23 g/L; 125 mL vs. 500 mL bioreactors after 20 days in culture), lactate (0.81–1.30 g/L), glutamine (1.21–1.13 mM), and ammonia (1.00–1.07 mM) are maintained when mixed NSC cultures are scaled from 125 to 500 mL bioreactors. 89 Studies suggest, however, that the hydrodynamic environment within several mixed culture formats, including rotary, roller ball, and spinner flasks, supports increased hESC stem cell proliferation compared to static suspension cultures.75,76,86 Additionally, hESCs adherent to microcarriers in mixed cultures with mouse embryonic fibroblast conditioned media exhibited modulation of the metabolic state, including high glucose and glutamine consumption, increased ammonia and lactate production, and acidification of the media (Fig. 4D); however, the metabolic state may also be altered depending on specific media formulations. 106 Glutamine supplementation may be a limiting factor for adipose stem cell expansion within batch cultures maintained over 150 h, because the cells also exhibit decreased proliferation in the absence of glutamine 107 ; removal of the amino acid from culture media results in cells entering a quiescent-like state with decreased metabolic requirements, including reduced glucose consumption. Additionally, the accumulation of metabolites, specifically lactate and ammonia, which can create unfavorable culture conditions, such as decreased pH, may be a factor leading to decreased cell viability, and therefore limiting total cell expansion. 76 Correlation of the physiochemical signature of the culture environment with phenotypic indicators may permit indirect real-time analysis of the differentiation state of stem cells by monitoring secreted metabolic factors. 98 In particular, due to the heterogeneous proliferative and differentiation states of stem cells, increased control of metabolites may be important for the development of more efficient expansion and directed differentiation protocols. For example, monitoring of bioreactor conditions and constant media exchange via perfusion are important modifications, which may enable the automation and standardization of culture conditions.102,108–110 Mixed culture conditions that support stem cell expansion and differentiation, therefore, require a complex balance between providing sufficient concentrations of nutrients and limiting accumulation of toxic metabolic by-products, while also taking into account the potential effects of fluid mixing on cell lysis.
Hydrodynamic effects on phenotype
As noted previously, stem cell phenotype is exquisitely sensitive to a variety of environmental cues, many of which can be modulated within mixed culture environments. Due to the heterogeneous nature of differentiation within stem cell spheroids and the complex hydrodynamic environment, it can be difficult to ascertain which parameters directly and indirectly impact cell phenotype.
Mixed bioreactors and bioprocessing systems have been utilized as high yield configurations for expansion and differentiation of stem cell populations. For example, in the presence of appropriate environmental conditions (leukemia inhibitory factor supplementation or conditioned media), mESCs and hESCs expanded as aggregates or on microcarriers within mixed bioreactors can be maintained in a pluripotent, undifferentiated state.76,105,111–115 Several groups have also demonstrated that stem cell differentiation potential is not altered within bioreactor systems of varying volumetric scale, agitated either externally or internally; MSCs, 116 NSCs,89,117,118 and ESCs67,72,75,119 retain the capacity to differentiate into their respective cell lineages, although some studies indicate lower efficiency of differentiation. 120 Compared to static suspension cultures, the endogenous propensity of some ESC lines to differentiate toward particular lineages does not seem to be altered over a range of hydrodynamic conditions. For example, Fok and Zandstra observed no statistical difference in cardiac or hematopoietic differentiation between EBs cultured in spinner flasks (60 and 100 rpm) compared to EBs differentiated in static cultures 119 ; similarly, there was no difference in cardiomyocyte differentiation between static mEBs and those cultured within slow turning lateral vessels. 71
However, studies indicate changes in global gene expression profiles within differentiating MSC and ESC spheroids maintained in hydrodynamic culture conditions compared to static conditions,65,121,122 suggesting discrepancies in the tendency of dynamically cultured stem cells to differentiate toward particular lineages. The propensity for MSCs to differentiate along adipogenic lineages is increased when cultured in mixed vessels.121,123 Additionally, MSCs exhibit enhanced osteogenic differentiation when MSCs are cultured dynamically as aggregates 121 ; however, evidence also supports decreased osteogenic differentiation of MSCs on microcarriers. 123 The difference in MSC phenotype may, however, be explained by culture on microcarriers compared to as aggregates; the properties of exogenous substrates may alter differentiation, as matrix elasticity can modulate stem cell phenotype, with stiffer matrices promoting osteogenic differentiation.39,40 Within aggregates of ESCs, the bioreactor type impacts the hematopoietic differentiation profile, with an increased Sca-1+ population derived from cultures in rotating wall vessels, compared to increases in C-kit+ cells maintained in spinner flasks. 124 Additionally, others have illustrated enhanced cardiomyocyte differentiation in spinner flasks compared to static conditions, 125 as well as within rotary orbital suspension culture. 68 The agitation speeds in various systems also modulate ESC phenotype, 65 as EBs maintained at a range of rotary orbital speeds exhibit temporal modulation of gene expression indicative of changes in differentiation among the three different germ lineages.
