
Abstract
Among the various types of cell-to-cell signaling, paracrine signaling comprises those signals that are transmitted over short distances between different cell types. In the human body, secreted growth factors and cytokines instruct, among others, proliferation, differentiation, and migration. In the hematopoietic stem cell (HSC) niche, stromal cells provide instructive cues to stem cells via paracrine signaling and one of these cell types, known to secrete a broad panel of growth factors and cytokines, is mesenchymal stromal cells (MSCs). The factors secreted by MSCs have trophic, immunomodulatory, antiapoptotic, and proangiogenic properties, and their paracrine profile varies according to their initial activation by various stimuli. MSCs are currently studied as treatment for inflammatory diseases such as graft-versus-host disease and Crohn's disease, but also as treatment for myocardial infarct and solid organ transplantation. In addition, MSCs are investigated for their use in tissue engineering applications, in which their differentiation plays an important role, but as we have recently demonstrated, their trophic factors may also be involved. Furthermore, a functional improvement of MSCs might be obtained after preconditioning or tailoring the cells themselves. Also, the way the cells are clinically administered may be specialized for specific therapeutic scenarios. In this review we will first discuss the HSC niche, in which MSCs were recently identified and are thought to play an instructive and supportive role. We will then evaluate therapeutic applications that currently try to utilize the trophic and/or immunomodulatory properties of MSCs, and we will also discuss new options to enhance their therapeutic effects.
Introduction
• In autocrine signaling, signals are transmitted to the producer cell. • In juxtacrine signaling, cells require direct contact and signals can be transmitted via gap junctions in the membrane. • In paracrine signaling, signals are transmitted only over short distances via factors that exert their effects locally. These factors are secreted by one cell and are only affecting neighboring cells due to fast degradation, consumption, or movement limitation by the extracellular matrix (ECM). • In endocrine signaling, signals are transmitted over long distances, via hormones secreted by organs and glands.
Paracrine signaling between stromal cells and parenchyma
In the human body, a great amount of tissues release signaling molecules such as growth factors, cytokines, and hormones to communicate, instruct, and to provide support to surrounding tissues. The interaction between stromal cells and tissues that they support may be a good example of such paracrine interactions. Not only do these cells function as a physical support layer, but they also provide cues for proliferation and differentiation. One of the most-studied networks is formed by hematopoietic stem cells (HSCs) and their progeny, and its supportive stromal system. Decades of research on the different anatomic compartments and cellular subtypes revealed that an intricate signaling network exists between the two, which on the one hand allows stromal cells to balance extensive immune activation within their direct proximity, and on the other to exert local homeostatic and instructive cues to hematopoietic, as well as epithelial and other cells types.
Under nonpathological conditions, these cues are tightly regulated, and may provide the basis for paradigms such as stem cell quiescence or the establishment of immune-privileged sites, via site-specific production of characteristic ECM components and local secretion of a vast array of soluble factors. However, in pathological conditions such as cancer, these cues might be misguided, to support tumor growth and metastatic spread. For example, in mouse models, mesenchymal stromal cells (MSCs) show a particular tropism for inflammatory and tumor sites, 1 where they potentially support cancer progression through secretion of immunomodulatory, proangiogenic, and prometastatic factors. 2 Whether this applies in humans is still a matter of intense discussion. 3
Recent reports pointed to a role for MSCs in the HSC niche, but besides that, these cells have also been investigated for their immunomodulatory and trophic properties for sometime now. We will therefore first discuss the specific role of MSCs within this intricate signaling network, then elucidate on their immunomodulatory and trophic properties, and eventually present different therapeutic approaches, which are based on exploiting the intrinsic paracrine functions of MSCs. At the end of this review, we will furthermore introduce new approaches with the potential to enhance the therapeutic efficiency of these cells, and we will also discuss current limitations and future perspectives of these therapies.
The HSC Niche
