
Abstract
The adult body harbors powerful reservoirs of stem cells that enable tissue regeneration under homeostatic conditions or in response to disease or injury. The hair follicle (HF) is a readily accessible mini organ within the skin and contains stem cells from diverse developmental origins that were shown to have surprisingly broad differentiation potential. In this review, we discuss the biology of the HF with particular emphasis on the various stem cell populations residing within the tissue. We summarize the existing knowledge on putative HF stem cell markers, the differentiation potential, and technologies to isolate and expand distinct stem cell populations. We also discuss the potential of HF stem cells for drug and gene delivery, tissue engineering, and regenerative medicine. We propose that the abundance of stem cells with broad differentiation potential and the ease of accessibility makes the HF an ideal source of stem cells for gene and cell therapies.
Introduction
HF Biology
The HF is part of the pilosebaceous unit that contains the sebaceous gland, the apocrine gland, and the arrector pilli muscle. The HF is composed of two main compartments: the upper part includes the infundibulum and the isthmus, whereas the lower part is comprised of the bulb, matrix, and dermal papilla (DP). The exact position of the lower part varies during the hair cycle. Adjacent to the lower part of the infundibulum lies the sebaceous gland that waterproofs the skin by secreting sebum. The isthmus contains the hair bulge, which is believed to be the stem cell reservoir regenerating the HF under homeostatic conditions or following injury. Cells migrate from the bulge toward the bulb, where they proliferate and differentiate to produce the hair shaft and all the epithelial cells that constitute the HFs. 2 Finally, the inner and outer root sheaths are composed mainly of keratinocytes surrounding the hair shaft (Fig. 1).

Schematic drawing of hair follicle. DS, dermal sheath; ORS, outer root sheath; IRS, inner root sheath; DP, dermal papilla; SG, sebaceous gland; APM, arrector pilli muscle. The illustration is not drawn in scale. Color images available online at www.liebertpub.com/teb
HFs undergo numerous cycles of growth and retraction throughout life. This dynamic process in adult life has three distinct phases—anagen, catagen, and telogen—each regulated by different signals. Anagen is the growing phase. Stem cells that are located in the bulge region differentiate to all hair lineages, resulting to hair elongation. The duration of anagen varies depending on the anatomic location of the follicle. On the scalp, anagen may last as long as 8 years resulting in long hair, but in other places, such as the eyebrow, anagen maybe as short as 3 months. Catagen is the regression phase. At this stage, the majority of the HF cells undergo apoptosis causing shortening of the lower compartment and bringing the DP cells closer to the bulge. Exchange of signals between the papilla and the bulge regulate the duration of catagen. The cells that escape apoptosis during this phase comprise the reservoir that leads to the next anagen. Telogen is the last phase of the HF cycle, also known as the resting phase. In telogen, cells enter a quiescent state waiting for the necessary signals to restart the cycle via the anagen. It is estimated that at any given time 5%–15% of HFs in the scalp remain in telogen.1,2
Location and Differentiation Potential of HF Stem Cells
Stem cells are characterized by three unique properties: self-renewal, capacity to differentiate into one or multiple cell types, and the ability to form tissues in vivo. Based on their differentiation potential, they can be classified as totipotent, pluripotent, multipotent, and unipotent. Totipotent cells can reproduce all the cells of a living organism. Pluripotent cells can also give rise to all tissues of the body, but unlike totipotent cells, they cannot form the extraembryonic tissues, which are necessary for proper growth of the embryo. Multipotent stem cells have even more restricted differentiation potential, but they can still differentiate into more than one cell type. For example, mesenchymal stem cells (MSCs) can be coaxed to differentiate into multiple tissues, including fat, bone, cartilage, and muscle. 6 The lowest in the hierarchy are the unipotent stem cells or progenitor cells, which can generate only one differentiated cell type. Stem cells can also be classified into embryonic or adult according to their origin. 7 Although embryonic stem cells (ESCs) have a broader differentiation potential, adult stem cells can be isolated from the patient directly overcoming possible immune rejection after transplantation.