As noted previously, mixing speeds can modulate spheroid aggregation kinetics and spheroid size, with slower speeds leading to larger spheroids, which form more quickly than at slower speeds. The noted differences in cardiomyocyte differentiation, therefore, may result from changes in the metabolic environment, as a result of decreased transport within larger aggregates compared to smaller spheroids. 126 Several groups have linked hypoxic conditioning to increased hematopoietic and cardiogenic differentiation.101,108,125,127 Similarly, metabolites in the environment, including lactate, alter MSC gene expression 128 and drive angiogenic response in macrophages. 129 Changes in bioreactor configurations that alter the metabolic environment may be more or less conducive to certain cell phenotypes, thus promoting or inhibiting differentiation to specific cell phenotypes. Additionally, cell organization is also central to morphogenesis and differentiation through the expression of lineage-specific cell adhesion molecules, such as E-cadherin. Binding and remodeling of adhesion receptors can alter differentiation through the activation of downstream signaling pathways such as the Wnt/β-catenin pathway,50,51 thus highlighting the importance of the temporal aspect of cell aggregation and the implications for altering downstream signaling kinetics. Altered phenotype based on aggregation kinetics and spheroid size has been established through research on the enrichment of mesoderm and cardiogenic phenotypes when aggregates are formed using hanging drop culture.54,56,68,130 As is discussed throughout this article, cell aggregation, metabolism, and phenotype are all interconnected within the context of stem cell biology, which underscores the importance of developing methods for decoupling these variables, in order to understand the impact of environmental conditions on stem cell spheroids.
Future Opportunities
The complexities of hydrodynamic culture environments drive the need to systematically examine individual parameters since elucidating direct, indirect, and synergistic effects will enable more rational design of bioreactors and bioprocessing systems. For example, simultaneous alterations in spheroid size and differentiation of ESCs within mixed culture systems highlight the importance of separating independent variables to understand the direct influence of hydrodynamic forces. Several groups have recently developed methods for controlling ESC colony and stem cell spheroid sizes131–135 as well as aggregate shapes (i.e., rods, tori, honeycombs, rectangles, annuli, and sinusoidal bands),136–139 using microprinting and microwell technologies to physically separate cells in high-throughput formats. These advances have enabled the analysis of stem cell differentiation as a function of aggregate shape 138 and size,132,140 and have noted increased cardiogenic differentiation within EBs of approximately 400 μm diameter.125,134 Changes in cardiogenic differentiation due to spheroid size have been correlated to non-canonical signaling of the Wnt pathway through altered expression of Wnt5a and Wnt11 134 ; however, the specific mechanism for this regulation remains largely unknown. Additionally, the technologies for producing size-controlled EBs may also enable analysis of the influence of hydrodynamics, independent of spheroid size. These studies may, however, only be amenable for a certain range of hydrodynamic conditions, due to the potential for individually formed spheroids to agglomerate in suspension. Additionally, formation of spheroids before hydrodynamic culture alters both spheroid size and formation kinetics. Ultimately, development of culture methods to independently control spheroid size and formation kinetics will be important for controlling stem cell expansion and differentiation.
Changes in differentiation due to spheroid size may also be a result of altered transport profiles within both diffusive and convective environments. Within mixed bioreactors, modulation of the hydrodynamic environment changes both transport and fluid shear; development of methods to decouple these variables may elucidate governing parameters within these cultures. For example, modeling of convective transport within various mixed culture systems may elucidate changes in metabolic parameters as a result of spheroid size. Additionally, methods for eliminating gradients of nutrients within the interior of spheroids will likely result in more homogeneous directed differentiation of stem cell aggregates. Microspheres have been incorporated within stem cell aggregates, enabling delivery of morphogens more homogeneously throughout the multicellular microenvironment.104,141–145 The combination of the hydrodynamic culture environment with microsphere-mediated delivery may enable analysis of the impact of fluid shear on cell spheroids, independent of small molecule and growth factor transport limitations. Additionally, combinatorial methods for simultaneously controlling morphogen delivery and the hydrodynamic environment could lead to synergistic methods to more efficiently and effectively direct stem cell differentiation.
The modulation of stem cell aggregation, metabolism, and phenotype by mixed cultures has implications for bioprocessing and bioreactor design for future applications in stem cell therapies. Bench-scale systems (100–1 mL) such as rotary orbital culture may be useful to examine the effects of hydrodynamic conditions on stem cell formation, morphology, and differentiation because, using smaller volume cultures, multiple parameters can be systematically probed in parallel in a rapid manner. Defining the governing properties within hydrodynamic culture systems that regulate differentiation will aid in the development of efficient bioprocessing protocols and bioreactor design. This goal necessitates detailed analysis of the hydrodynamic parameters within various bioreactor formats, under different mixing configurations and speeds. Several formats currently exploit the inherent scalability of suspension cultures, to employ large volume mixed systems (102–3 mL), enabling the increased production of differentiating cells required for large-scale drug screening or the potential therapeutic application of stem-cell-derived products. Large-scale stirred culture systems are advantageous because they are amenable to numerous parallel modifications, which provide media perfusion and dialysis, enable automated maintenance of appropriate metabolic conditions, and decrease physiochemical fluctuations, which occur during manual feeding by bulk media exchange.89,90,108,117,125,146,147 Ultimately, defining the governing hydrodynamic parameters for altering aggregation, metabolism, and phenotype, as well as synergies between the interdependent variables will enable engineering of bioreactor and bioprocessing systems for controlled, scalable, and efficient expansion and differentiation of stem cells.
Footnotes
Acknowledgments
The authors are supported by funding from the NIH (EB010061). M.A.K. is currently supported by a National Science Foundation Graduate Research Fellowship, and C.Y.S. was supported by an American Heart Association Pre-Doctoral Fellowship.
Disclosure Statement
No competing financial interests exist.