
Abstract
Bioengineered tissue scaffolds are a potential tool for improving regenerative repair of damaged peripheral nerves. Novel modes of fabrication coupled with scaffold design strategies that are based on an understanding of the biology of nerve injury offer the prospect of intervention at a more sophisticated level. We review the etiology and incidence of peripheral nerve injury and the biological events that unfold during nerve regeneration after an injury. Newly available tissue scaffold fabrication technologies using bioplotting and laser-based techniques are described. Scaffold design strategies are also discussed, including the incorporation of living cells during scaffold fabrication, inclusion of neurotrophic gradients, use of electric stimulation, inclusion of antioxidant compounds to counteract neural apotosis, and promotion of angiogenesis. Use of these advanced fabrication techniques and incorporation of one or more of these active biological strategies may eventually lead to a greater success in peripheral nerve tissue engineering.
Introduction
Peripheral Nerve Injury
Incidence and etiology
Noble et al. 2 compiled the incidence and prevalence rates of peripheral nerve injuries over the preceding decade. Their study illustrated that almost 46% of nerve injuries were due to road traffic incidents. The radial nerve, a nerve in the posterior compartment of the forearm, was injured most frequently. Nerve injuries also commonly occur as complications of regional anesthesia. 3 In a retrospective study by Taylor et al., 4 the total incidence of peripheral nerve injuries due to upper- or lower-limb trauma was 1.64%, with crush trauma having the highest rate of associated nerve injury. Grade-III ankle sprains, arising mostly from sporting activities, lead to a significant incidence of nerve injury to the nerves in the leg, causing a prolonged rehabilitation time. 5 Compression neuropathies, such as carpel tunnel syndrome, can also induce chronic peripheral nerve injury. Moreover, some neuropathic disorders and tumors of the peripheral nerve require excision of part of the nerve.
Events in nerve repair
Functional losses after peripheral nerve injuries arise primarily from interruption of the integrity of sensory, motor, and autonomic axons. Crush or transection of the axon leads to reactive biological events in the neuronal cell bodies, the proximal segment of the injured nerve, the distal segment, and the peripheral innervation targets.
Axon transection separates distal lengths of axon from their parent cell bodies and results in a set of events in the distal peripheral nerve called Wallerian degeneration. 6 The lengths of axon in the distal nerve that have been disconnected from the neuronal cell body degenerate, although the remaining proximal lengths of axon remain largely viable. Wallerian degeneration entails a set of histological changes in the distal nerve that include disintegration of severed axons and their myelin sheaths and invasion of macrophages to clear degenerating cellular debris. 7 Beirowski et al. 8 showed that in mice, distal fragmantation of damaged nerves occurs within a short time frame of 40–44 h. Resident Schwann cells also phagocytize the myelin debris, 9 a phenomenon which has also been demonstrated in vitro. 10 It is thought that timely clearance of degenerating debris is necessary for peripheral axons to regenerate properly. 11 Disintegration of disconnected axons also appears to require activation of the intrinsic ubiquitin–proteasome system, which causes cytoskeletal disintegration. 12 The ubiquitin–proteasome cascade helps in the targeted degradation of proteins.
Three to 4 days after injury, Schwann cells throughout the distal stump and at the tip of the proximal stump increase their rate of division 13 and elaborately rearrange themselves to form bands of Büngner, the vital guidance cue for the regenerating axon. 14 Schwann cells help to guide the regrowth of axons toward peripheral targets to restore the nerve function. 15 Son and Thompson 16 show that axonal regeneration is limited if Schwann cells are prevented from extending processes. Schwann cells also produce various neurotrophins, such as brain-derived neurotrophic factor (BDNF), neurotrophin (NT)-3, NT-4/5, and nerve growth factor (NGF),17,18 which can promote neuronal survival and/or axonal growth.
In addition to events that occur near the damage site in peripheral nerves, axonal growth also depends on the type of neuron, age, and inherent regenerative capacity. 19 Injured neurons must switch from a metabolic state that supports the function of mature, stable axons to one that supports the formation of a motile growth cone expansion of the axon tip. Nerve injury results in the upregulation of various growth-associated proteins, including transcription factors, signalling proteins, cytoskeletal proteins, 20 and neurotrophic factors and their receptors. One example is the growth-associated protein-43 (GAP-43), a neuron-specific protein that undergoes elevated expression during nerve regeneration. 21 Partial depletion of GAP-43 in growth cones using antibodies 22 or antisense oligonucleotides 23 results in the reduction of F-actin, thereby impairing the motility function of growth cones. The synthesis of cytoskeletal proteins required for axonal reconstruction is also increased during regeneration. 24 Lund et al. 25 used in situ hybridization to show an increase in the β-actin mRNA levels, suggesting the importance of β-actin during axon regrowth.
Important extrinsic growth factors include the neurotrophins, insulin-like growth factors 26 , hematopoietic cytokines such as leukemia inhibitory factor 27 and interleukins, 28 fibroblast growth factor, 29 and transforming growth factor. 30 Extracellular matrix proteins such as laminin, fibronectin, collagen, matrix metalloproteinases, and cell adhesion molecules also support and regulate nerve regeneration through specific interactions with the cell surfaces. These matrix components can also interact with the growth factors and promote axonal advancement. Addition of antilaminin-2 antibodies to a culture of embryonic chick sensory neurons reduces the growth and total length of the neurites. 31 Thus, extracellular matrix factors, Schwann cells, and neurotrophins could potentially be exploited and integrated into scaffolds as biological factors that enhance regenerative success.
