
Abstract
The repair and management of full-thickness skin defects resulting from burns and chronic wounds remain a significant unmet clinical challenge. For those skin defects exceeding 50%–60% of total body surface area, it is impractical to treat with autologous skin transplants because of the shortage of donor sites. The possibility of using tissue-engineered skin grafts for full-thickness wound repair is a promising approach. The primary goal of tissue-engineered skin grafts is to restore lost barrier function, but regeneration of appendages, such as hair follicles, has to be yet achieved. The successful regeneration of hair follicles in immunodeficient mice suggests that creating human hair follicles in tissue-engineered skin grafts is feasible. However, many limitations still need to be explored, particularly enriching isolated cells with trichogenic capacity, maintaining this ability during processing, and providing the cells with proper environmental cues. Current advances in hair follicle regeneration, in vitro and in vivo, are concisely summarized in this report, and key requirements to bioengineer a hair follicle are proposed, with emphasis on a three-dimensional approach.
Introduction
The lack of hair in grafted sites has both psychosocial and physiological impacts on the patient. 10 Hair follicles cannot regenerate on their own after damage. 8 In this regard, new and healthy hair follicles or trichogenic cells need to be introduced to the affected area for regeneration of hairs. Several models have been established to address the reconstitution of hair follicles; however, most are based on the intact, healthy skin of immunodeficient mice,7,11 overlooking the complex effects of the wound microenvironment. In contrast to the extensive efforts in developing tissue-engineered skin grafts from hair follicle-derived stem cells, 12 attempts to regenerate hair follicles in skin grafts have been limited. This is partially due to the lack of an effective approach to incorporate follicular cells into skin grafts. This review briefly discusses the fundamental components of hair follicles, current advances in follicular regeneration, and key requirements to engineer follicles.
Anatomy and Biology of Hair Follicles
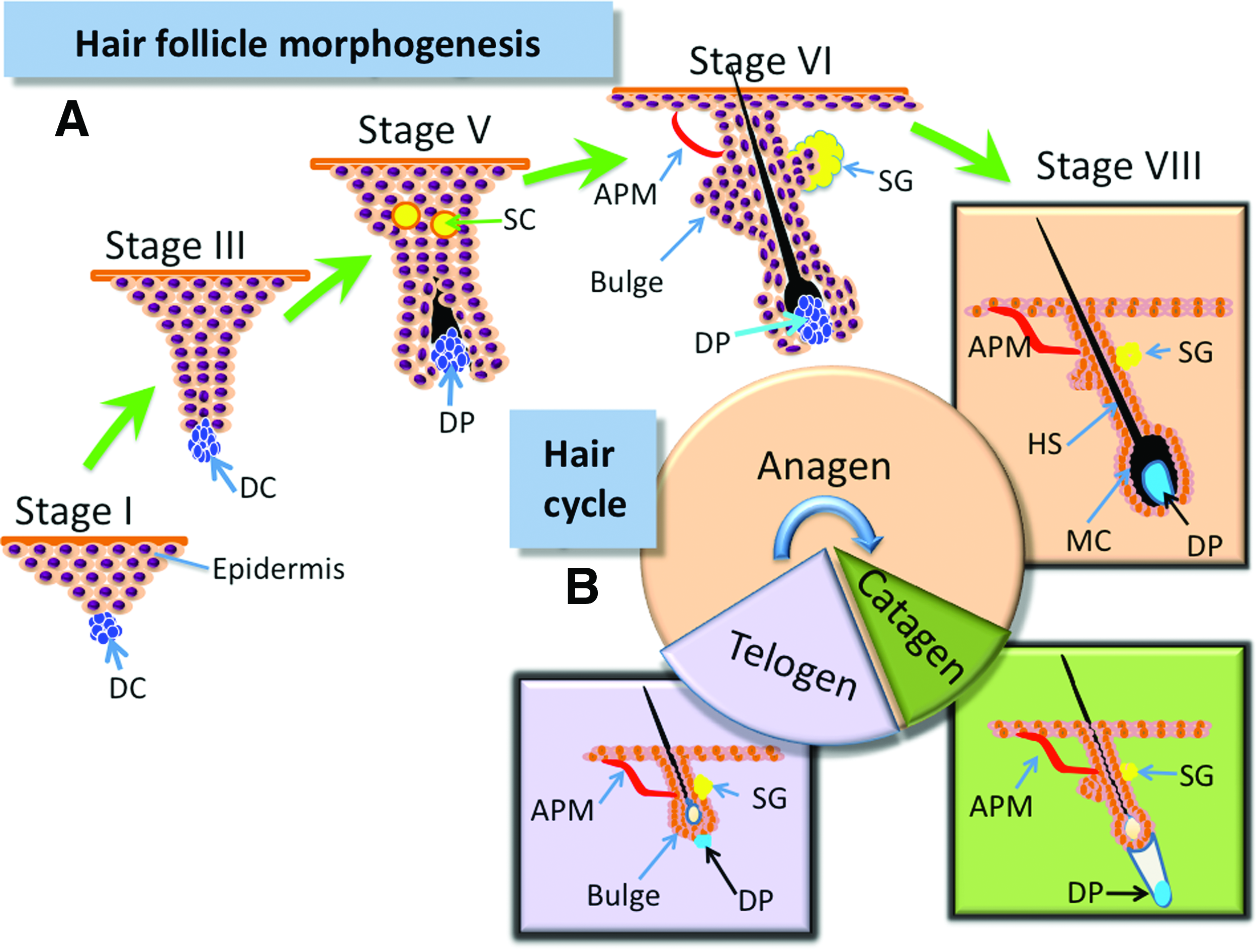
The mature hair consists of the hair shaft and the oval hair follicle with a multicylindrical stem. A human has around 5 million hair follicles with two types of hairs—terminal hair (long, thick pigmented) and vellus hair (thin, unpigmented).2,13 Hair follicles are complex, highly regenerative, ectodermal–mesodermal structures enriched with stem cells. They have lifelong cycles of growth following three distinct stages: anagen (rapid growth), catagen (regression), and telogen (resting period) (Fig. 1B). Intrinsic control of each stage involves various factors (e.g., cytokines, hormones, neurotransmitters, and their cognate receptors),14,15 even though the coordination of these factors remains to be elucidated.