In vivo adult stem cells reside in multiple tissues usually in a well-protected microenvironment called niche. Examples of adult stem cell populations include the intestinal stem cells, the neural stem cells, bone marrow-derived stem cells, and the HF stem cells. Although in vitro stem cells proliferate markedly in response to appropriate signals in the culture media, in vivo they remain quiescent until they are coaxed to proliferate and/or differentiate only when needed, for example, in case of injury. Under homeostatic conditions, the stem cell pool is maintained through asymmetric division, where the parent stem cell divides into two cells with varying differentiation potential: one retaining the stem cell characteristics (self-renewal), and the other assuming a more differentiated phenotype (differentiation). This way, the body holds a powerful reservoir of cells that can be readily mobilized in case of emergency, for example, injury to expand the stem cell pool through symmetric division, where two stem cells are generated from each parental stem cell. 8
Similar to other organs, the HF contains a rich pool of stem cells residing in various anatomic locations within the HF, prompting some scientists to call the HF a stem cell zoo. 9 In the next chapter, we will present the different stem cell populations, we will elaborate on their broad differentiation potential, and their interactions in the HF microenvironment.
Bulge and hair germ
Due to the complex architecture of the HF, the location of the stem cell reservoir remained elusive for many years. Initial studies reasoned that stem cells resided in the bulb, 10 but this hypothesis was abandoned, as removal of the bulb did not inhibit generation of new HFs. 11 In the early 1990s, Cotsarelis et al. were the first to propose that stem cells reside in the bulge area of the HF. They took advantage of the fact that in vivo stem cells cycled very slowly so that long time after administration of tritiated thymidine, only the cells that retained the label (label retaining cells) were the slow cycling stem cells.12,13 Several years later, this finding was verified using transgenic mice that were engineered to express the fusion protein histone H2B-GFP under the Krt5 promoter in a tetracycline regulatable manner. As a result, skin cells expressed GFP except when the mice were fed Doxycycline, which suppressed GFP expression. Once GFP expression stopped, the protein disappeared from the fast cycling cells, but was retained in the slow cycling stem cells that were localized in the bulge region of HFs. 14 Additional tracing studies with transgenic mice expressing the lacZ transgene under the control of either the Krt5 or Lgr5 promoter further supported the bulge activation hypothesis, which states that during anagen, stem cells from the bulge migrate in the bulb region where they are induced to proliferate and differentiate to all epithelial cell types of the HF.15,16
Notably, transplantation of KRT15+ or LGR5+ cells along with dermal fibroblasts in the dermis of nude mice generated new HFs with high efficiency.15,16 On the other hand, damage of the bulge from the autoimmune disease Lichen planopilaris resulted in permanent hair loss, 17 further highlighting the importance of bulge-derived stem cells for hair regeneration. Finally, experiments with single-cell clones showed that single bulge cells were highly clonogenic and after expansion and transplantation they were capable of generating intact follicles in vivo,18,19 providing strong evidence that the bulge harbors true stem cells and not a collection of progenitors.
Although it is widely accepted that the bulge harbors stem cells, the exact stem cell population is still under debate. Jaks et al. challenged the notion of label-retaining cells as the true stem cell population in the HF, as LGR5+ cells can regenerate the whole follicle, but do not coincide with the label-retaining cells of the bulge. The same study reported that LGR5+ cells were found in the hair germ, a region between the DP and the bulge, which remains discrete during telogen, but overlaps with the matrix during anagen. 16 In a recent review, Li and Clevers argue that the bulge contains both active and quiescent (label retaining) stem cells. 20 The former express CD34 and LGR5, whereas the latter are CD34+ LGR5−. 16 LGR5+ cells seem to be the first to respond to the inductive signals in early anagen and start proliferating and differentiating. 16 On the other hand, the slow cycling stem cells located in the bulge may not contribute to hair cycling, but get activated only if needed (e.g., injury).