Newly sprouted axons enlarge and attain close-to-normal diameters with a greater myelin sheath thickness as they proceed toward the distal end. 32 Quan and Bird 33 report that remyelination of injured peripheral nerves takes between 2and 12 weeks, depending on the extent of the injury. Although the mechanism is not completely elucidated, it is known that many new axonal fibers grow, and those that do not achieve close proximity to the target are pruned away. This specificity of reinnervation, produced by selective prunning of axon collaterals, could be influenced by relative levels of target-derived trophic support 34 and by molecular cues on Schwann cells. 35
Fugleholm et al. 36 assessed axonal growth in cats by using implanted electrodes and found the regenerative growth rate to be 3–4 mm/day after crushing a nerve, but lowered to 2.5 mm/day in animals with sectioned nerves. Hoffman and Lasek 37 tagged rat cytoskeletal proteins with radioactive isotopes and found the actin in lesioned nerves to move distally at a rate of 2–5 mm/day. Buchthal and Kühl 38 studied the sensory potential, evoked by stimuli to the digits, in patients treated for a complete section of the median nerve. They found the rate of sensory axonal growth to be 1.5–2 mm/day. The rate of regeneration of peripheral axons is slower in humans than in rodents. 39 Studies on in vivo nerve regeneration have usually been done on acute models of injury. However, it is known that chronic denervation impedes the success of nerve regeneration, 40 suggesting that delayed treatment is likely to lead to a poorer clinical outcome. Better understanding of the molecular mechanisms that impede or delay nerve regeneration would be beneficial in designing tissue-repair scaffolds that are implanted at variable postinjury inervals to support axon regeneration.
Fabrication of Tissue Scaffolds
The emerging field of tissue engineering offers a new prospect for treatment strategies to foster regeneration of injured nerves, and thereby promote restoration of sensoy, motor, and autonomic functions. Tissue scaffolds are designed to serve as templates that support the regeneration of a tissue, organ, or specific structure. These scaffolds can be fabricated using a wide selection of biomaterials, ranging from hydrogels to composite polymers. Rapid-prototyping (RP) methods have advantages in the fabrication of tissue-engineered scaffolds over conventional techniques. 41 RP enables fabrication of scaffolds with greater pore interconnectivity and higher porosity, versatile control over the scaffold architecture, and discrete placement of different biomaterials. However, RP techniques such as fused-deposition modeling, selective laser sintering, and stereolithography require either high temperatures or the use of resin in the fabrication of scaffolds, making them incompatible with most biomaterials. Three-dimensional printing and three-dimensional bioplotting are examples of dispensing-based RP techniques that are more compatible with natural biomaterials and could be carried out under physiologically mild conditions, making the incorporation of biological materials, even living cells, within scaffold biomaterials more practical. 42 Indeed, this approach can allow specific and directed incorporation of living cells anywhere within the scaffold, which would be difficult to achieve by seeding living cells onto the surfaces of preformed scaffolds.
Computer-Aided Design-Based Tissue Engineering
Computer-aided design (CAD) is used extensively in the medical realm. Surgical reconstructions of large bone defects, especially cranial bones, are heavily dependent on CAD-based reconstruction. Using CAD techniques and stereolithography with computed tomography guidance enables the fabrication of implants with good results in the reconstruction of complex skull defects. 43 Similarly, CAD techniques are useful in the fabrication of joint prostheses; for example, Jiankang et al. 44 report the use of CAD techniques in fabricating and successfully implanting a customized tibial hemiknee-joint prosthesis in a 14-year-old girl. With increasing demands for biomemitic tissue scaffold designs, CAD techniques have been utilized in a number of recent fabrication methods (Table 1).
Three-Dimensional Bioplotting
As the name suggests, this technique plots scaffolds in three dimensions, by which a viscous biomaterial is dispensed through a nozzle into a liquid medium, thereby retaining its shape without any temporary supportive structures. Landers et al. 45 describe a procedure for effective dispensing of hydrogels using the three-dimensional bioplotting technique. To prevent the collapsing of the scaffold layers before gelation, the material is dispensed into a liquid medium with approximately the same density as the plotted material, allowing buoyancy to counteract gravity. Structural rigidity of thermoreversible hydrogels is achieved if the temperature of the receiving medium is below the gelation temperature of the hydrogel. They also suggest the use of roughened surfaces as substrates for the first hydrogel layer dispensed, for example, the use of sandblasted metal plates as receiving surfaces. Lee et al. 46 included rat embryonic (day 18) neurons in collagen and gelatin gels and then printed them into three-dimensional scaffolds. Low pneumatic pressures (<5 psi) and nebulization of the hydrogel crosslinker over each layer (once deposited) helped in achieving a good construct.
Pfister et al. 47 compare three-dimensional bioplotting with three-dimensional printing, finding the former to be more accurate and requiring less quantity of the dispensed biomaterial. Three-dimensional bioplotting is versatile and can be used to dispense different materials, including polymers, resins, and biopolymeric hydrogel solutions such as agar and gelatin. 48 Although plotting of most synthetic polymers using dispensing techniques could be challenging, polymeric formulations such as TEEX-29b (copolymer of L-lactide and D,L-lactide dissolved in trichloroethylene and tetrachloroethane) and TEEX-31b (poly-l-lactide-caprolactone in trichloroethylene and tetrachloroethane) may be used due to their bioabsorbability, ability to be ink-jet printed, and cellular adhesion properties. 49 However, the ability to print patterns with an adequate and controlled density of living cells or biologically active molecules would be precluded by the use of organic solvents in the fabrication process.
Laser-Based Fabrication Techniques
Laser-based techniques accurately deploy scaffold materials without the use of orifice- or nozzle-based dispensing. Thus, the process is immune to polymer adhesion and nozzle blockades. This approach also tolerates variations in the viscosity of the polymer or biomaterial solution. Laser-based methods are fast and can be more accurately controlled than most other techniques. Material deposition speeds are influenced by the laser pulse rate, stage-control accuracy, and the nature of the receiving substrate.