HSCs reside at specific locations in the bone marrow, where the very complex control of their proliferation, differentiation, and migration is regulated. A schematic representation of the cells and factors involved in the niche is shown in Figure 1. The idea of a so-called niche, where surrounding cells provide cues and instructive signals to control residing cells, was first proposed by Schofield. 4 Traditionally, the HSC niche was thought to consist of two compartments: an osteoblast and a vascular component. At the endosteal bone surface, HSCs colocalize with osteoblasts 5 and the number of long-term HSCs, which can contribute to hematopoiesis for a lifetime, was shown to correlate directly with the number of spindle-shaped N-cadherin+ CD45−osteoblastic cells, 5 thus providing an environment in which HSCs remain quiescent. Within the vascular niche, HSCs localize adjacent to sinusoidal endothelial cells, 6 supporting proliferation, differentiation, and subsequent migration into the bloodstream.7–9 However, recent research using high-resolution microscopy demonstrated the very close proximity of vessels and osteoblasts, thus suggesting that HSCs located in one compartment are subjected to paracrine factors from the other,9,10 and debate thus exists as to whether these two components form two physically separated compartments. As reviewed by Bianco et al. only 0.1%–7.3% of the endosteal surface of the human bone is actually covered by osteoblasts and they exist only for about 3 weeks, which would imply that the niche itself would be constantly migrating. 11
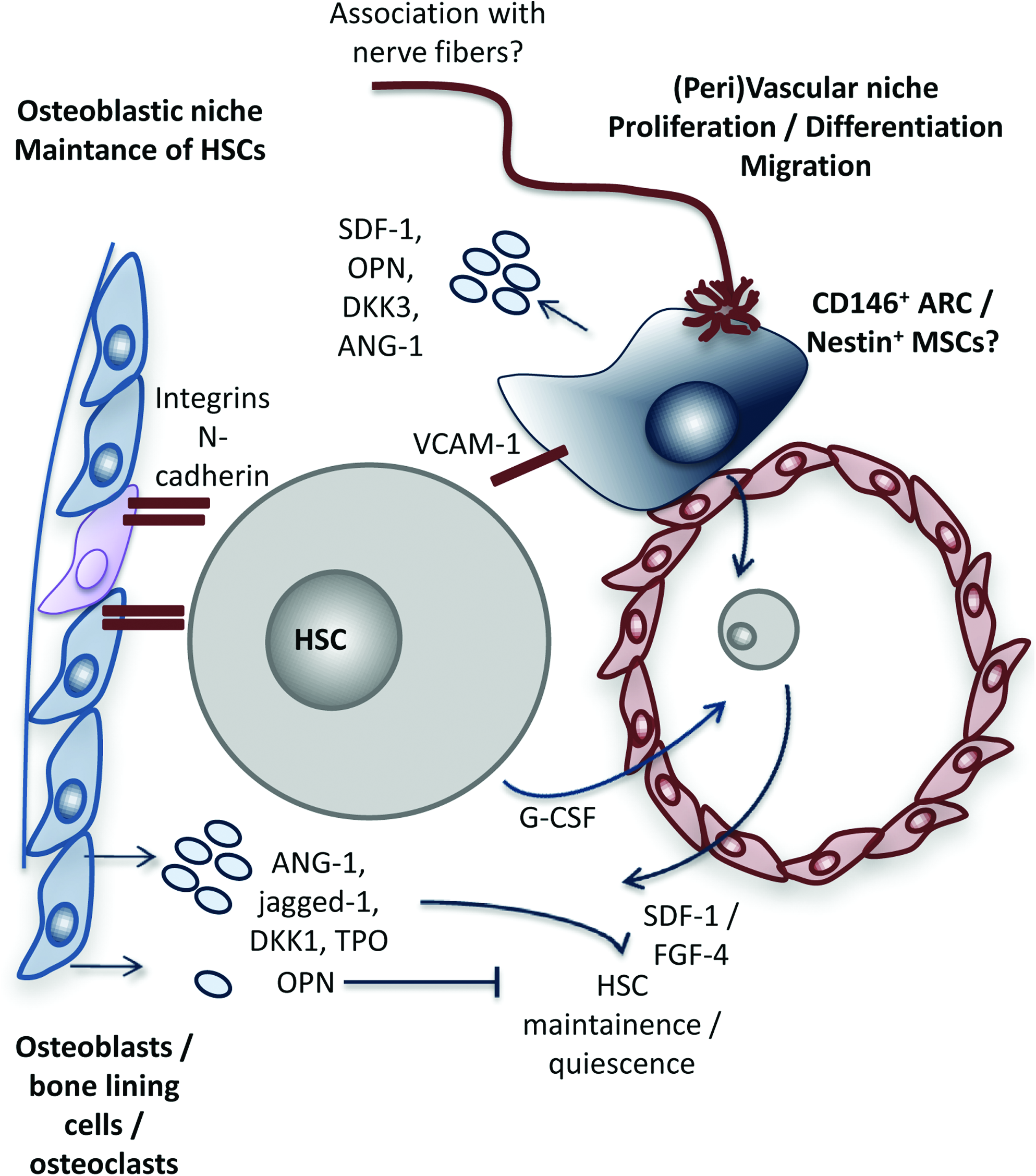
Schematic representation of the hematopoietic stem cell niche. Hematopoietic stem cells (HSCs) reside in a niche in which their maintenance, proliferation, differentiation, and migration are tightly regulated by various cell types. Traditionally, this niche was thought to be composed of an osteoblastic and vascular component, but recent data have led to a discussion as to whether these two are physically separated, and it thus likely that a HSC is in contact with both compartments. Cells in the osteoblastic component include osteoblasts, osteoclasts, but also bone lining cells, whereas the vascular compartment is mainly composed of sinusoidal endothelial cells, but CD146+ adventitial reticular cells (ARCs) and/or mesenchymal stromal cells (MSCs) also retain in this part of the niche. The HSC is retained in the niche by adhesion molecules such as integrins, N-cadherins, and vascular cell adhesion molecule-1 (VCAM-1). Osteoblasts are thought to secrete angiopoietin-1 (ANG-1), jagged-1, dickkopf 1 (Dkk1), and thrombopoietin (TPO), which keep the cells in a quiescent state, but also osteopontin (OPN), which antagonizes the effect of ANG-1. ARCs/MSCs express, among other genes, stromal cell–derived factor-1 (SDF-1), OP, DKK1, and ANG-1 and are thought to regulate the maintenance of HSCs and to control their migration. Migration into the bloodstream is induced by growth colony-stimulating factor (G-CSF), which is opposed by SDF-1 and FGF-4. Color images available online at www.liebertonline.com/teb
A role for MSCs within the niche?
Another cell type identified to be present in the HSC niche is CD146+ adventitial reticular cells, which were demonstrated to behave as colony-forming unit fibroblasts and are capable of osteogenic differentiation. 11 These cells were shown to share certain similarities with MSCs and to reside within a perivascular location in close proximity to HSCs. It was furthermore shown that MSCs and pericytes share a number of surface markers.12,13 Supporting this idea, recently, Méndez-Ferrer et al. described the presence of nestin+ cells with an exclusive perivascular distribution close to HSCs, which were characterized to be MSCs. 14 These cells showed high expression of HSC maintenance genes, appeared to regulate the maintenance of HSCs within the bone marrow and to control their migration toward the marrow. In addition, transcriptional analysis has identified the expression of several molecules specifically related to hematopoiesis in MSCs, such as fibronectin-1 (FN1), osteopontin (OPN), angiopoietin-1 (ANG-1), thrombospondin (TSP)-1 and -2, vascular cell adhesion molecule-1 (VCAM-1), fibroblast growth factor-7 (FGF-7), transforming growth factor (TGF)-β2, insulin-like growth factor (IGF) binding protein-4, bone morphogenetic protein (BMP) receptor type 1A, dickkopf-3 (DKK-3), secreted frizzled-related protein (Sfrp)-1 and -2, and CXCL12. 15 Furthermore, HSCs also maintain their niche by directing the differentiation of MSCs toward osteoblasts through secreted BMP-2 and BMP-6, 16 and they increase proliferation of osteoblasts, via platelet-derived growth factor-β (PDGF-β) and basic FGF (bFGF). 17 In this way, HSCs contribute to the regeneration of their own niche after, for example, irradiation.