In the hair follicle, dermal papilla (DP), also called follicular papilla, an onion-like structure surrounded by hair bulb matrix (Fig. 2), is considered the commander of the hair follicle. It determines the hair thickness, length, and life cycle by adjusting the volume of DP cell (DPC) aggregate and secretory activity.10,16 The hair bulb not only produces the shaft, but also provides melanin granules to trichocytes for pigmentation.
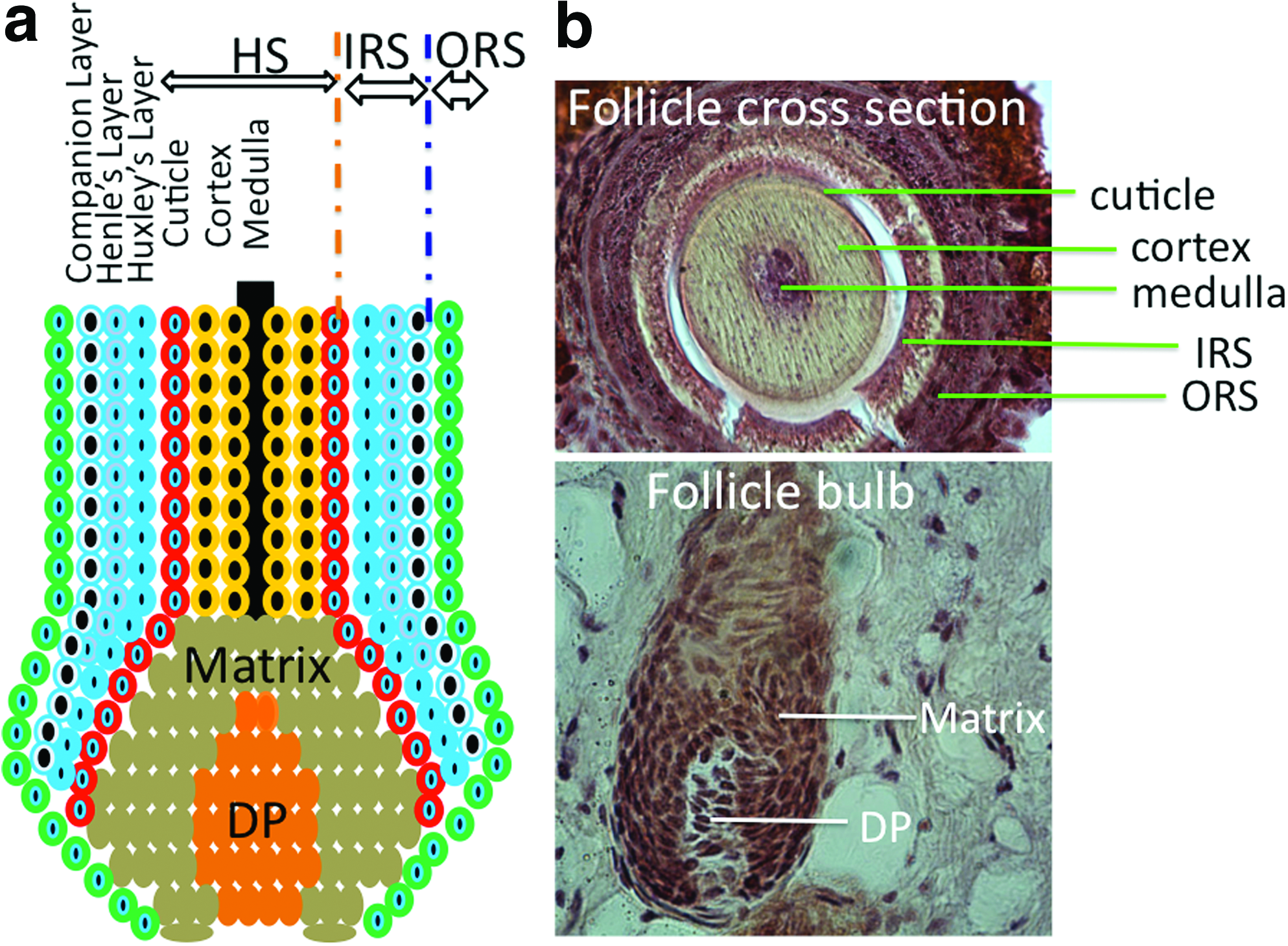
Hair matrix keratinocytes, located within the hair bulb, are the point of origin for trichocytes and cells of the inner root sheath (IRS) (Fig. 2). These cells have a high proliferative capacity, important for the growth and regeneration of the hair follicle and epidermis. The pluripotent epithelial stem cells located within the bulge can differentiate into outer root sheath (ORS), hair matrix, and hair shaft 17 and can generate new hair by forming a secondary hair germ and help the wound healing process by reconstituting keratinocytes in response to damages to the bulge area. 10
The dermal sheath (DS) surrounding the hair follicle contains progenitor cells that can differentiate into DP upon the removal of papilla. 18 Beneath the DS, lies the ORS, and around the IRS is the basal layer of interfollicular epidermis. 14 The IRS contains four layers: companion layer (CL), Henle's layer, Huxley's layer, and the IRS cuticle (Fig. 2). 19 The interaction between mesenchymal (mainly DPCs) and epithelial (mainly follicular keratinocytes) portions of the hair follicle plays an essential role in normal hair growth. 14 Other elements of the hair follicle, such as neurons and blood vessels, are derived from mast cell precursors. 20 Neuronal stem cells are also found in hair follicles. 21 As a center for several populations of stem cells, hair follicles have gained increasing attention for use as a cell source in regenerative medicine.
Functions of Hair Follicles
Although the precise role of hair cycling is unclear, it is generally accepted that this process removes debris and parasites from skin surface and encapsulates harmful chemicals within trichocytes. 15 Hair cycling also serves as a self-protecting system, by removing rapidly proliferating keratinocytes in the catagen phase and preventing oxidative stress and malignant degeneration. 10 It is speculated that the hair follicle may also have paracrine and endocrine functions on various cells and structures within the skin.10,22,23 Many factors secreted by follicles are known to be involved in wound healing, and mouse models have shown a relationship between the presence of follicles and the rate of wound healing, reepithelialization, and tissue expansion. 23