Two-photon polymerization
In two-photon polymerization (2PP), a beam of infrared laser is focused onto a photosensitive biomaterial, and polymerization is initiated by two-photon absorption. The greatest advantage of 2PP is that the absorption of photons from the laser by the material occurs only in the immediate vicinity of the focal point. 50 Using this approach, selective focusing of the laser within a three-dimensional space can bring about the fabrication of three-dimensional structures with high spatial resolution. 51 Biodegradable triblock copolymer scaffolds of poly(ɛ-caprolactone-co-trimethylenecarbonate)-b-poly(ethyleneglycol)-b-poly(ɛ-caprolactone-co-trimethylenecarbonate) have been made using 4,4′-bis(diethylamino)benzophenone as the photoinitiator with 4-μm resolution. 52 Two-photon polymerization of photosensitive materials are used to fabricate scaffolds with 100-nm resolution. 53 Lee et al. 54 reported micropatterning of a cell adhesive ligand (peptide RGDS) in poly(ethylene glycol-co-peptide) diacrylate hydrogels using a 2PP technique. Human dermal fibroblasts seeded into these hydrogels underwent migration selectively into the RGDS-patterned regions of the hydrogels. Livnat et al. 55 used multiphoton abalative laser to pattern fibrinogen hydrogel containing polyethylene glycol. Neurons could display motility within these constructs, and their processes elongated in specific directions, aligning themselves to the patterns made on the gel. Thus, similar techniques might be used in peripheral nerve engineering to guide migration of Schwann cells along bioadhesive pathways to form artificial bands of Büngner. Unfortunately, certain photoinitiators used in the polymerization technique can be cytotoxic. For example, acetophenone is toxic to cultured HepG2 cells at amounts <0.9 mg/mL. 56 Thus, caution is needed during the initial selection of hydrogels, photoresists, or photoinitiators used in this technique.
Laser-induced forward transfer
Laser-induced forward transfer (LIFT; Fig. 1) is a method that harnesses energy from nanosecond pulses of laser light and induces forward transfer of the material. When used specifically to transfer living cells from cultures, this technique is known as biological laser printing (BioLP). 57 BioLP is a nozzle-free technique that minimizes the cellular damage and maintains the cellular phenotype. 58 No direct laser interaction with the substrate occurs, and this technique is highly controllable and reproducible. Chen et al. 59 show that bovine aortic endothelial cells are almost 100% viabile with minimal effects from heat or shear stress when they are included during the BioLP process.
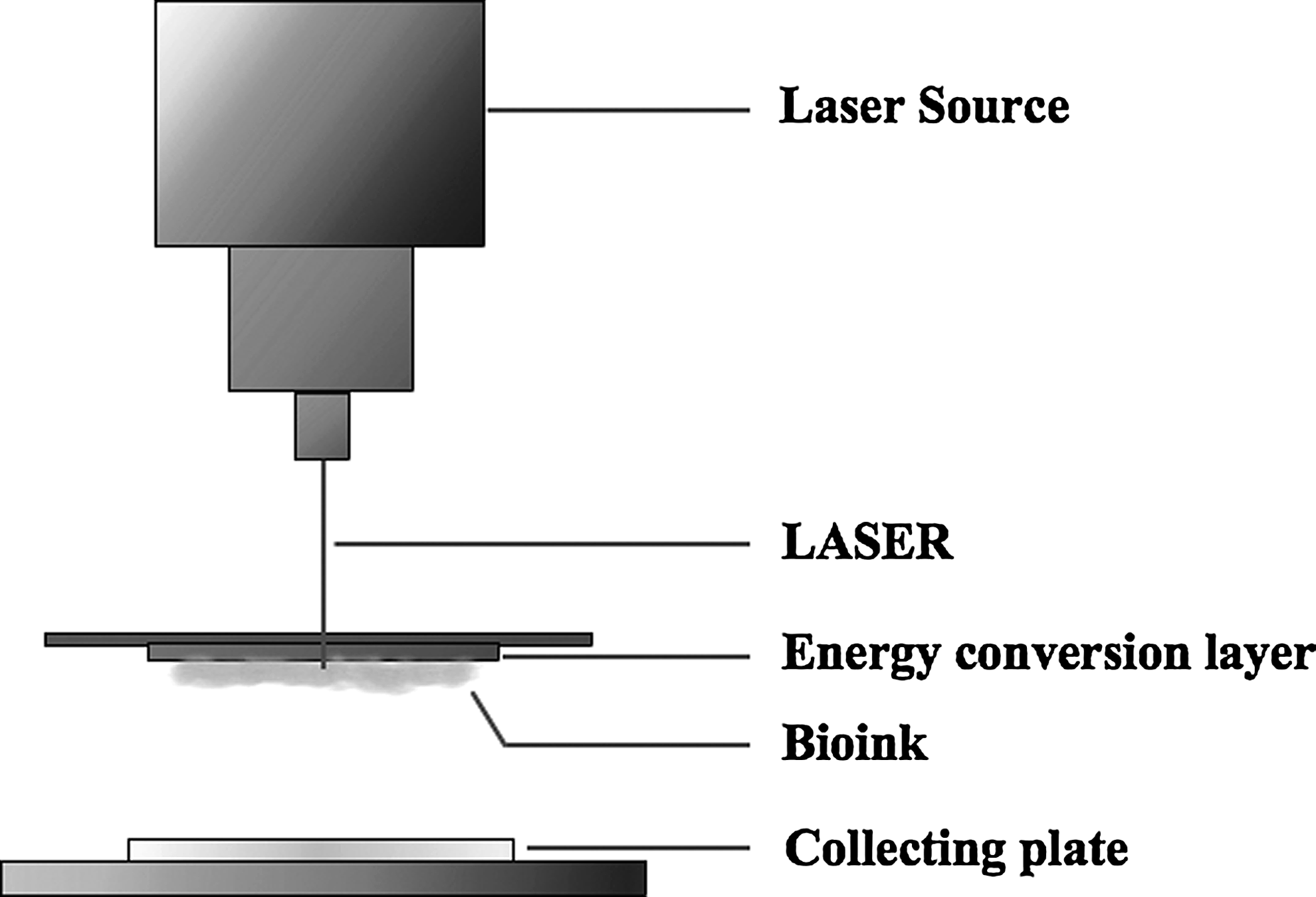
Laser-induced forward transfer.
Laser absorption onto any material causes an increase in temperature, leading to volatilization or even material expulsion. These material changes occur due to the thermoacoustical mechanisms initiated by photoabsorption of incident laser energy. BioLP utilizes an energy conversion layer consisting of a thin film of titanium oxide (TiO2), which absorbs most of the energy, thus preventing the cellular damage. The cells to be transferred are maintained in a hydrogel material known as bioink, positioned immediately below the TiO2 layer. Upon laser irradiation, the TiO2 layer absorbs most of the energy, and the heat induces displacement of cells from the bioink layer. Barron et al. 60 exposed the TiO2 layer to laser beams with a wavelength of 266 nm and estimated that 99.9% of the incident laser energy was absorbed by the TiO2 absorption layer. Materials printed from the bioink by this technique are therefore exposed to substantially less heat. 61 The absorption material must have a thickness greater than the penetration depth of the laser, but the transfer of heat through the laser absorption layer is independent of the absorption material thickness.