Others believe that the hair germ originates from the bulge and contributes to the generation of the new HFs in the beginning of anagen. 21 In agreement, Greco et al. showed that the transcriptional profile of hair germ cells resembles that of bulge cells. They also found that hair germ cells proliferate faster than bulge cells and respond first to the DP signals at the late telogen. However, they also lose their proliferative capacity faster than bulge cells during long-term expansion in vitro. 22 Using a sophisticated tracing method, Hsu et al. reported recently that during anagen some stem cells exit the bulge, but remain slow cycling, retain the stem cell properties, and become the reservoir of stem cells for the next cycle. The same study also demonstrated that although some cells are committed to differentiate, they still return to the bulge. Surprisingly, these lineage-committed cells expressed stem cell markers, but failed to regain stemness; instead, they appeared to regulate the activity of bulge stem cells by secreting key factors. 23
Additional studies showed robust multipotency of bulge stem cells in vivo, where they were found to participate in angiogenesis, and in vitro, where they were coaxed to differentiate into neurons, glial cells, melanocytes, keratinocytes, and mesenchymal cells.24–28 In addition, Yu et al. reported that single bulge cells can differentiate into mesenchymal as well as neuronal lineages in vitro. 28
In addition to hair regeneration, bulge stem cells were found to contribute to wound healing following skin injury by migrating and differentiating into epidermal keratinocytes.29,30 However, they are not necessary for the maintenance of the epidermis (ablation of the cells does not affect the homeostasis of the epidermis) and in the long run they fail to stay at the sites of injury. 30 Others showed that HFs also harbors melanocytes, which differentiate and produce melanin during each hair cycle stimulating hair pigmentation. Nishimura et al. identified melanocyte stem cells in the bulge and sub-bulge areas. These cells were slow cycling, undifferentiated cells that were activated during anagen to produce melanocytes. 31
Isthmus/infundibulum
Cells located above the bulge are believed to retain multipotent properties. Studies have reported that they can differentiate not only into the epithelial lineages of the HF, but also into the sebaceous gland and the epidermis. However, it is not yet known whether these cells represent a unique stem cell population, a subset of bulge stem cells, or even progenitors with limited differentiation capacity. Isolated cells from the area between the bulge and the sebaceous gland were found to be distinct from the bulge-derived stem cells since they did not express bulge-specific markers, such as KRT15 and CD34. Although they maintained high clonogenic potential in vitro, they were also actively proliferating in vivo—in contrast to the notion that in vivo stem cells are the slow cycling, label-retaining cells. 32 Similarly, Jensen et al. reported that cells of the upper bulge region were not quiescent in vivo, but they could generate new follicles after implantation in vivo, suggesting that quiescence might not be a requirement for maintaining multipotency. 33 Several other studies showed that cells derived from a region above the bulge can give rise to epidermis and persist there for a long time following injury.33–35
Sebaceous gland
There are two theories with regard to the origins of the sebaceous gland. The first asserts that stem cells residing in the bulge region migrate and give rise to resident gland cells. This theory is supported by transplantation studies showing that bulge cells generated functional sebaceous gland in vivo.15,16 The second theory suggests that stem cells located above the bulge differentiate into sebocytes.34–36 Horsley et al. bridged these two theories as they identified a unique cell population in the region of the sebaceous gland that expressed the transcription factor BLIMP1 and exhibited a unipotent differentiation potential into sebocytes. Loss of BLIMP1 in HFs resulted in activation of bulge cells, suggesting a possible connection between the bulge and sebaceous gland. In support of this hypothesis, the same study also showed that implanted bulge stem cells could give rise into BLIMP1+ cells. 36
DP and dermal sheath