Paracrine interactions within the niche
Supportive paracrine interactions between HSCs and stromal cells were already demonstrated in 1989 by Dexter et al. who showed that stromal cells secrete an array of hematopoietic cytokines, including growth colony-stimulating factor (G-CSF), stem cell factor (c-kit ligand), granulocyte-macrophage colony-stimulating factor (GM-CSF), and interleukin-6 (IL-6).18,19 Osteoblasts within the endosteal niche are thought to maintain HSCs in an immature state via production of factors such as ANG-1,20,21 thrombopoietin (TPO), 22 and DKK-1. 23 Their production of OPN may oppose quiescence via downregulation of ANG-1. 24 Maintenance of HSCs is furthermore facilitated by cell adhesion molecules such as VCAM-1, integrins, N-cadherin, and annexin II.25–27 Furthermore, production of stromal cell–derived factor-1 (SDF-1) supports homing of HSCs toward endosteum. 10 But G-CSF antagonizes SDF-1 expression in osteoblasts, to augment proliferation and mobilization of HSCs,28,29 which may be augmented further by a release of proteolytic enzymes.25,30 Application of G-CSF is therefore widely exploited clinically to mobilize stem cells into the peripheral blood.
Immunomodulatory and Trophic Effects of Therapeutic MSCs
Clinical trials using stem cells are expanding quickly. A search on www.clinicaltrial.gov shows the versatility in applications and cells used, including, but not only, HSCs, endothelial progenitor cells, c-kit+ cells, bone marrow mononuclear cells, whole bone marrow, and MSCs. Human MSCs are a heterogenous pool of cells and can be isolated from different sources of the adult body, typically the bone marrow. Guidelines, as set by the International Society for Cellular Therapy, to identify these cells include plastic adherence; expression of surface markers CD73, CD90, and CD105; lack of CD11b, CD19, CD34, CD45, and HLA-DR; and the ability to differentiate in vitro into at least osteoblasts, adipocytes, and chondrocytes. 31 Especially their nonimmunogenicity and their ability to differentiate into various tissues make these cells an ideal source for cell therapies as well as tissue engineering applications. MSCs secrete immunomodulatory factors as well as reparative and regenerative factors. Several studies have demonstrated differentiation of MSCs into different target cells, whereas low engraftment percentages, the short window in which MSCs exert their effects, and the fact that conditioned medium alone often exerts similar responses underline the particular importance of immunomodulatory and trophic mediators. 32
Trophic effects of MSCs
Already in 1996, it was noticed that isolated MSCs are able to secrete a broad spectrum of cytokines and growth factors that affect neighboring cells (Figure 3). 33 Specific growth factor panels and concentrations were found to vary between differently committed cells, but not between donors, donor age, and donor health. This trophic effect does not require differentiation of MSCs at target sites. 34 A selection of trophic factors implied in regenerative function is shown in Table 1. Work by Chen et al. showed that nonactivated mouse and human derived mesenchymal progenitor cells differ in their secretion proteome from more mature stromal cells, such as fibroblasts. 35 A higher expression was found in MSCs for most investigated growth factors, such as VEGF-A, ANG-1, TPO, hepatocyte growth factor (HGF), leukemia inhibitory factor (LIF), IGF, IGF-binding proteins-1, -2, -3, -4, FGF-4, -6, -7, -9, and a number of other molecules such as leptin, fractalkine, neutrophil activating peptide-2 (NAP-2), macrophage inflammatory protein-1β (MIP-1β), and MIP-3α, while a small number of cytokines (IL-6, -7, -8, and -10) and growth factors (G-CSF, M-CSF, GM-CSF, SDF-1, and SCF) was expressed at similar levels in both cell types.
HSCs, hematopoietic stem cells; MI, myocardial infarction; MSCs, mesenchymal stromal cells.
Immunomodulatory effects of MSCs
Apart from their high growth factor production, MSCs can modulate their microenvironment by locally suppressing immune responses via several direct and indirect mechanisms, which may potentially also lead to systemic shifts in immune activation (Figure 3). First, MSCs produce factors that decrease proliferation of CD4+ and CD8+ T-cells, B-cells, dendritic cells (DCs), and natural killer (NK) cells upon close contact.36–39 MSCs can also polarize the differentiation and function of myeloid cells such as DCs. 40 MSC-derived factors may therefore alter the maturation of antigen-presenting cells, 41 as well as the cytokine profile of various other cells. They may change the proinflammatory profile of T-helper1 (TH1) cells toward a TH2 cells' antiinflammatory profile and decrease the secretion of inflammatory proteins, such as tumor necrosis factor-α (TNFα) and interferon-γ (IFNγ) by DCs and T-cells, respectively. 42 These effects are mainly mediated by soluble factors, as summarized in Table 2, but are enhanced by cell–cell contact.43,44 The factors identified to mediate these processes include short-lived metabolites produced by enzymes such as prostaglandin E2 (PGE2), a product of cyclic oxide synthase (COX) 42 ; kynurenine, a product of indoleamine 2,3-dioxygenase (IDO) 45 ; biliverdin and carbon monoxide, the products of hemoxigenase-1 (HO-1); and nitric oxide, the product of nitric oxide synthase (NOS) 46 ; but also cytokines such as IL-10 and TGF-β,41–43 and a number of other factors such as HGF, 44 LIF, 47 and soluble human leukocyte-antigen-G5 (sHLA-G5)48,49 were identified as possible immunomodulatory mediators. More recently described molecules with immunoregulatory functions include CCL2 (MCP-1), galectin-1 (GAL-1), and TNFα-stimulated gene/protein 6 (TSG-6). MSCs were demonstrated to secrete high amounts of GAL-1, and by means of retroviral knockdown, GAL-1 was shown to mediate the antiproliferative effects of MSCs on peripheral blood mononuclear cells (PBMCs) and T-cells, but not on NK cells. 50 Furthermore, expression of TSG-6 was demonstrated to be highly upregulated after infusion of MSCs in mice with myocardial infarcts and siRNA against TSG-6 markedly reduced the beneficial effects of the infused MSCs on infarct size and heart function. 51 Similar antiinflammatory effects of TSG-6 were demonstrated in a mouse peritonitis model after stimulation with TNFα. 52 The CC chemokine ligand CCL2 mediates migration of inflammatory cells toward the spinal cord in autoimmune encephalomyelitis. The antagonistic form of this ligand, obtained via MSC-derived matrix metalloproteinase (MMP)-mediated cleavage, reduced secretion of inflammatory cytokines, suppresses proliferation of T-cells, and in vivo, MSC-secreted CCL2 was shown to suppress disease symptoms by preventing immune cells from infiltrating the spinal cord. 53
PBMCs, peripheral blood mononuclear cells; COX, cyclic oxide synthase; IFNγ, interferon-γ; TNFα, tumor necrosis factor-α; MCP-1, monocyte chemotactic protein-1; PBLs, peripheral blood lymphocytes.