Ringeisen et al. 57 show good viability of olfactory ensheathing cells transferred onto a layer of Matrigel (a commercial mixture of exctracellular matrix proteins) and success with the co-printing of olfactory ensheathing cells with rat cortical neurons. Othon et al. 58 also demonstrate the capability of BioLP to print three-dimensional tissue scaffolds containing olfactory ensheathing cells, where the cells retain their phenotype and cell–cell interactions. This technique can transfer a volume of cells or other biomolecules at the picoscale. Barron et al. 61 demonstrate transfers as small as 0.5 pL and a droplet diameter as low as 30 μm. Dinca et al. 62 used BioLP to dispense SH-SY5Y human neuroblastoma cells and show the ability of these cells to orient, migrate, and produce organized cellular arrangements on patterns generated on polyethyleneimine. Hopp et al. 63 were unable to obtain 100% survival rates of transferred cells using BioLP (which they referred to as absorbing-film-assisted LIFT). This may have been due to their use of femtosecond lasers instead of nanosecond lasers for the transfer of cells. Although ultrashort laser pulses reduce the thermal effects on the cells, they likely cause a greater mechanical damage induced by the energetic plasma.
Matrix-assisted pulsed-laser evaporation direct-write
The matrix-assisted pulsed-laser evaporation direct-write (MAPLE DW) technique 64 is a microprinting tool that could help in the deposition of living cells within a scaffold biomaterial after fabrication of three-dimensional scaffold constructs. MAPLE DW is used to create microchannels, micropatterns, or networks within scaffolds. The microarchitectures created this way can be exploited for alignment of cells, as most cells have a tendency to align themselves along certain cues, such as the presence of extracellular matrix proteins 65 or fields of electric current 66 or also along the direction of small channels or patterns. 67 Harris et al. 68 created channels of various depths in gelled agarose by altering the aperture of the laser. The A431 human epithelial carcinoma cells and B35 neuroblast-like cells grown on these agarose scaffolds were subsequently found to align themselves in the direction of the microchannels. Laser direct-write has also been used to pattern mouse embryonic stem cell distribution within gelatin; 69 these cells maintained their stem cell pluripotency as evidenced by persistant expression of Oct-4. Creation of microchannels in scaffolds for peripheral nerve regeneration therefore constitutes another method that could be used to produce alignment of Schwann cells in a pattern akin to the bands of Büngner. Indeed, sculpting patterns into scaffolds using the MAPLE DW technique could be of a general value in the production of biomimetic cellular architecture to solve various problems in tissue engineering.
Strategic Design of Scaffolds for Nerve Regeneration
The architecture, material, and even the fabrication process used to create a tissue-engineering scaffold must be chosen with reference to, and with optimization for, the organ or tissue system being engineered. The unique structure and biological pathology of the peripheral nervous system impose certain constraints. Biomimetic approaches that seek to duplicate or enhance the normally favorable aspects of peripheral nerve regeneration are likely to be successful. 70 These include strategies to recreate endogenous healing mechanisms and appropriate mechanical environments within the engineered tissue. 71 Some strategies discussed here in the context of peripheral nerve regeneration might also prove applicable to other tissue-engineering aplications, such as the repair of central nervous system injuries. Some of the fabrication techniques discussed previously have already been implemented for the development of tissue-engineering strategies in other organs.
Incorporation of Cells Within Scaffolds
Previous studies have reported the incorporation of a variety of living cell types into scaffolds to support peripheral nerve regeneration, including Schwann cells, olfactory ensheathing cells, 72 or neural stem cells. 73 Living cells can either be seeded into scaffolds upon completion of fabrication or be incorporated during the scaffold-manufacturing process, provided that harsh conditions are avoided.
As decribed earlier, Schwann cells of the distal nerve stump align themselves into bands of Büngner, 14 and the importance of this arrangement in promoting axonal growth is well documented. 74 Since activated Schwann cells stimulate axon elongation 75 after a peripheral nerve injury, the incorporation of Schwann cells into tissue-engineered scaffolds is likely to be a beneficial strategy. Hall 76 shows that regenerating axons do not elongate through acellular nerve grafts in which the migration of Schwann cells is impeded. Li et al. 77 included Schwann cells and neural cells from the cortex of rats in composite scaffolds fabricated using alginate and gelatin. They found the cells to be viable for a week and also suceeded in seeding them in a gradient pattern. Dewitt et al. 78 incorporated Schwann cells into composite scaffolds of Matrigel and collagen. Although Schwann cells have a spherical morphology in collagen type I scaffolds, the addition of Matrigel resulted in increased elongation of Schwann cell processes (by 4.2 times), better survival of the cells, and superior mechanical properties.
Olfactory ensheathing cells are a type of macroglia exclusively located in the olfactory nerve that can continuously produce neurotrophic factors to mediate neuronal survival and axonal elongation. 79 In addition to enhancing axonal regeneration, 80 they can produce myelin under certain circumstances 81 and have better migratory potential to penetrate glial scars. Verdu et al. 82 show that bridging proximal and distal nerve stumps with a laminin-filled silicone tube does not repair resected rat sciatic nerves, but axonal regeneration is observed when olfactory ensheathing cells are seeded along with laminin. Olfactory ensheathing cells have also been examined for their potential use to promote spinal cord regeneration because of their ability to promote growth of axons across the central/peripheral nervous system interface.79,83
Macrophages have a pivotal role in clearing the myelin and cellular debris during Wallerian degeneration of distal nerve tissue, thereby facilitating axon regeneration through this tissue. 84 There is in vivo evidence that macrophage recruitment could be important for the increase in NGF synthesis seen after a nerve injury. 85 The peripheral nervous system receives ample infiltration by phagocytic cells (compared with the central nervous system), and these are not usually included in peripheral nerve tissue engineering. 70 However, enhanced recruitment of macrophages could be achieved by improving the vascularization of peripheral nerve scaffolds. 86
Beside the addition of supportive cells, their orientation in scaffolds is also necessary for the maintenance of adequate function. Alignment of myocytes in muscle repair scaffolds is pivotal for triggering adequate and forceful contractions along a specific direction. 87 Working with bone repair scaffolds, Kerschnitzki et al. 88 concluded that formation of oriented collagen matrices in bone, and in turn better mechanical strength, requires specific alignment of osteoblasts. In peripheral nerve repair, Seggio et al. 89 show that even in the absence of secondary guidance cues, a positive correlation exists between neurite outgrowth and orientation of the Schwann cells.