DP and dermal sheath (DS) are cell populations within the HF that are believed to contain stem cells. Whereas bulge cells originate from ectoderm, DP and DS cells are derived from mesoderm and they are known to regulate hair cycling by exchanging signals with the bulge. 2 Multiple studies showed that DP and DS cells have a broad differentiation potential. In a pioneering study, Lako et al. demonstrated that DP and DS cells could reconstitute multiple lineages of the hematopoietic system in lethally irradiated mice. 37 A more recent study showed that DP/DS stem cells are the precursors of dermal stem cells and contribute to dermal maintenance and wound healing. 38 Several laboratories, including ours, showed that rat and human HF-derived DP and DS cells could also be induced to differentiate toward the myogenic, osteogenic, chondrogenic, and adipogenic lineage resembling bone marrow MSCs.3–5,39,40 Notably, we showed that single human DP/DS-derived cell can differentiate into all mesenchymal lineages with high efficiency, suggesting these are truly multipotent stem cells and not a collection of unipotent progenitors. 5
The HF stem cell niche
The interaction of HF-SCs with the microenvironment is critical for the maintenance, expansion, and differentiation of the HF stem cell pool. In contrast to the other stem cell population located in the HF, the microenvironment of bulge-derived stem cells is extensively studied because of its significance in regulating hair cycling. Stem cells located in the bulge are surrounded by cell populations derived from diverge developmental origins, for example, follicular epithelium-ectoderm, melanocyte stem cell-neural crest, DP/DS-mesoderm that all together interact, coordinate and exchange signals to generate a pigmented hair shaft.
Several studies have been demonstrated that the Wnt pathway plays a critical role in HF development, the activation of the bulge niche and consequently the telogen to anagen transition. Activation of the canonical Wnt pathway results in stabilization of beta-catenin by inhibiting its phosphorylation and subsequently degradation. Stabilized beta-catenin accumulates in the nucleus where it makes a complex with TCF/LEF1 and regulates transcription. Indeed, transgenic mice expressing the WNT inhibitor DKK1 under the Krt14 promoter were hairless, as DKK1 inhibited the onset of HF development. 41 In addition, ectopic expression of a truncated stabilized form of beta-catenin by an epidermal promoter resulted in the formation of new HFs and tumors in adult mice. 42 In agreement, transient activation of beta-catenin was sufficient to activate entry of telogen HF into the anagen phase and induce de novo hair formation.43,44
Interestingly, beta-catenin is important in mediating the mesenchymal–epithelial interactions between the DP and bulge cells, as ablation of beta-catenin in the DP reduced proliferation of the adjacent matrix cells and subsequent premature entry into catagen. 45 Interestingly, the nuclear beta-catenin 22 and W/s (a gene responsible for Wnt secretion) appear first in hair germ during the early stages of anagen, suggesting that secretion of the WNT ligand by the follicular epithelium precedes its expression by the follicular mesenchyme. Indeed, deletion of Wntless in the follicular epithelium led to decreased nuclear beta-catenin in both the epithelial and mesenchymal compartments and ultimately, hair cycle arrest. 46
In addition to the WNTs, the TGFB family also regulates stem cell activation and HF cycling. Bone morphogenetic proteins (BMPs) are secreted from both the dermal 47 and bulge cells 23 during telogen and maintain the quiescence of bulge stem cells. Indeed, conditional ablation of the bone morphogenetic protein receptor 1α (Bmpr1a) gene resulted in beta-catenin stabilization and and stem cell activation; conversely, overexpression of Bmpr1a induced differentiation of HF-SCs. 48 In agreement with the inhibitory role of BMP in hair cycling, NOG (NOGGIN) (BMP inhibitor) expressed by the mesenchymal cells of the follicle, was reported to promote hair induction.49,50 Whereas BMP signaling is mediated through SMAD1/5/8, TGFBs signals through SMAD2/3, suggesting a possible differential role in hair cycling. Indeed, a recent study reported that TGFB2 (secreted by DP) decreased the activity of BMP signaling during telogen and this effect was mediated by the TGFB target TMEFF1. 51 Collectively, these studies suggest that there is a fine balance within the bulge niche between stimulatory and inhibitory signals that regulate HF-SC quiescence/activation and hair cycling.