Trichogenesis and Trichogenic Cells
The term trichogenesis refers to the cells' ability to form a new hair follicle. It is easy to postulate that cells at different stages of folliculoneogenesis have different trichogenic potentials and the most trichogenic cells should be those in the earliest stages of hair follicle development. During embryogenesis, mesenchymal stem cells aggregate into dermal condensations beneath the epidermis. With the participation of various factors, these aggregates induce the proliferation and differentiation of epidermal cells into follicular cells (Fig. 1A). 7 Zheng et al. 24 found that most trichogenic dermal cells were in a brief window between stages 1 and 3 of follicle development (Fig. 1A) and they showed that hypodermal injection of trichogenic mouse cells from stages 1 to 3 into mouse skin led to rapid formation of hair follicles. Although cells in early folliculoneogenesis stages have the most trichogenic capacity, adult hair follicles still retain their ability to regenerate. 25
Regeneration of hair follicles during the normal hair cycle is mainly controlled by stem cells located in the epithelial bulge. Recent studies have shown that follicular stem cells can be isolated and used to regenerate hair follicles upon injection into immunodeficient mice when combined with follicular dermal cells. 26
Follicular dermal cells play an essential role in hair morphogenesis and cycling. They can be divided into two distinct groups: DPCs and DS cells (DSCs), based on their specific markers, morphology, and function. 27 Both cells are thought to be from the same mesenchymal origin, a dermal condensation, 28 although some consider the DPCs to be of neural crest origin because of their expression of neural markers.29,30 DPCs and DSCs share several properties, but also have notable distinctions. For example, DSCs have a low alkaline phosphatase (ALP) activity and a high expression of alpha-smooth muscle actin (α-SMA) both in vivo and in vitro. In contrast, DPCs have a high ALP activity and an undetectable expression of α-SMA in vivo. Only upon in vitro culture, they show strong α-SMA expression (Fig. 3). 27 DSCs are thought to play an important role in maintaining the DP, acting as a reservoir for DPCs.31,32 DSCs are able to differentiate into DPCs upon DPC loss, 30 and vice versa, 13 suggesting functional similarity and comparable hair formation potential between DPCs and DSCs. 33 Actually, both DPCs and DSCs can substitute for dermal condensations to produce hair follicles as shown in mouse ears and footpads. 30 However, without epidermal cells, neither DPCs nor DSCs can form hair follicles, 34 demonstrating the necessity of mesenchymal–epidermal interactions.