Random seeding of cells into scaffolds may be insufficient to permit subtle tracking of cell migration and reproduce biologically important cell–cell interactions. Also, close confinement of cells at a high density may result in regulation of specific cell signaling, resulting in reduced cell motility and proliferation. Advanced scaffold fabrication techniques such as laser-based deposition and bioprinting could help achieve precise cell deposition in tissue scaffolds, applicable to peripheral nerve regeneration. Othon et al. 58 showed that BioLP could be used to create three-diminsional hydrogel constructs with olfactory ensheathing cells confined in certain directions of the constuct strands. This oritented arrangement signicantly promoted the migration of the cells in the hydrogel, reaching up to 400 μm from their original printed position within 24 h. Bioprinting also helps regulate the spatial arrangement of cells during fabrication, and it could be exploited for the simultaneous use of different materials, thereby enabling construction of composite scaffolds. This may not be possible by means of conventional methods. With the bioprinting method, Schuurman et al. 90 fabricated composite scaffolds using cell-laden alginate and polycaprolactone. They showed that these composite scaffolds had better mechanical properties and also had the advantage of specific placement of various cell types in chosen patterns.
Incorporation of Bioactive Peptides Within Scaffold Materials
Naturally occurring proteins, such as laminin, 91 fibronectin, 92 and collagen 93 (or peptides derived from them), can be added to scaffold materials to enhance cellular activities. Laminin induces potent neurite outgrowth, and thus promotes nerve regeneration over long distances. 94 Laminin is a heteromer composed of three protein subunits (A, B1, and B2) that each contains functional domains mediating interaction with various cell surface receptors. 95 Synthetic peptides based on laminin sequences have been shown to have biological activity. 96 The peptides Tyr-Ile-Gly-Ser-Arg (YIGSR) and Cys-Asp-Pro-Gly-Tyr-Ile-Gly-Ser-Arg (CDPGYIGSR) on the B1 chain of laminin are involved in cell adhesion and migration. 97 YIGSR also prevents tumor metastasis. 98 The Ile-Lys-Val-Ala-Va (IKVAV) peptide sequence is found on the long arm of the A chain. This sequence is known to aid the growth of neurites. 99 The Arg-Gly-Asp (RGD) sequence in laminin is found in the short arm of the A chain. 100 This sequence is pivotal in cellular attachment through integrin receptors and is also a key sequence in another extracellular matrix protein, fibronectin.
Fibronectin is also a multimeric protein that is present in the extracellular matrix, but also exists as a soluble disulfide-linked dimer in plasma. 101 The fibronectin monomers consist of three different domains (F1, F2, and F3). In fibronectin, the RGD peptide sequence is found in a loop between the two β-strands of the domain F3. 102 The RGD sequence is the primary integrin-dependent cell-binding site on fibronectin and is associated with cell adhesion.
Specific peptide sequences from other extracellular matrix proteins also can aid in neuronal regeneration. Kajiwara et al. 103 investigated fetal hippocampal-derived neurospheres cultured on alginate gel. They observed adhesion and neurite outgrowth only when peptide sequences from the tumor necrosis factor receptor-1 protein were included in the alginate. The efficacy of peptides as growth-enhancing components depends on the expression of associated receptors in neural tissue.104,105
Itoh et al. 106 show that axonal growth cones and Schwann cells have a high affinity toward laminin, but may not recognize RGD sequences. However, the YIGSR sequence of laminin enhances the adhesiveness of Schwann cells. Successful bridging of transected sciatic nerves was seen in rats when these transected nerves were grafted with silicone tubes filled with either Type I collagen fibers coated with laminin or the YIGSR sequence. The YIGSR sequence is thought to be recognized by receptors present on the growth cone or Schwann cell, promoting their attachment and migration. Recently, laser-transfer fabrication techniques have been employed in the transfer of peptide sequences to enhance cell adhesion. Spots of REDV peptide spots were transferred onto polyethylene terephthalate (PET) surfaces using LIFT. Human endothelial cells were layered over these peptide spots, and the patterned peptide was found to secure better attachment. 107 Similarly, Zouani et al. 108 deposited RGD peptides onto PET surfaces using the LIFT technique.
Incorporation of Neurotrophic Factors
The neurotrophin family of signaling molecules, including NGF, BDNF, NT-3, and NT-4/5, were initially thought to act primarily as regulators of neuronal survival. It is now clear that these neurotrophins are required for a variety of signaling functions in the regulation of development, maintenance, and function of the nervous system. They also control cellular differentiation, axonal growth, dendrite pruning, and as well as the expression of various neural proteins and neurotransmitters. 109 Since the acknowledgement of their diverse functions and mechanisms of action, many studies have incorporated the use of neurotrophins in peripheral nerve tissue engineering.
Artificial polysulfone nerve conduit tubes incorporating NGF and the extracellular matrix adhesion protein laminin promote successful nerve repair on par with autologous nerve implants. 110 Synthetic polymeric scaffolds made of poly(lactic-co-glycolic acid) (PLGA) and coated with another neurotrophic protein, ciliary neurotrophic factor (CNTF), can repair 25-mm-long canine tibial nerve defects. 111 Lee et al. 112 used copolymers of N-hydroxyl succinimidyl ester pyrrole and pyrrole and found that the addition of NGF to the copolymers favored neurite extension in neuron-like PC12 cells.