Putative HF Stem Cell Markers
The majority of the studies in the HF have been conducted in murine models. However, there are several differences that have to be taken into account between human and murine models. Whereas humans have only two types of hair (vellus and heavily pigmented hairs), mice are endowed with several distinct hair types (pelage, vibrissae, cilia, hairs on the tail, ear, genital, perianal area, nipples, and around the feet). In addition, the biological cycles of human and mouse HF are different; while the human HFs cycle independently after birth, to a large extent the mouse HFs (e.g., pelage hair) cycle in synchrony. 2 Finally, the biological markers characterizing the stem cell populations in human and mouse are strikingly different. Table 1 summarizes the most common markers of HF stem cells in mice and humans.
Murine HFs
Bulge
Several markers have been proposed to characterize murine bulge stem cells. In addition to KRT15 and LGR5, CD34 is coexpressed with KRT15 and has been proposed as a potential bulge stem cell marker. CD34+ cells are relatively quiescent and have a higher clonogenic potential in vitro as compared to CD34− cells. 52 Several transcription factors have been identified in the bulge region, including TCF3, SOX9, LHX2, and NFATC1. TCF3 was shown to maintain the undifferentiated cell state by repressing numerous genes that induce the sebaceous gland and HF differentiation. 53 Another key transcription factor that is expressed in the bulge area is SOX-9. SOX-9+ cells are first detected during the formation of hair placode (the precursor of HF during prenatal life) and colocalize with early label-retaining cells, which subsequently give rise to bulge stem cells. Notably, deletion of SOX9 decreased the proliferation of bulge stem cells, impaired the generation of proliferative matrix cells, and resulted in inhibition of HF morphogenesis.54,55
Similar to SOX9, the Lim-homeodomain transcription factor, LHX2 is also expressed during hair placode formation as was seen by microarray analysis in the P-cadherin+ cells that mark early hair progenitors. In postnatal life, LHX2 is expressed in the bulge and suppresses differentiation, prompting some investigators to hypothesize that it may be required for stem cell maintenance. 56 However, a recent study challenged this notion and reported that LHX2 is required for the induction of anagen and not for stem cell maintenance. 57 Another bulge-specific transcription factor is NFATC1, which is regulated by the intracellular levels of calcium. Under high calcium conditions NFATC1 is dephosphorylated and translocates to the nucleus, where it downregulates cyclin-dependent kinase 4 and suppresses proliferation in the bulge. As a result, downregulation of NFATC1 leads to activation of bulge-derived stem cells. 58 Interestingly, NFATC1 expressing cells coincide only partially with CD34+, TCF3+, LHX2+, and SOX9+ cells in the bulge region, suggesting there is no unique marker of bulge stem cells, but rather a group of transcription factors that regulate stem cell maintenance and activation through a series of complex and dynamic interactions. In fact, as discussed above, the bulge harbors multiple types of stem cells (slow cycling, actively proliferating, and melanocytes), suggesting that the seemingly intricate distribution of various stem cell markers might reflect the cellular complexity of this stem cell niche.