Licensing and activation of MSCs
As outlined previously, MSCs produce factors that have so-called immunomodulatory or trophic effects on the cells in their direct proximity, and they can actively respond to the environment that they encounter, as depicted in Figure 2. To exert their paracrine function in an optimal manner, MSCs need to be activated, or licensed, which can be achieved by various external stimuli. Proinflammatory cytokines, chemokines, blood activation products, toll like receptor (TLR) ligation, but also physical cues such as oxygen tension, surface ligand interactions, and mechanical stimulation will change the secretion proteome and the amount of individual factors produced by MSCs, as well as their mobility and differentiation status.54–57
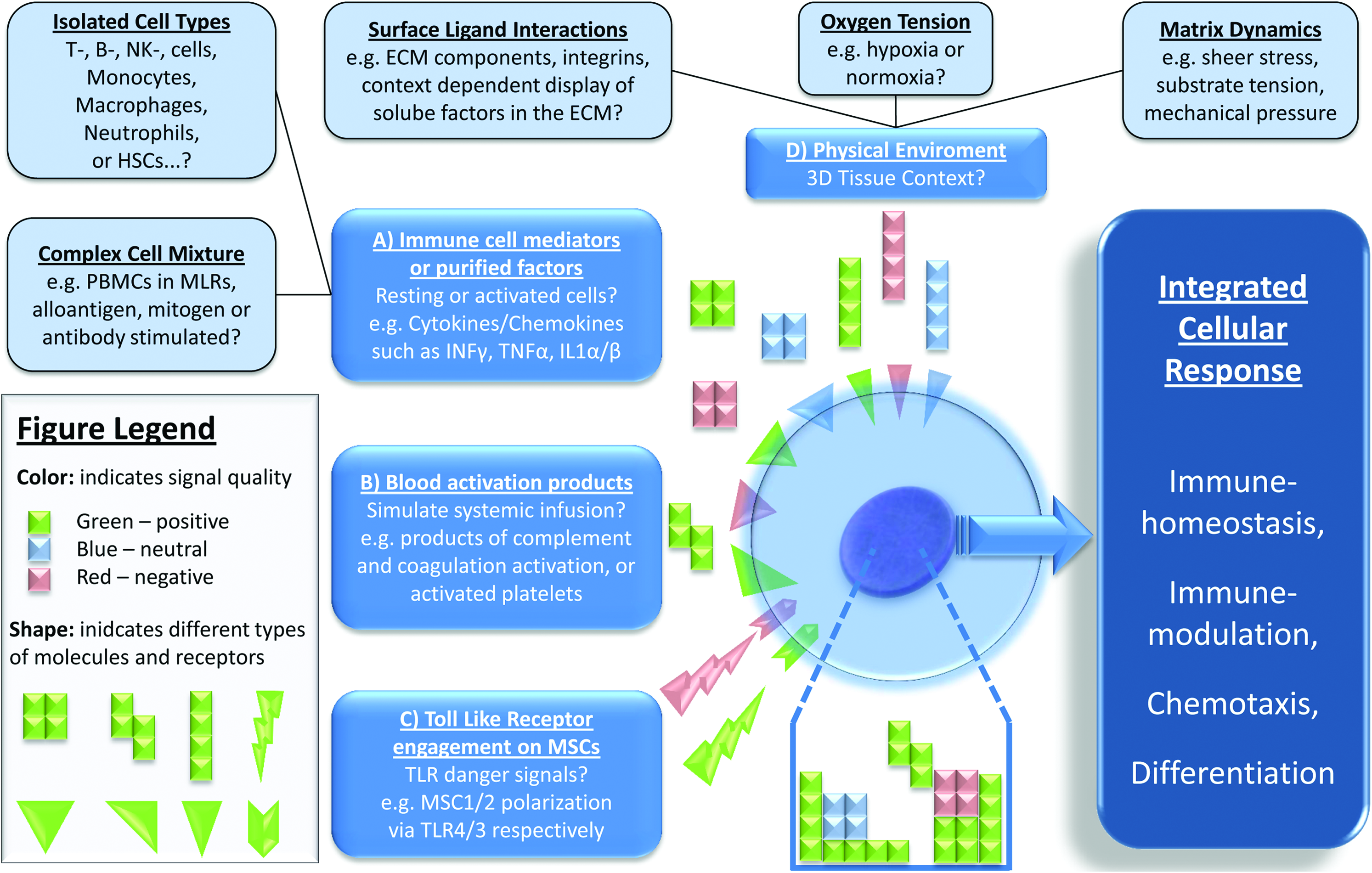
Licensing of MSCs by different stimuli. MSCs can be licensed by

Immunomodulatory and trophic effects of MSCs. Factors secreted by MSCs can have either immunomodulatory, or regenerative/reparative (trophic) effects. Immunoregulatory factors are depicted on the bottom and exert antiproliferative effects on T-cells, decrease secretion of antiinflammatory cytokines, alter the inflammatory profile of Thelper1 cells toward the more antiinflammatory Thelper2 profile, and increase the amount of antiinflammatory T-regulatory cells. The maturation of dendritic cells (DCs) is decreased, which is accompanied by a change in secretion profile. Trophic factors secreted by MSCs are depicted on the top and induce angiogenesis, increase mobilization of stem and progenitor cells toward the injury, enhance cell survival and proliferation, and support stem cells, whereas they decrease scarring (fibrosis) and apoptosis. In brain injuries, neural-specific growth factors can also exert neuroprotective effects. Color images available online at www.liebertonline.com/teb
Both IFNγ-independent and -dependent pathways appear to be important for triggering the immunomodulatory functions of MSCs. 58 IFNγ-independent priming leads to production of PGE2 via COX1 and -2, presumably due to engagement of soluble mediators, whereas stimulation of MSCs with IFNγ induces their expression of IDO, which suppresses T-cell activity through production of tryptophan metabolites and depletion of tryptophan from the local environment. Although MSCs can be activated by single proinflammatory mediators, combinations of IFNγ and either TNFα, IL-1α, or IL-1β potentiate their immunomodulatory effects. 46 Furthermore, work by Nemeth et al. demonstrated that, upon intravenous infusion into septic mice, MSCs are activated by both TNFα and LPS. Furthermore, their immunomodulatory function required cross talk with macrophages, which resulted in increased production of PGE2 by MSCs and IL-10 by macrophages. 59 This is only one example how activated MSCs may actively polarize adaptive and innate host resident immune cells, such as regulatory T-cells (T-regs), myeloid-derived suppressor cells (MDSCs), tolerogenic immature DCs (toll-DCs), or alternatively activated macrophages, to synergistically produce an antiinflammatory environment, as reviewed elsewhere. 60 However, MSCs are also subject to polarization themselves, which can occur in a TLR-dependent manner, resulting in generation of anti- or proinflammatory cells upon engagement of TLR3 and TLR4, respectively, which alters their migratory capacity and production of immunomodulatory cytokines. 61 Additionally, we and others have found that blood activation products, such as complement anaphylatoxins, clotting factors, and activated platelets, may have the potential to license MSCs, resulting in altered chemotactic, immunomodulatory, and ECM-degrading/infiltrating properties.62–64
Therapeutic Applications of MSCs
Infusion or injection of MSCs has been utilized in a wide range of applications and here we provide an overview of the performed work and the current status per application.