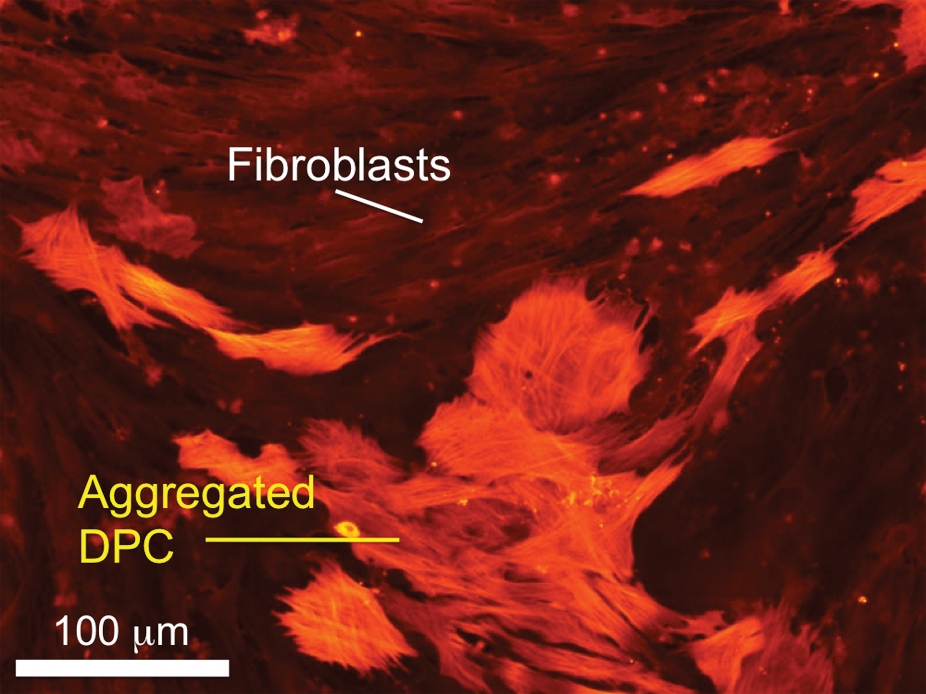
Coculture of rat vibrissa DPCs with DFBs. DPCs expressed alpha-smooth muscle actin upon in vitro culture tended to aggregate, whereas DFBs did not. Scale bar=100 μm. DPCs, DP cells; DFBs, dermal fibroblasts. Color images available online at www.liebertonline.com/teb
Isolation and Culture of Trichogenic Dermal Cells
Both DSCs and DPCs can be expanded in vitro and used to generate hair follicles in vivo. The feasibility of this approach has been demonstrated by Reynolds and Jahoda using adult rat skin.19,34 To obtain DPCs or DSCs, various protocols have been explored, including explant outgrowth and enzymatic dissociation. In both approaches, microdissection is required to extract the hair follicles.7,33 In contrast to the low yield from explant outgrowth, 35 which is thought to be mainly due to poor explant adhesion, enzymatic dissociation using type IV collagenase, 36 dispase, 37 or a combination of type I collagenase and dispase 38 has proven to be more effective in obtaining a large quantity of viable DPCs within a short time frame. However, enzymatic damage to isolated cells and cell surface proteins may still be a problem. Li et al. 39 have tried to facilitate the attachment and outgrowth of DPCs from follicular explants by a brief digestion of the hair follicles with collagenase I. This approach still requires approximately 1 week for the cells to migrate out. Further improvement of cell isolation efficiency is still needed.