Finally, other studies provided evidence that Nestin (NES) is expressed in mouse bulge stem cells. Transgenic mice expressing GFP under the Nes promoter showed that NES-positive cells were located in the bulge region during telogen, but in the upper two thirds of the outer root sheath during anagen. In vivo these cells participated in the formation of new blood vessels and in vitro they could be coaxed to differentiate into neurons, glial cells, smooth muscle cells (SMCs), melanocytes, and keratinocytes, demonstrating the multipotency of HF stem cells.24,25,59
Upper bulge
Several markers have been identified over the years that target putative murine stem cells in the upper bulge region. LGR6, an orphan G protein-coupled receptor, is expressed in the region immediately above the bulge. LGR6+ cells were shown to play a critical role in the formation of the HF, sebaceous gland, and epidermis during development. 35 MTS24, a cell surface glycoprotein also marked potential stem cells in a region above the bulge. MTS24+ cells exhibited an increased colony-forming capacity as compared to MTS24− cells and showed a similar gene expression profile with CD34+ bulge cells. 32 However, the differentiation potential of these cells was not examined. Another putative stem cell marker characterizing the region right above the bulge is the transmembrane protein Leucine-rich repeats and immunoglobulin-like domain protein 1 or LRIG1. LRIG1 was shown to regulate epidermal growth factor signaling by promoting the degradation of the epidermal growth factor receptor 60 and to keep cells in this region in a quiescent state. 61 Indeed, in vivo LRIG1+ cells appeared to be quiescent and multipotent, two of the main attributes of stem cells. 34 Finally, cells residing in the upper isthmus were shown to be multipotent as they could form the HF, sebaceous gland, and epidermis after implantation. These cells expressed low levels of integrin alph6 (ITGA6), were negative for the hematopoietic markers CD34 and SCA1, and exhibited distinct gene expression profile as compared to bulge cells. 33
DP and DS
The DP and DS is known to induce HF generation by interacting with epidermal stem cells. 62 In 1999, Kishimoto et al., reported that cells in DP express the proteoglycan versican, which is usually present in the condensed mesenchyme. The same group employed the versican promoter to express either lacZ or GFP and found that when implanted on the back of nude mice along with keratinocytes, the versican (VCAN)+ cells could reconstitute the HF, but VCAN− cells could not. 63 Others observed that NEXIN1, a protease inhibitor, was highly expressed in DP during anagen and that the NEXIN1 expression level correlated with the rate of hair growth. 64 Similarly, the expression of alkaline phosphatase (ALPL)—an enzyme expressed in bone cells and ESCs—correlated with hair growth and was also highly expressed in DP during anagen, suggesting a positive correlation between hair induction and ALPL activity. 65 Finally, CD133 was expressed in DP cells during the HF development, but after birth it was expressed only transiently during anagen. Nevertheless, when coimplanted with embryonic epidermal cells, CD133+ cells enabled generation of HFs in vivo. 66 Interestingly, a subpopulation of CD133+SOX2+ cells within the DP was shown to be essential for the formation of particular types of hair, such as awl/auchene follicles. 67 Rendl et al. compared the transcriptional profile of five distinct cell populations within the HF, namely, melanocytes, DP, matrix, outer root sheath, and dermal fibroblasts. This approach successfully identified several genes and signaling pathways that were unique to each population and need to be further explored in the future. 68
Human HFs
Murine HFs have been largely explored with respect to stem cell markers, however, human HFs remain relatively unexplored. In contrast to the murine bulge, the human bulge cannot be identified as a distinct anatomic projection, rendering isolation of bulge cells very challenging. Screening a number of markers in vivo, Kloepper et al. identified CD200, KRT15 and KRT19 as putative bulge stem cells markers, although their location is not restricted to the bulge, but extends to a wider area of isthmus as well. In contrast to mouse, human bulge cells do not express CD34, NES, or LHX2. 69 CD200 expressing cells that were isolated from a population of label-retaining cells using laser capture microdissection showed an increased clonogenic potential in vitro, but the multipotency of CD200+ cells was not examined. 70 On the other hand, KRT15high/CD200+/CD34−/CD271− bulge-derived cells showed an increased clonogenic potential as compared to KRT15low/CD200+/CD34−/CD271− cells. 71 More recently, our laboratory reported that human DP/DS cells displayed a cell surface profile characteristic of MSCs being positive for CD90, CD44, CD49b, CD105, and CD73.4,5 These cells were clonally multipotent as individual clones could be coaxed to differentiate into fat, bone, cartilage, and smooth muscle with high efficiency. 5
Extensive studies on murine models provided a better understanding of the HF biology and identified many putative stem cell markers. Unfortunately, very few studies on human HFs exist and a reliable marker is still under quest. However, due to the apparent differences between human and mice, the conclusions derived from experiments with mice do not necessarily apply in the human HF. Therefore, further studies have to be conducted on human HFs to verify the results. In this direction, engineering-reliable in vitro models that can recapitulate the in vivo environment of the human HF are absolutely necessary. The most prevalent approach has been the isolation and culture of intact, anagen HFs. Using this model, the effect of several growth factors and cytokines on the HF growth and morphology have been examined.72–76 Although the culture of intact follicles is a well-established system, still many issues need to be resolved so that it can better mimic the in vivo environment. The main challenges facing this model are the absence of hair cycling and consequently, the limited culture time in vitro (9 days).