Graft-versus-host disease
The most advanced treatment protocol based on the immunomodulatory effects of hMSCs aims to treat graft-versus-host disease (GvHD). GvHD, currently treated with steroids, is caused by rejection of host tissue by transplanted bone marrow and can result in inflammations in the liver, skin, and gastrointestinal tract. Rejection is mostly due to CD4+ and CD8+ T-cells present in the transplant that produce excess amounts of cytokines, but activation of DCs, macrophages, and NK cells also plays a role. The disease is accompanied by increased secretion of TNFα, IFNγ, IL-1, IL-2, and IL-12. 65 Based on positive effects with infused MSCs in GvHD in various animal models, 66 several clinical trials were launched, with different outcomes. In 2008, we infused 55 patients with 1–5 doses of expanded bone marrow–derived MSCs in a phase II clinical trial. Thirty patients showed a complete response, 9 patients showed a partial response, and 16 patients showed stable or progressive disease. Patients with complete response showed higher survival than patients with partial or no response. 67 We have also reported on a single-patient case of a 9-year-old boy with treatment-resistant grade IV gastrointestinal GvHD, where intravenous infusions of ex vivo–expanded MSCs resulted in complete recovery of the gut and liver. 68 In another clinical trial performed by Osiris Therapeutics Inc., 3–21 intravenous infusions of culture-expanded allogeneic hMSCs resulted in complete remission in 7 out of 12 children after 1 month, and after 6 months, 95% of these children were still alive. 69 However, a larger, more recent phase III clinical trial performed by the same company did not result in any effect of infused MSCs as compared with placebo treatment, 70 which makes it difficult to draw any general conclusions, although on the positive side, adverse or side effects of infused MSCs were not reported in any of these trials. Data from animal models suggest that MSCs might act differently depending on their mode of activation,66,71 as supported by the fact that IFNγ prestimulated MSCs were more effective in combating GvHD than nonstimulated MSCs. 72 Further research is required to truly validate the effect of infusions of MSCs as an anti-GvHD therapy.
Crohn's disease
Also based on immunomodulatory mechanisms, MSCs may be useful in Crohn's disease, an inflammatory disease affecting the gastrointestinal tract. In a mouse colitis model, injections of adipose-derived MSCs resulted in increased survival rate, reduced inflammation, and increased weight, whereas injections of dead MSCs or human myoblasts did not have an effect. 73 In 2005, a clinical trial was performed, in which 35 patients with complex perianal fistulas received either fibrin glue or fibrin glue with adipose-derived MSCs. In the control group only 16% of the patients showed fistula healing, whereas in the group receiving MSCs, 71% showed healing. 74 As with GvHD, the data are inconsistent and another study showed less promising results. In a phase I clinical study including nine patients, only three patients were responsive to infusions of MSCs, whereas in three others the disease worsened. 75 Several other clinical trials are currently scheduled, for example by Osiris therapeutics, but disappointing preliminary results have reduced plans for late-stage trials. 70
Solid organ/graft transplantation
Systemic administration of MSCs could potentially also prolong survival of various grafts, which is currently achieved with continuous treatment of immunosuppressive drugs. Again, immunomodulatory factors suppress the immune response by acting on T-cells, DCs, and NK cells. In immunocompetent baboons infusions of allogeneic MSCs prior to graft placement slightly prolonged survival of a skin allograft 76 and the same was demonstrated with a heart allograft in rats. 77 On the other hand, another study showed that MSCs did not affect survival of a vascularized heart transplant in rodents 78 and a study in a mouse model demonstrated that MSCs only improved heart transplant survival in the absence of HSCs, which might explain failures in other studies. 79 Still, in combination with immunosuppressive drugs, administration of MSCs has demonstrated some promising results. A short course of low-dose mycophenolate mofetil (MMP) 80 or rapamycin 81 combined with administration of MSCs prior to transplantation resulted in long-term graft survival in rat heart transplant models. No studies in humans have been completed so far, but clinical trials have been approved to examine the effect of expanded MSCs on kidney transplants.