Approaches to hair follicle formation with isolated cells are largely similar, but work is still needed to find ways of maintaining trichogenic potential. It has been found that the capacity of DPCs to aggregate closely correlates with their ability to form hair follicles. 40 Freshly isolated DPCs retain a high tendency to aggregate, but this characteristic gradually disappears with extended culture. Horne and coworkers 41 observed that DPCs cultured beyond 6 passages could no longer form cell aggregates or induce hair follicles upon implantation. Supplementation of culture media with 10 ng/mL fibroblast growth factor 2 (FGF-2) could eliminate this restriction by rescuing the lost inductive capacity of DPCs, even at passages as late as 26. 42 This has been demonstrated with both rat and human cells.41,42 Coculture with keratinocytes or culture in keratinocyte-conditioned media also helps to retain the follicle-inducing capacity of passaged DPCs up to passage 90. 43 These findings allow the generation of large numbers of inductive dermal cells from a small number of isolated hair follicles.
In Vivo Induction of Hair Follicles
Formation of new hair follicles does not occur in the adult; however, follicle formation can be experimentally induced using appropriate hair follicle structures or cells.25,44 In this endeavor, early efforts were made by transplanting either entire intact follicles or partial follicles. For instance, Cohen has shown the formation of new whisker by implanting isolated rat and guinea pig vibrissa trichogenic papilla to the rat ear. 45 Following a similar approach but with only different portions of whisker follicles, a series of studies were performed by Oliver in an attempt to regenerate partial or complete hair follicles. It was found that the regeneration of DP was a prerequisite step to hair follicle formation, and root sheath cells surrounding hair follicles could regenerate the DP.18,46 It has been reported that intact DP, with epidermis formed from the keratinocytes of a rat footpad, can reconstruct the hair follicle and also sebaceous gland-like structures upon transplantation into the dorsal skin of a rat. 47 Despite the success of hair follicle transplantation, its potential application is strictly constrained by the availability of intact follicles.
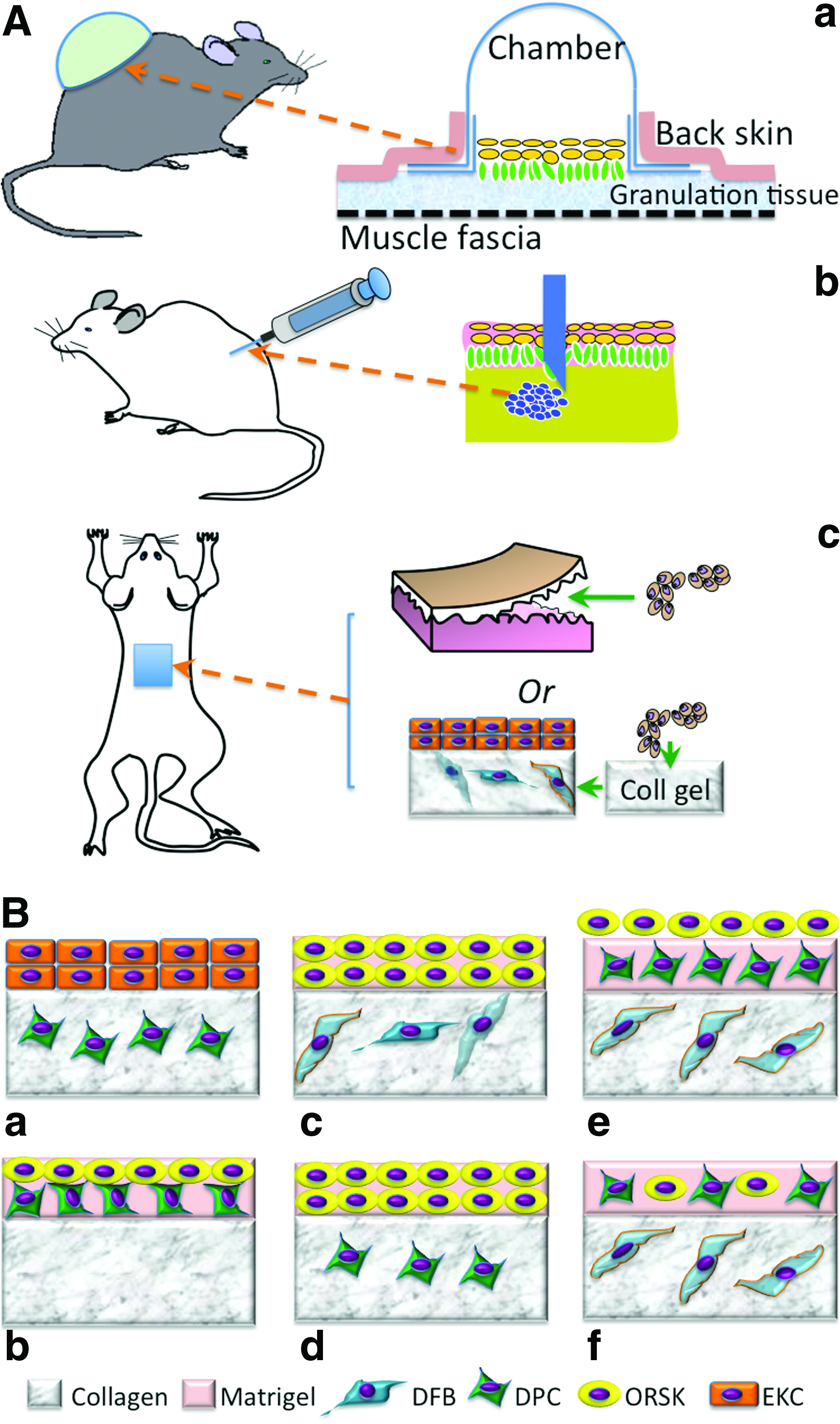
In recognition of the imperative contribution of DP to in vivo neofollicle formation, it is reasonable to assume that the cells isolated from DP alone may be sufficient to induce new hairs. To better evaluate the regeneration of hair follicles by dissociated single cells or aggregates, several suitable experimental models have been established. Three major models, that is, the silicon chamber model, 48 the patch implantation model, 24 and the subcutaneous cell injection model, 49 have been developed for in vivo use in immunodeficient mice (Fig. 4A). In the first model, a bell-shaped silicon chamber is used to cover the murine full-thickness wound and thus protect cells in the wound bed. Using this model, it has been shown that a mixture of neonatal mouse epidermal and dermal cells can lead to hair formation after 3 weeks.48,50 With the same model, Kishimoto and Ehama assessed the trichogenic efficiency of dissociated DPCs using an elegant green fluorescent protein tag to determine the involvement of versican-positive DPCs in folliculogenesis. 