On the other hand, engineering the HF from its constituent cellular components that is, epithelial stem cells and dermal stem cells in the presence of the appropriate functional scaffold—might provide the appropriate microenvironment to support cycling and sustain longer culture times. To this end, Krugluger et al. injected human DP and outer root sheath cells into an in vitro skin organ culture model and observed induction of velus-like HFs, demonstrating that engineering HFs in vitro is feasible. 77 Recent studies showed that biomaterials can be decorated with various biomolecules to provide appropriate growth or differentiation signals as necessary.78–83 It might be interesting to examine whether this approach could be applied in the context of HFs to overcome the current limitations and facilitate studies of human HF biology.
Methods for Isolating HF Stem Cells
Three techniques have been routinely used for the isolation of putative stem cells from the HF: microdissection, enzymatic digestion, and fluorescence-activated cell sorting (FACS). In the following, we describe each technique and elaborate on their advantages and disadvantages.
Microdissection
Microdissection is a technique that has been commonly applied for the isolation of cells from DP11,37,39,84,85 or the bulge.70,86 This technique requires use of fine forceps and blades for the isolation of the area of interest, which is subsequently transferred onto tissue culture plates, where the cells migrate out of the tissue and proliferate in the presence of an appropriate culture medium. For DP cell isolation, application of pressure on the suprabulbar region by forceps was shown to compress the bulb and facilitate removal of the connective tissue sheath surrounding the DP, which is subsequently detached from the epithelium using a scalpel blade. 87 Finally, a highly reliable technique that has been used for isolating human bulge cells is laser capture microdissection. 70 A thermolabile membrane is placed on top of the sample and the area of interest is targeted by laser, which melts the membrane locally marking the cells that are subsequently separated. 88 The major advantage of microdissection is that it preserves the whole tissue, thereby increasing the efficiency of cell isolation. However, it is quite laborious and requires experienced technicians.
Enzymatic digestion
Another approach that has been employed for isolation of HF stem cells involves enzymatic digestion of the follicle from the surrounding dermis, usually with dispase or collagenase. The incubation time and concentration of enzymes used vary depending on the amount of extracellular matrix present around the follicle. Generally, collagenase treatment requires few hours of incubation at 37°C, whereas dispase requires overnight treatment.3–5,27,70 Others use a combination of enzymes to isolate DP cells. Specifically, dispase was employed initially to remove follicles from the cutaneous fat, followed by collagenase D to digest the DS and isolate the DP. The remaining DS fibroblasts could be removed by low-speed centrifugation of the DP. 89 Enzymatic digestion is a simple method of HF stem cells isolation, but with little control over the type of cells that are obtained leading to possible variation between isolations.
Fluorescence-activated cell sorting
FACS is a common method for isolating stem cells, especially from the murine HF.15,16,24,33,35,63,66,67 Fluorescently labeled antibodies are used to tag the cell surface and cells are sorted based on the fluorescence intensity, which is proportional to the expression level of the particular target receptor. FACS can also be applied for isolating cells based on markers that are not expressed on the cell surface. Our group made use of the smooth muscle alpha-actin (ACTA2) promoter-driven GFP to isolate a homogeneous population of SMCs from ovine and human HF-MSCs.3,4 FACS yields highly purified cell populations that can be further expanded or directly analyzed for mRNA or protein expression. Regrettably, lack of reliable stem cell markers hampers use of this method in sorting human HF stem cells.