Myocardial infarct
MSCs secrete many growth factors that can have reparative and regenerative functions, of which most data come from studies in myocardial infarcts, although immunomodulatory factors also contribute to the repair. Already in 2001, Orlic et al. showed that transplantation of Lin-c-kit+ bone marrow cells resulted in the formation of new myocardium, consisting of infused cells. 82 Later, cardiac adipose tissue–derived progenitor cells were shown to engraft into the myocardium and to express both cardiac and endothelial markers, but in this case the secretion of proangiogenic growth factors suggested a trophic effect. 83 Debate thus exists on whether a therapeutic effect is due to trophic/immunomodulatory effects or due to direct differentiation into target cells. A trophic mechanism is supported by various studies that demonstrated a positive effect of injected MSCs, without showing evidence of engraftment or differentiation or that showed only very few engrafted cells.84,85 Often, conditioned medium derived from MSCs alone has positive effects as well. 86
A study by Zisa et al. showed beneficial effects of conditioned medium on heart function after myocardial infarcts, which was attributed to VEGF, IGF, or monocyte chemotactic protein 1 (MCP1). 87 Other known cardioprotective growth factors include FGFs, HGF, and VEGF, which are produced by skeletal muscle cells, and whose overexpression was shown to benefit the myocardium.88–90 Injection of MSCs resulted in elevated levels of both circulating HGF, LIF, and granulocyte/macrophage colony-stimulating factor (G/M-CSF), as well as the HGF, IGF2, and VEGF levels in the myocardium. MSCs appear to exert beneficial effects by attracting progenitor cells and by triggering growth factor secretion by host cells. 86 Implantation of MSCs into mice activates muscle cells to secrete VEGF via activation of IL-1β 91 and MSC conditioned medium contains IL-6 and LIF factors, which activate host myocytes to produce elevated levels of HGF and VEGF. 92 In line with this, Cho et al. demonstrated that growth factor expression in injected endothelial progenitor cells (EPCs) returns to basal levels 3–7 days after injection, whereas expression of FGF-2, Ang-2, HGF, IGF-1, SDF-1α and, to a lower extent, VEGF, ANG-1, placental growth factor (PlGF), and PDGF-β in host cells was still increased after 2–14 days. 93 These trophic factors are thought to exert effects on scar formation, apoptosis, vascular repair, angiogenesis, and the recruitment of stem cells and other regenerative factors.94,95
Regardless the underlying mechanisms, intracoronary administration of bone marrow–derived cells for myocardial infarction (MI) has been tested in several clinical trials, with various outcomes (reviewed in Ref. 96 ). Meta-analysis of 13 clinical trials, with a total of 811 patients, concluded that stem cell therapy improves left ventricular ejection fraction (LVEF), and reduces LV end-systolic volume and myocardial lesion area. No effect was found on postinfarction remodeling, which is considered a major predictor of late adverse outcomes. 96 Of these 13 trials, the Reinfusion of Enriched Progenitor Cells and Infarct Remodeling in Acute Myocardial Infarction trial was the largest one with 204 patients. From this trial it was concluded that death, the necessity for revascularization, or recurrence of infarction was reduced in patients administered with bone marrow cells after reperfused MI. 97 However, more recent trials show less encouraging results. In the HEBE trial, 200 patients were treated with either bone marrow–, peripheral blood-derived cells, or standard therapy, but no effects of either cell type on LVEF was found. 98 The REGENT trial included 200 patients, with a control group, a group receiving bone marrow nuclear cells, and a group receiving CD34+CXCR4+ cells. Again, no effects on LVEF or volumes were found, although a trend in favor of cell therapy was observed. 99 In the FINCELL trial, 80 patients received either mononuclear cells or a placebo and LVEF was found to increase more in the cell therapy group compared with the placebo. It has to be noted though that the absolute LVEF values after 6 months did not differ between the two groups. 100
The nonuniformity in the results throughout these clinical trials is thought to be due to several aspects. First, there seems to be a relation between the amount of the infused cells and the observed effects. Second, red blood cells and contamination of platelets in mononuclear cell fractions are suggested to affect functional behavior of the cell fraction, 96 and third, the site of injection plays a role; systemic infusion mainly results in uptake of cells by the lungs 101 whereas direct injection into the myocardium results in higher engraftment than a systemic approach. 96 Lastly, timing of the treatment is crucial. Activity of injected mononuclear cells was observed to be the highest within the first 24 h after a myocardial infarct, probably due to chemokines that are highly expressed shortly after the infarct. 102 In animal models, injection of MSCs almost always takes place directly after inducing the infarct, whereas in clinical trials, infusion often takes place within 6–12 months after the infarct, which might explain the more negative outcomes in clinical trials compared to studies in animals.
Wound healing
Trophic factors secreted by MSCs are also implicated in the acceleration of wound healing. Interactions between MSCs and endothelial cells, but also their immunomodulatory properties, seem to play a key role in accelerating wound healing and reducing scar formation. 103 Conditioned medium derived from MSCs was demonstrated to contain factors that promote recruitment of macrophages and endothelial cells into the wound. 35 Although the exact function of specific growth factors and cytokines was not revealed, a comparison between conditioned medium derived from MSCs and from fibroblasts (which does not affect wound healing) revealed clear distinctions between their secretomes, as outlined in “Trophic effects of MSCs” section. Conditioned medium promoted proliferation and cell survival of fibroblasts as well as production of collagen, elastin, and fibronectin in vitro. 104 Although evidence exists for the differentiation of MSCs into keratinocytes and endothelial cells,105,106 again, the low engraftment, the release of proangiogenic growth factors, 105 and the fact that conditioned medium alone also has substantial effects on migration, proliferation, and overall wound acceleration make a trophic effect a more likely explanation. 107 In rat and mouse models, injection of MSCs around the wound,108,109 but also systemic injection, 110 improved wound healing and a preliminary clinical study including 10 patients demonstrated that MSCs can be safely applied in chronic as well as acute wounds and that wound healing correlated with the amount of infused MSCs. 111 Another application includes cutaneous radiation syndrome, which occurs from overexposure to ionizing radiation. No controlled clinical trials have been performed so far, but Lataillade et al. have demonstrated possibilities of MSCs in several case studies. 112
Kidney failure
Kidney failure can occur as a result of damaging stimuli, such as sepsis-associated cytokines, toxins, and ischemia. Due to these stimuli, renal tubular cells can become apoptotic or necrotic and there is a loss of tubular epithelial cells. 113 Damage and swelling of renal endothelial cells, which leads to an impaired microvasculature system 114 and an inflammatory reaction, also contribute to kidney failure. Trophic factors released by MSCs can exert effects on the vasculature system but also reduce the immune response. Although integration of injected MSCs into kidney compartments has been reported 115 and it is believed by some that differentiation of bone marrow MSCs into renal epithelial cells is possible,116–118 studies have also indicated an effect with little or no integration119,120 and intraperitoneal injection of MSC conditioned medium had reparative effects as well. 121 In addition, Tögel et al. reported that reparative effects of injected MSCs were visible as early as 24 h after administration. 122 Paracrine and/or autocrine factors are thus likely explanations. Critical growth factors for this process include VEGF, 119 IGF-1, 123 and TGF-β, 124 which might activate recently revealed local stem cells in the adult kidney.125,126
Several animal studies have demonstrated that injections of MSCs have beneficial effects in both acute and chronic kidney failure. Injected MSCs migrate toward the kidney 120 and significantly improve renal function and regeneration, restore capillary defects, whereas they decrease apoptosis, renal injury scores, and mortality122,127 (for a detailed overview see Ref. 126 ). Observed effects were attributed to a reduction in the expression of proinflammatory cytokines (IL-1, TNFα, IFNγ, and NOS) and an increase in antiinflammatory cytokines (IL-10, bFGF, TGFα, and Bcl-2). 122 In contrast, infusion of MSCs did not affect renal injury in a sheep model, suggesting a species-dependent effect. 128 To the best of our knowledge, no clinical trials using MSCs to treat acute or chronic kidney failure have been completed so far, but one trial is currently ongoing and several others are scheduled.