The use of hydrogels in the delivery of neurotrophins is advantageous due to their sustained-release behavior. Katz and Burdick 113 studied nerve conduits filled with the hydrogel poly(2-hydroxyethyl methacrylate-co-methyl methacrylate) along with NGF-loaded poly(D,L-lactide-co-glycolide) microspheres. The result was a more sustained release of NGF compared to conduits without microspheres. The effective role of hydrogels as carriers of microspheres or other delivery methods is the subject of the continuing study. 114
Neurotrophins have also been encapsulated in microspheres made of various polymers such as chitosan, alginate, poly(lactic acid), poly(glycolic acid), poly(lactic-co-glycolic acid), and poly(caprolactone).115,116 Viral vectors have also been used to transfect genes that code for expression of neurotrophins. 117 Nonviral methods, such as transfection via lipoplexes, have also been used to introduce genetic material into cells. 118 Although a positive role of neurotrophins in peripheral nerve regeneration is widely recognized, a few studies indicate a potential dark-side of these proteins and their receptors. Specifically, NT-4/5 may exacerbate free radical-induced necrosis of neurons both in vitro and in vivo. For example, the neuronal death was accelerated upon addition of 10–100 ng/mL of NT-4/5 in neurons exposed to Fe2+ or L-buthionine-[S, R]-sulfoximine. 119 Park et al. 120 report similar effects with 100 ng/mL BDNF or NGF, with the damage dependent on the level of neurotrophins. Prolonged activation of TrkB, a receptor for BDNF, may have some detrimental consequences for neuronal survival. 121 Thus, the optimal quantity and temporal profile of release of neurotrophins from the scaffold for efficient neuronal re-growth have yet to be established.
Higher concentrations of neurotrophins could saturate cell surface receptors and lead to receptor downregulation. 122 Hence, controlled and graduated concentrations of neurotrophins (Fig. 2) could help as guidance cues to the growth cones of axons. 123 The axons are guided to their targets by a combination of contact-mediated and diffusible cues, either attractive or repulsive. 124 Cao and Shoichet 116 devised a model to estimate the minimal and maximal effective concentration gradients of NGF in a custom-designed diffusion chamber. The minimum concentration required to guide neurite outgrowth from PC12 cells was assessed and found to be 133 ng/mL per mm. Dodla and Bellamkonda 125 describe a polysulfone nerve conduit filled with agarose hydrogel imbued with different concentrations of laminin and NGF. The concentration varied between 8 and 33 μg/mL across four increasing step-gradients for laminin, and between 1.2 and 4.8×108 liposomal microtubules (containing NGF)/mL across similar increasing step-gradients. Regenerating axons were observed in rats implanted with these anisotropic scaffolds, but were absent in those implanted with scaffolds without gradients (isotropic). Moore et al. 126 created a gradient hydrogel scaffold using a gradient maker with two interconnected chambers; both were filled with p(HEMA), whereas only one chamber contained NGF. Spin bars blended both chambers as a peristaltic pump drove NGF-containing p(HEMA) into the p(HEMA)-alone chamber, followed by extrusion of a hydrogel containing an increasing gradient of NGF. Although this system is capable of establishing a general global gradient, the use of more recent fabrication techniques, such as bioplotting, permits better control over the configuration of gradients. Bioprinting techniques have been used to print spatially varying concentrations of neurotrophins by adjusting the printing resolution or the number of dispensed droplets in a given area. Ilkhanizadeh et al. 127 printed a gradient of increasing levels of CNTF on a polyacrylamide-based hydrogel and found a linear increase in the expression of the astrocyte marker glial fibrillary acidic protein by neural stem cells cultured on the surface.
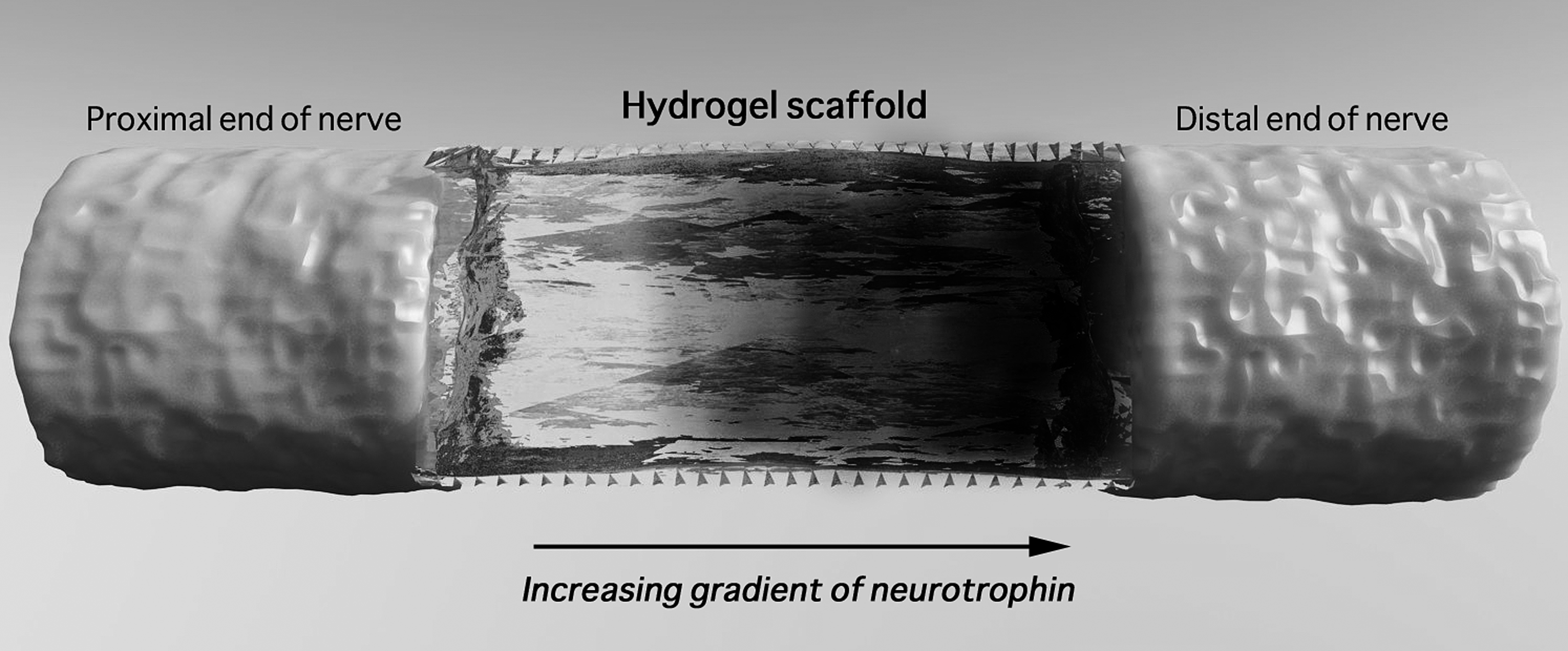
Schematic representation of a scaffold with a neurotrophic factor gradient.