51 The result clearly confirmed that only versican-positive DP-derived cells could result in new hair follicle formation but not negative ones. Compared with the chamber model, the subcutaneous injection model [Fig. 4A(b)] is more convenient and rapid for a wide application. Using this model, Jahoda et al. 41 have also shown that cultured papilla cells can lead to hair growth after injection into the follicles of a nude mouse ear. However, this subcutaneous injection of DPCs mainly restores preexisting injured hair follicles rather than completely regenerating a new one probably because of the lack of mesenchymal–epidermal interaction. To this end, an effort was made to subcutaneously inject a “hair bud,” composed of epidermal cell aggregates and DPCs, and indeed new hair formation was observed, 25 suggesting the need of epidermal component in DPC-induced folliculogenesis. Interestingly, in this study the hair follicles grew outward from the aggregates, with the hair shaft projecting inward to the apoptotic cavity in the center of the epidermal aggregate. 25 The subcutaneous implantation of a mixture of DS and ORS from human hair follicles within a customized cabin (1 cm in diameter and 0.5 cm in height) or as a “cyst” induced new follicle-like structures, but only the “cyst” showed hair shaft formation, 28 indicating the need of cell aggregation. Potential challenges with this model are the difficulties of controlling the injection location and cell confinement and guiding hair to grow outward. In this regard, patch implantation can better overcome these issues. Jahoda and coworker34,52 demonstrated that both pelage (i.e., small hairs of animals) and vibrissa DPCs retained their trichogenic capacity by introducing either pelage or vibrissa DPCs in between the enzymatically separated epidermis and dermis of embryonic rat foot pads and then transplanting the combination to the rat dorsum. Similarly, the subcutaneous implantation of mouse foot pad tissues with DPC aggregates (1×104 cells/aggregate) inserted in the epidermal–dermal pockets formed follicular structures after 2 weeks in nude mice. 42 So far all these in vivo models have proved the competency of DPCs in inducing hair follicle formation; however, this capacity requires the synergy of epidermal cells from either the epidermis, ORS keratinocytes (ORSKs), or bulge keratinocyte stem cells. Notably, the obtained in vivo folliculogenesis is mainly evidenced in mice or rats, which do not completely replicate the human environment. Further studies are needed to translate the results in mice to those in humans.
In Vitro Generation of Hair Follicles
Unpredictable in vivo hair formation has inspired efforts to find a more controllable and reliable in vitro system. Several potential advantages can be identified from the in vitro system as summarized in Table 1 along with some foreseeable challenges.