Neuroprotective effects
In addition to immunoregulatory, proangiogenic, and antiapoptotic factors, MSCs also secrete neurotrophic factors, which could potentially be used in neurological disorders, such as stroke, amyotrophic lateral sclerosis (ALS), multiple sclerosis (MS), Alzheimer's disease, Huntington's disease, and Parkinson's disease (PD). These diseases are mainly characterized by a loss of neural cells, but also by occurrence of inflammatory reactions. MSCs were shown to be able to differentiate into neuron-like cells,129,130 but they also secrete neural growth factors, such as NGF, BDNF, and GDNF. Both stroke and PD are characterized by an inflammatory reaction, 131 but also by a loss of (dopaminergic) neural cells.132,133 Immunomodulatory factors secreted by MSCs thus act on the inflammatory response, whereas trophic factors may act to reduce infarct size, and to improve survival, functional recovery, and regeneration of neural cells.134–138 In addition, they can reduce apoptosis, improve vascularization, and attract progenitor cells to the site of injury. 139 Improved function after injection of MSCs intravenously or into the cerebellum in various diseases has been demonstrated in several animal models,136,140–144 but the exact underlying mechanisms remain to be elucidated.
In a small pilot study with five stroke patients, some improvements after injection of MSCs were demonstrated, but due to the small study number, no definitive conclusions could be made. 145 Similarly, a small, uncontrolled study with seven patients for treatment of PD showed encouraging results, but due to the lack of a control group and the small study number no conclusions could be drawn. 146 Another pilot study for the treatment of ALS including seven patients showed a trend toward slowing down of muscular strength decline, but again no conclusions could be drawn. A somewhat larger study with 21 patients for the treatment of MS and ALS reported fever as a small side effect of MSC injection, but no major adverse effects. They also showed slight increases in functional behavior. 147 Due to these encouraging results, several clinical trials are currently on the way or scheduled to test the use of MSCs in PD, stroke, ALS, and MS.
Functional Improvement of MSCs by Preconditioning
To enhance survival, grafting, and function of infused or implanted MSCs, cells can be pretreated or preconditioned prior to implantation, or specific genes can be overexpressed. Implantation of MSCs into an oxygen and nutrient-deprived environment, such as the myocardium, results in survival of very few cells. 148 Preconditioning of MSCs by culturing them in an hypoxic environment, which resembles the natural oxygen environment of the bone marrow (1%–7%) more closely than standard culture conditions (21%), 149 improves their survival via HIF-1α and Akt-dependent mechanisms. In addition, hypoxia stimulates the secretion of proangiogenic growth factors150,151 as well as expression of the chemokine receptors CX3CR1 and CXCR4 and enhances engraftment of MSCs in vivo. 152
Another popular approach to achieve secretion of growth factors or overexpression of function-specific genes is insertion of these genes into the cell by (non-)viral methods. Using these methods, Akt 148 or phosphoinositide-3-kase class II alpha (PI3K-C2α) 153 overexpression was shown to improve survival of MSCs in the infarcted myocardium. Heat shock protein-20 (Hsp-20), which interacts with Akt, also improves survival of implanted MSCs, probably through induced secretion of VEGF, IGF-1, and FGF. 154 Another example is overexpression of tissue inhibitor of matrix metalloproteinase-3 (TIMP-3), which was shown to improve cardiac function. 155 Since bone marrow cells are also affected by, for example, cardiovascular risk factors such as diabetes mellitus or hypercholesterolemia, cells collected from patients often have diminished therapeutic potential. Mees et al. demonstrated that overexpression of endothelial NOS (eNOS) rescues reduced neovascularization and the proangiogenic potential of bone marrow mononuclear cells from diseased mice. 156 Overexpression of eNOS was associated with increased differentiation potential of these cells into endothelial-like cells, increased secretion of VEGF, and enhanced NO-dependent vasodilation.
Third, MSC function can be improved by pretreating the cells to alter their secretome, either with growth factors or small molecules. Treatment with TNFα or endotoxin, for example, increases VEGF, FGF2, HGF, and IGF-1. 157 TNFα treatment also increases the expression of a panel of inflammatory cytokines in adipose MSCs, among which IL-6, IL-8, and MCP-1 were demonstrated to enhance migration of monocytes, 56 which suggests enhanced attractancy of these cells to the site of injury in vivo. Treatment with epidermal growth factor before infusion in ischemic limbs 158 was demonstrated to improve functional outcome, most likely due to an increase in secreted proangiogenic growth factors such as VEGF. 159 Similarly, induction of MSCs into a cardiac phenotype using a cocktail of various growth factors prior to injection resulted in improvement of functional cardiac behavior as compared with noninduced counterparts. 160 For applications in ischemic stroke, pretreatment of MSCs with brain extract resulted in enhanced secretion of BDNF, VEGF, and HGF. 57 Recently it was also demonstrated that even the culture protocol can alter growth factor secretion; culturing of hMSCs in spheroids enhanced secretion of antiinflammatory and antitumorigenic proteins as compared with conventional two-dimensional culturing. 161 Pretreatment with specific compounds thus can result in cells with a more specific secretome for a particular application or tissue.