Electrical Stimulation of Scaffolds
Artificially applied electrical stimulation can enhance peripheral nerve regeneration.128–130 A strategy of electrical stimulation might usefully be incorporated into the design of nerve repair scaffolds. Conductive polymers are a subgroup of materials with electronic and ionic conductivity, such as polyaniline, polypyrrole, polythiophene, and polyacetylene. The conductive property of these polymers is the result of certain dopant ions that help carry charges as electrical current. 131 These polymers have been used in the synthesis of biosensors, bioactuators, 132 and neural prosthetics. 133 Polyaniline and poly(D,L-lactide) electrospun at a ratio of 3:1 is electroconductive, and cells seeded over these scaffolds are able to proliferate. 134
Weak electrical currents can accelerate axonal growth in vitro. For example, PC12 cells grown on oxidized polypyrrole films show a twofold increase in neurite outgrowth when subjected to weak electrical stimulation. 135 The growth of axons in a constant electrical field in vitro is dependent on the charge of the substratum: axons move toward the cathode on negatively charged laminin surfaces and toward the anode on positively charged poly-L-lysine-coated surfaces. 136
Induction of weak direct currents by implanting an electric circuit in rats enhances nerve regeneration and increases blood supply, as seen by an increase in the number and diameter of the vasa nervorum 137 in association with enhanced axon regeneration. Thus, enhancement of vascular function (see below) may be a second beneficial outcome of elctrical stimulation in nerve tissue engineering.
The beneficial effect of electrical stimulation on axon regeneration in vivo is greater if electrical stimulation is initiated rapidly after an injury. Converesely, Yeh et al. 138 found better myelination in rat sciatic nerves stimulated 2 weeks postinjury than those immediately treated with weak currents. However, the effects of electrical stimulation are not always positive. In a study involving nerve regeneration through silicone rubber chambers, electrical stimulation hindered the growth of axons across the gap, although it accelerated the maturation of regenerating nerves if applied later. 139 Moreover, McConnell at al. 140 report local, deleterious, inflammatory changes in tissue surrounding chronically implanted electrodes, akin to late-onset neurodegenerative diseases. Hence, electrical stimulation could have both positive and negative effects, depending on the pattern, strength, and/or timing.
Addition of Antioxidants
Oxidative stress can lead to apoptotic death of neurons. 141 Apoptotic death of cholinergic neurons can be induced by NGF withdrawal, but this is prevented by high concentrations (50 mM) of the antioxidant ascorbic acid. 142 Rogério et al. 143 show that the antioxidant melatonin decreases neuronal death at doses of 1–50 mg/kg in rats. Acetyl-L-carnitine (ALCAR) also has inherent antioxidant properties and is involved in mitochondrial aerobic glycolysis. 144 In a study by Wilson et al., 145 long-term systemic administration of ALCAR at 50 mg/kg/day increased nerve regeneration after an injury and also resulted in greater myelination, improving the innervation of target organs. Carnosine (beta-alanyl-L-histidine), also an antioxidant, is protective for PC12 cells subjected to excitotoxic stress caused by N-methyl-d-aspartate overactivation of glutamate receptors. 146 Kilmartin et al. 147 used nuclear magnetic resonance spectroscopy studies to evaluate the antioxidant properties of polyaniline and polypyrrole and observed that these substances could neutralize the damaging effects of 2,2-diphenyl-1-picrylhydrazyl, a potent free radical. Thus, concurrent therapy with antioxidants, either administered systemically or included in nerve repair scaffolds, could be a useful strategy for tissue engineering targeting peripheral nerve repair.
Enhancement of Angiogenesis
Proximity of a nutrient source is pivotal in the regeneration or growth of any cell or tissue type. In vivo, most cells lie within one-to-three cell widths of a capillary. 148 Vascular compromise is prominent in most peripheral nerve injuries, including either the intrinsic or the extrinsic system of local blood vessels, or both. Incorporation of blood vessels within silicon conduits bridging nerve defects as long as 2–3 cm has been successful in rats. 149 Feeder blood vessels within artificial nerve conduits also help axons regenerate for longer distances. 150 In the current practice, a vascularized nerve graft, or one that has viable blood supply, is often used to bridge a gap between transected nerve stumps. This attempt to maintain blood supply can also help sustain the Schwann cell population and decrease fibroblast infiltration. 151 Podhajsky and Myers 152 show an increase in the endothelial surface area of local small blood vessels 1 week after a crush injury of the rat sciatic nerve, followed by a decrease at 3 weeks. However, the pattern was bimodal, and the endothelial surface area increased again 6 weeks after the injury. The initial increase was attributed to endothelial hypertrophy and the later increase to an increase in the number of blood vessels.
The importance of coexisting capillary growth in promoting nerve regeneration has been emphasized by others,153,154 but has received a limited direct examination. In a study comparing angiogenesis after a peripheral nerve injury, endoneurial capillaries were observed to sprout 5-mm distal to a nerve crush, repaired transection, or ischemic injury, but not after a resection injury that created a gap. 155 Neovascularization after a nerve injury could be promoted by signals arising either as a consequence of nerve ischemia or as an inherent aspect of the regenerative process. 156 Delayed vascularization in most tissue-engineered scaffolds is attributed to the absence of a pre-existing vascular network. Penkert et al. 157 show that Schwann cells can survive for only 1 week in a graft without blood supply.