The development of various three-dimensional (3D) culture methods has dramatically enhanced the possibility of in vitro approaches toward follicle formation. Clearly, effective in vitro culture needs to maximally mimic the elements of an in vivo environment, for example, creating a 3D culture where follicular cells can form concentric aggregates, maintaining the interaction between ORSKs and DPCs, incorporating basement membrane proteins, and maintaining DPCs at a low level of proliferation and apoptosis. The replication of DPC–epithelial cell interaction is considered a crucial step leading to follicle formation. To do this a most popular organotypic culture model has been developed and widely used, which combines different cell populations such as DPCs, keratinocytes, and follicle ORS cells in either collagen gel or matrigel, or a sandwiched mixture of both.28,53 The model can be as simple as just encapsulation of the cells in the hydrogel, for example, coculture of keratinocytes and ORS cells from human hair follicles in collagen gel or matrigel produced epidermoid cyst-like spheroids and spike-like structures. 54 The culture of epithelial cells of the inferior portion of human hair follicles on DSC-populated gel showed the formation of hair follicle-like concentric structures. 28 Compared with single gel cultures, the sandwiched collagen and matrigel system better mimic the spatial organization of skin extracellular matrix (ECM). Several representative combinations as shown in Figure 4B have been proposed for studying potential trichogenesis. Among them the most promising configurations are those with the presence of ORSKs, DPCs, and dermal fibroblasts (DFBs) [Fig. 4B(e), (f)]. 53 In this model, a pseudo-dermis composed of collagen I mixed with interfollicular DFBs was created, and then on top of it either stacked layers of matrigel/DPCs and matrigel/ORSKs [“layered” system; Fig. 4B(e)] or single layer of mixed DPCs and ORSKs in the matrigel [“mixed” system; Fig. 4B(f)] was placed. In both systems, the ORSKs formed spheroid aggregates and retained their characteristic keratin expression patterns, but with better ORSK proliferation in the “mixed” culture. The kinetics of proliferation and apoptosis of DPCs were similar in both systems, as was the expression of their characteristic markers including versican.
Qiao et al. 55 have also tried to explore the formation of follicles using stem cells, in which cell aggregates from a mixture of embryonic mouse follicular dermal and epidermal cells were prepared using a hanging-drop method with the assistance of methyl cellulose. The extended culture of such cell aggregates led to the development of a “proto-hair,” further confirming the necessity of mesenchymal–epidermal interaction in folliculogenesis.
Compared with overly simple two-dimensional monolayer cultures, 3D culture models could more accurately replicate the in vivo environment favorable for follicle development. However, significant progress is still required to achieve the delicate hair follicle architecture. In any in vitro model, the obtained structures must be grafted into a host in order to demonstrate in vivo utility. 55
Creation of Hair Follicles in Skin Grafts
Preferably, tissue-engineered skin grafts should contain both epidermal and dermal layers.
56
The epidermal layer protects the wound from dehydration and microorganism invasion, and the dermal layer integrates with the wound bed upon grafting. Although no hair follicles have been regenerated in current skin grafts, the mesenchymal–epidermal interaction between epidermal and dermal layers would facilitate hair follicle regeneration. During graft fabrication, a 3D interconnected porous scaffold is often used to provide the initial surface for skin cells to adhere and grow and then define the final tissue shape.
9
Either made from natural polymers such as collagen, glycosaminoglycans, or chitosan or from synthetic polymers such as poly(
Temporary 3D scaffolds will be gradually replaced by newly formed skin tissue; however, the initial cell–scaffold interactions are crucial to functional tissue development. In recognition, a great deal of ongoing efforts focus on improving the scaffold design in order to better regulate the cells for desired phenotypes. The superiority of nanofibrous matrices in supporting tissue formation has been constantly highlighted mainly because of their size and morphologic similarity to native tissue ECM fibers. The flexibility of incorporating bioactive molecules such as collagen, elastin, and growth factors into the nanofibers adds another factor in recapitulating ECM composition. 58 The utilization of such nanofibers for skin regeneration has been steadily increasing. For instance, the use of polycaprolactone nanofibers containing type I collagen or gelatin can facilitate the growth of skin fibroblasts and keratinocytes and subsequently formation of skin grafts,59–61 even though current reports mainly focus on in vitro studies, and nanofiber-based skin grafts are not yet available for clinical application. A foreseeable need for hair follicle regeneration is to spatially arrange hair-forming cells to mirror the in vivo structure, 57 particularly to restore the trichogenic mesenchymal–epithelial interaction; however, the small pore size of these nanofibrous matrices (<5 μm) does not allow a complete infiltration of skin cells, resulting in nonuniform spatial distribution of seeded cells. Thus, diverse attempts have been made to seek a solution to this challenge. The layer-by-layer cell assembly method, established by our group, has proven to be effective in spatially organizing cells to achieve the uniformity. 62 In this method, a thin layer (5–10 μm) of nanofibers is first collected on the media surface, and then cells are evenly seeded onto the thin nanofiber layer. After cell seeding, another thin layer of nanofibers is electrospun onto the seeded cells. By repeating this process, 3D multilayered cell/nanofiber constructs can be created with great similarity to in vivo tissues where cells distribute in between ECM fibers. During this cell layering process, the flexibility to vary the cell density and type allows the spatial assembly of follicular cells together with interfollicular DFBs and epidermal keratinocytes into 3D constructs to form follicle-containing skin grafts.