Current Limitations and Future Perspectives
The research and clinical trials described here apply MSCs in various therapeutic applications. Research is expanding quickly and in addition to the applications described, recent articles have also reported the use of MSCs for corneal repair,162,163 pulmonary hypertension, 164 diabetes,165,166 and ischemic wounds. 167 Although injection or infusion of (trophic) MSCs has rendered promising results, data are often conflicting and hampered by different culture methods, isolation protocols, and amounts of infused MSCs and, therefore, have to be interpreted with caution. Many clinical trials are scheduled for the coming years and it is of crucial importance that future trials are setup in a controlled manner with proper control groups, although it might be difficult to obtain patient groups of sufficient size. In past studies many parameters fluctuated, which makes it difficult to compare results.
First, application sites vary between different studies, which could account for varying outcomes. For example, direct infusion of MSCs into the heart instead of systemic infusion was suggested to improve myocardial infarcts.96,101 Application of cells in close proximity to the target tissue can potentially be further improved by preventing migration of cells away from the tissue, for example, by implanting a hydrogel that encapsulates the cells. Second, it was shown that the window between myocardial infarct and MSC application is of significant importance, 102 possibly explaining discrepancies between clinical trials and animal studies. In line with this finding, different phases of axonal sprouting after stroke are initiated 168 and depending on the mode of action of infused MSCs, application timing may be crucial. Third, one of the major sources of variation lies in the cell type used. A wide range of cell types is currently under investigation, which makes it difficult to compare individual trials and moreover, isolation and culture protocols are not standardized. In myocardial infarcts, not only MSCs, but also c-kit+ cells and endothelial progenitor cells (EPCs), both derived from bone marrow, have been systemically infused or injected into the myocardium, with different success rates.93,169 Similarly, injections of HSCs did not affect kidney failure, whereas MSCs did 116 and, also in kidney failure, identical doses of autologous MSCs were shown more effective than allogeneic cells. 119 The best cell type for specific applications thus remains to be determined. In line with this, it has been shown that the contents of the secretome differ widely between cell types 170 and it thus remains to be discovered whether it is the differentiation and grafting, trophic and/or immunomodulatory effects, or a combination that accounts for the observed positive effect of infused MSCs. Activation or licensing of the cells prior to infusion also plays an important role here. In animal studies, it was demonstrated that cells from healthy individuals improve myocardial infarcts, whereas MSCs from diseased counterparts have no effect. Autoimmune diseases, as well as underlying causes of myocardial infarct and kidney failure, may affect bone marrow–residing stem cells, thus diminishing their therapeutic potential.156,171,172 This clearly demonstrates the need to investigate differences between these cells as well as the mechanisms underlying effects of injected cells. A better understanding of these mechanisms can help improve cellular performance and pretreatment of cells then can render their secretome more suitable for the intended application or, in contrast, induce differentiation prior to implantation.
For tissue engineering applications, the direct differentiation of MSCs is generally thought to be responsible for newly formed tissues, although in urinary tissue engineering, trophic effects of MSCs have been suggested to contribute to tissue formation. In this case, the use of a matrix led to induced secretion of IL-4 and TGF-β, which enhanced bladder wall remodeling and reduced expression of proinflammatory cytokines. 173 For bone and cartilage applications, attempts to enhance differentiation prior to implantation have not led to huge improvements in new tissue formation. Since MSCs also secrete bone- and cartilage-specific growth factors, trophic factors might very well play a role in these applications. So far, pretreatment of MSCs before implantation has been demonstrated beneficial for mainly two applications. As described, treatment with TNFα increases secretion of immunomodulatory factors whereas hypoxia treatment enhances secretion of angiogenic factors. Recently, it was demonstrated in our lab that treatment of hMSCs in vitro with the small molecule dibutyryl-cAMP (db-cAMP) results in more robust bone formation in vivo, which was accompanied by secretion of bone-specific growth factors such as IGF-1 and BMP-2. 174 These trophic factors are biologically active and affect proliferation and differentiation of various cell types in vitro, and are thus also likely to play a role in in vivo bone. 175 Determination of the origin of the newly formed tissues, donor or host, should elucidate whether and how these factors contribute to new bone formation. Besides db-cAMP, TNFα, or hypoxic culture conditions, other compounds, growth factors or small molecules could increase specificity or enhance secretion of specific growth factors even further. In addition, properties of substrates used to deliver the cells could be modified in such a way that cells will secrete growth factors that are useful for the intended application, for example, proangiogenic and antiapoptotic factors for myocardial infarcts and osteoinductive, but also proangiogenic, factors for bone tissue engineering. In our lab it was recently discovered that osteoinductive ceramics release calcium that induces expression of BMP-2 in hMSCs, possibly indicating a trophic mechanism behind osteoinductivity 176 (unpublished data). High-throughput screening may reveal novel compounds, but also optimal material properties like surface stiffnesses and topographies that induce differentiation and/or secretion of trophic factors for specific applications.
In conclusion, encouraging results in animal models as well as clinical trials strongly suggest an effect of the trophic factors secreted by MSCs in a wide range of applications. Application of MSCs in closer proximity to the target tissue as well as pretreatment to alter their secretome for specific applications could potentially result in further enhancement of MSC performance. However, additional research is required to unravel underlying mechanisms and determine optimal factors for specific clinical applications.
Footnotes
Acknowledgments
The authors gratefully acknowledge the support of the TeRM Smart Mix Program of the Netherlands Ministry of Economic Affairs and the Netherlands Ministry of Education, Culture and Science.
Disclosure Statement
The authors declare no conflict of interest.