Nitric oxide (NO) is a short-lived signaling molecule with multiple functions, including acting as a potent modulator of vascular growth. Three isoforms of nitric oxide synthase (NOS) are expressed in the peripheral nerve: neural NOS accumulating in growing axons of regenerating peripheral nerves, inducible NOS expressed in the invading macrophages, and endothelial NOS (eNOS) expressed by blood vessels in the vasa nervoram. 158 Nerve regeneration and vascularization are delayed in eNOS knockout mice when compared with wild-type mice. 159 Specific inhibition of eNOS using antisense oligodeoxynucleotides inhibits vascular remodeling. 153 Thus, incorporating a source of NO in scaffolds could be useful for peripheral nerve engineering to promote simultaneous establishment of new capillaries concurrent with axon regeneration.
Vascular endothelial growth factor (VEGF) is also a fundamental regulator of both normal and abnormal angiogenesis. 160 VEGF induces endothelial sprouting from sources of endothelial tissue embedded in a collagen gel. 161 Aizawa et al. 162 show that the movement of endothelial cells can be guided in three-dimesnional hydrogels by concentration gradients of VEGF. This could thus be a promising strategy to enhance blood vessel growth into artificial tissue scaffolds. Hobson et al. 163 assessed the effects of VEGF on the enhancement of vascularization and indirectly, on axonal regeneration in the damaged nerve. VEGF and the synthetic extracellular matrix product Matrigel were enclosed in a nerve conduit and implanted into rats to bridge a sciatic nerve transection. After 180 days, the number of regenerated and myelinated axons was 78% greater when compared to control conduits lacking VEGF. In another study, Lee et al. 164 constructed three-dimensional scaffolds incorporating murine neural stem cells, collagen, and VEGF-releasing fibrin gel. The neural stem cells in the closest proximity to the VEGF-releasing fibrin gel demonstrated better morphological changes as well as migratory responses. Thus, VEGF could have roles beyond angiogenesis in neural regeneration.
Novel delivery techniques for VEGF have also been explored. Kim and Kiick 165 propose the use of a specific polyethylene glycol–low molecular weight heparin hydrogel for carrying VEGF. VEGF was well contained in these hydrogels, due to its interaction with heparin, but was released for specific interaction with the VEGF receptor-2. Similarly, Golub et al. 166 demonstrated a sustained release of VEGF from poly(lactic-co-glycolic acid) (PLGA) nanoparticles, leading to enhanced blood vessel growth in mouse femoral artery ischemia models. Thus, patterned hydrogels could be synthesized to include bioactive substances enabling a sustained delivery at specific sites. The benefits of vascular growth in parallel to nerve regeneration are apparent in many studies, and bioengineering efforts to enhance peripheral nerve regeneration might include strategies that utilize factors and techniques that aid angiogenesis. With recent advances in fabrication techniques, supportive nerve cells and vascular endothelial cells could be printed simultaneously. Gaebel et al. 167 showed that LIFT-based seeding of cells enhanced cell survival and modified their growth characteristics, leading to increased blood vessel formation. Better formation of vessel-like structures was attributed to the precise control of arrangement of endothelial cells using the LIFT technique. Khalil and Sun 168 used a bioprinting fabrication system to print alginate scaffolds with a cell suspension of rat heart endothelial cells. Beside endothelial cells, VEGF has also been incorporated in fibrin gels and printed into collagen scaffolds using a bioprinter, to enable a sustained release of the growth factor. 164
Conclusions and Future Perspectives
Much progress has been made since the earliest use of biomaterials as simple conduits in nerve regeneration. Techiques to incorporate neurotrophins and control their release 169 or produce gradients 170 have been achieved. Material modification with peptide sequences has made scaffolds more adhesive, aiding in acclerated neural repair. Methods to promote adjunct angiogenesis 163 are also of key interest. Recent advances in the fabrication technology show a great promise for even more sophisticated, precise, and combinatorial strategies, thus generating a synergy, in peripheral nerve repair.
Although techniques to fabricate scaffolds with precise resolution are constantly explored or improved, methods to assess and visualize tissue growth or axonal regeneration in vivo are in need of improvement. Particularly useful would be the improvement of two-photon imaging, diffusion tensor imaging (DTI), and ultrasound-based techniques to enable better monitoring of cell distribution during the process of nerve repair. Two-photon excitation can be used to visualize autofluorescent cells seeded onto three-dimensional scaffolds, 171 where most of the seeded cells remain close to the scaffold surface. This technique might also help assess cellular distribution in more sophisticated scaffolds where cell distribution is micropatterned during the fabrication process, as the scaffolds need not be sectioned or manipulated for imaging. DTI is a magnetic resonance-based technique that provides a good imaging contrast and resolution to visualize nerve tracts. 172 This technique could be used to monitor nerve fiber content within scaffolds and along distal pathways toward peripheral targets. Hence, nerve regeneration in vivo could be assessed noninvasively. Ultrasound elasticity imaging is a technique that images objects based on the induction of distortions, which reflect the elastic properties of the object being investigated. 173 Speckle tracking has been used extensively in echocardiography, 174 and was recently used to noninvasively assess the degradation of poly(1,8-octanediol-co-citrate) scaffolds both in vivo and in vitro. 175 Further development of these techniques is needed for more effective monitoring and analysis of nerve regeneration in vivo.
The emerging fabrication techniques are themselves poised to be refined further for the advanced design of artificial scaffolds for applications such as peripheral nerve tissue engineering. Novel methods can be developed to simultaneously incorporate biomaterials such as growth factors or adhesive peptides along with living cells or other associated components that would closely mimic the natural environment and physiology of the tissue targeted for repair. Research aimed at understanding the cellular mechanisms of repair and at further improving the engineering of biofabrication is likely to overcome some of the short-term impediments to the successful use of scaffolds to promote the improved repair of peripheral nerve injury.
Footnotes
Acknowledgments
The authors would like to acknowledge the financial support from the Natural Sciences and Engineering Research Council of Canada (NSERC) and the Saskatchewan Health Research Foundation (SHRF).
Disclosure Statement
No competing financial interests exist for any of the authors.