In designing the in vitro hair follicle-inducing environment, it is critical to provide the cells with necessary chemical cues for both growth and differentiation. These factors can be either incorporated into the scaffold during fabrication 63 or supplemented into the culture media. Several major signaling pathways have been identified for their involvement in hair follicle formation. Work has shown that Wnt signaling, responsible for the formation of hair placodes, is necessary for maintaining the hair follicle-inducing ability of cultured DPCs.23,64 Although sonic hedgehog (Shh) signaling is not required for initiating hair follicle development, Shh signaling is essential for controlling growth and morphogenesis of the hair follicle. 65 It is reported that Shh expressed by hair follicle epithelium controls DP development, formation, and maturation via influencing both mesenchyme and epithelium beyond the hair germ stage of development. 66 On the other hand, overexpression of Shh suppressed the morphogenesis of hair follicles in embryos. 67 Platelet-derived growth factor-A has been found to be responsible for the formation of DP and DS in a synergic fashion with Shh. 68 Hepatocyte growth factor (HGF), a mitogen, 69 motogen, 70 and morphogen 71 for a number of different organs, 69 is also expressed by isolated and cultured human hair follicles72,73 and involved in hair growth. HGF has been shown to stimulate follicle growth and DNA synthesis in human hair as well as mouse vibrissae, 74 upregulate DNA synthesis in hair bulb-derived keratinocytes, 75 and modulate cyclic hair growth in mice. 76 Another factor involved in hair growth is insulin growth factor I (IGF-I). The use of both IGF-I (10 ng/mL) in conjunction with IGF-II (100 ng/mL) showed an increased ability over insulin in preventing the catagen stage in hair follicles. 36 Ascorbic acid-2-phosphate has been shown to have a stimulatory effect on DPC growth at a concentration of 0.25 mM, and this effect, however, was not shared by ORS cells or keratinocytes. 77 The drug of minoxidil was also found to prolong the anagen stage through proliferative and antiapoptotic effects on the cells. 78 The FGF and bone morphogenetic protein inhibitor, noggin, is among the other important signals involved in hair morphogenesis. 79 With increasing understanding of molecular contribution of various factors to hair follicle development, more efforts are required to synergistically incorporate these factors during the creation of a follicle-friendly environment, especially with nanofibers, in tissue-engineered skin grafts.
Perspectives and Challenges
The innate regeneration ability of hair follicles has motivated our attempts to explore the possibility of producing new hair follicles through tissue engineering and stem cell technology. Among several critical hair-inducing factors identified, in vitro emulation of the in vivo epithelial–mesenchymal interaction has been shown to be important. Therefore, more effective 3D approaches that recapture epithelial–mesenchymal interactions are needed. 80 The development of folliculoid organotypic systems has helped us to move along this path. 53 Layering of ORS cells on top of DPC-containing matrigel in a layered sandwich approach or the mixing of DPCs and ORS cells in matrigel in a mixed sandwich improves on previous 3D models (Fig. 4B). However, several challenges have emerged during in vitro culture. These include replicating neofollicles so that they produce hair in the same way as natural hair follicles, ensuring a high efficiency of hair follicle formation with appropriately chosen trichogenic cell populations, attracting other skin cells such as melanocytes and Merkel cells into the engineered hair follicles, guiding the migration of stem cells and progenitor cells to the proper areas, and regulating their differentiation.
Conclusions
The addition of hair and hair follicles to skin grafts provides a number of benefits. In addition to psychosocial effects, follicles can influence wound healing especially reepithelialization by generating short-lived “transient amplifying” cells from bulge stem cells and contribute to the ultimate success of skin grafts.81,82 To create hair follicles in tissue-engineered skin grafts, a 3D skin scaffold that incorporates the necessary conditions is needed. Although current 3D skin scaffolds meet the need for 3D architecture and environment, they still do not provide other conditions necessary to induce hair follicles, such as the presence of certain growth factors and hormones as well as the mesenchymal–epithelial and neuroepithelial interactions.14,16,83 As such, further work must be done to incorporate these factors into an in vitro 3D system for inducing the formation of follicle-like structures in skin grafts.
Footnotes
Acknowledgments
The authors apologize for not including many other colleagues' wonderful work in this manuscript because of page limitations. The authors are grateful to Mr. Lawrence Chan, Dr. Jason Fong, and Mr. Thomas Cattabiani for their editorial assistance. The authors acknowledge the financial support partially from NIAMS (Grant No. 1R21 AR056416) and by Innovation and Entrepreneurship Doctoral Fellowship from Stevens Institute of Technology.
Disclosure Statement
No competing financial interests exist.