HF Stem Cells for Tissue Engineering and Cell Therapy
The emergence of the tissue engineering and regenerative medicine fields led to advances in the development of novel strategies or restore or replace lost tissue and organ function. Although technological advances toward engineering of suitable biomaterials have been made, obtaining large number of cells with enhanced therapeutic properties as required for regenerative medicine remains a major challenge. On the other hand, stem cells are promising cell sources that have increased proliferation and broad differentiation capacity, making them suitable for tissue engineering applications. Although ESCs and induced pluripotent stem cells are great potential sources due to their unlimited proliferation capacity and wide differentiation potential, ethical, immunogenic, and tumorigenic concerns prevent their clinical application at the moment. In contrast to ESCs, adult stem cells, such as MSCs, are more accessible, nontumorigenic, autologous, and have been used successfully in the clinic. 90 Regrettably, the properties of adult stem cells are greatly affected by donor aging91–93 and culture-induced senescence,94–96 leading to loss of proliferation capacity and multipotency. This is of particular clinical interest since the majority of the patients that are in need for cellular therapies are elderly.
In the present review, we propose HF stem cells as an alternative cell source for tissue engineering and cellular therapies. HFs are easily accessible as they are abundant in human skin and can be isolated relatively easily as compared to other stem cell populations, such as bone marrow (BM)-MSCs. In addition, HF-SCs are highly proliferative in vitro4,5,32,52 and it was estimated that a single human HF could give rise to ∼1015 MSCs, even when originating from elderly donors (>65 years old).4,5 This number is considerably higher than the reported values for human BM-MSCs that could be expanded to ∼1010 cells from individual aspirates.94,95,97 Given that the number of HFs in the human adult scalp between 175–300 hairs/cm2, 98 the HF appears to be a very rich source of stem cells.
In addition, HF-SCs have broad differentiation potential. Bulge stem cells not only could they generate intact HFs if mixed with dermal fibroblasts, but they also exhibited the potential to differentiate into keratinocytes, neurons, melanocytes, and mesenchymal cells.18,19,24–28 In addition, the DP/DS-derived stem cells could be coaxed to differentiate into functional SMCs, adipocytes, osteocytes, chondrocytes, hematopoietic cells, and dermal fibroblasts.3–5,37–40 Notably, anagen HFs downregulated the major histocompatibility complexes I and II molecules and produced immunosuppressants, suggesting that they might be nonimmunogenic. 99 Although these observations pertained to the epithelial portion of the HF, transplantation of human DP also evaded immune rejection. 100 Therefore, the potential of HF-SCs—epithelial stem cells or DP/DS-derived MSCs—for use in tissue engineering and regenerative medicine is high and some of the potential applications are discussed below.
Tissue-engineered vascular grafts
A functional arterial graft should contain both endothelial cells (ECs) and SMCs. ECs line the lumen of a vessel, endow it with thromboresistant properties, and are selectively permeable to substances circulating in the blood. SMCs form the medial layer of an artery and are mainly responsible for the dilatation and constriction of the vascular wall in response to vasoactive agonists. Our laboratory showed that DS cells of ovine and human HF stained positive for ACTA2, a marker of SMC (Fig. 2). This finding prompted us to hypothesize that functional SMCs can be derived from the HF. To this end, HFs were transduced with a lentivirus encoding for GFP under the control of the ACTA2 promoter and GFP+ cells were sorted out using flow cytometry. We found that both ovine and human HF-derived SMCs exhibited a significantly higher proliferation and clonogenic potential compared to vascular SMCs. In addition, tissue-engineered vascular grafts prepared from HF-derived SMCs displayed high reactivity in response to vasoactive agonists and generated significant mechanical force as shown by compaction of fibrin hydrogels.3–5,101 More recent studies in our laboratory showed that these vascular grafts could be implanted into the arterial circulation of an ovine animal model where they remained patent for at least 3 months 102 (and unpublished data), suggesting that the HF may be a readily accessible source of stem cells for cardiovascular tissue regeneration and cell therapies.
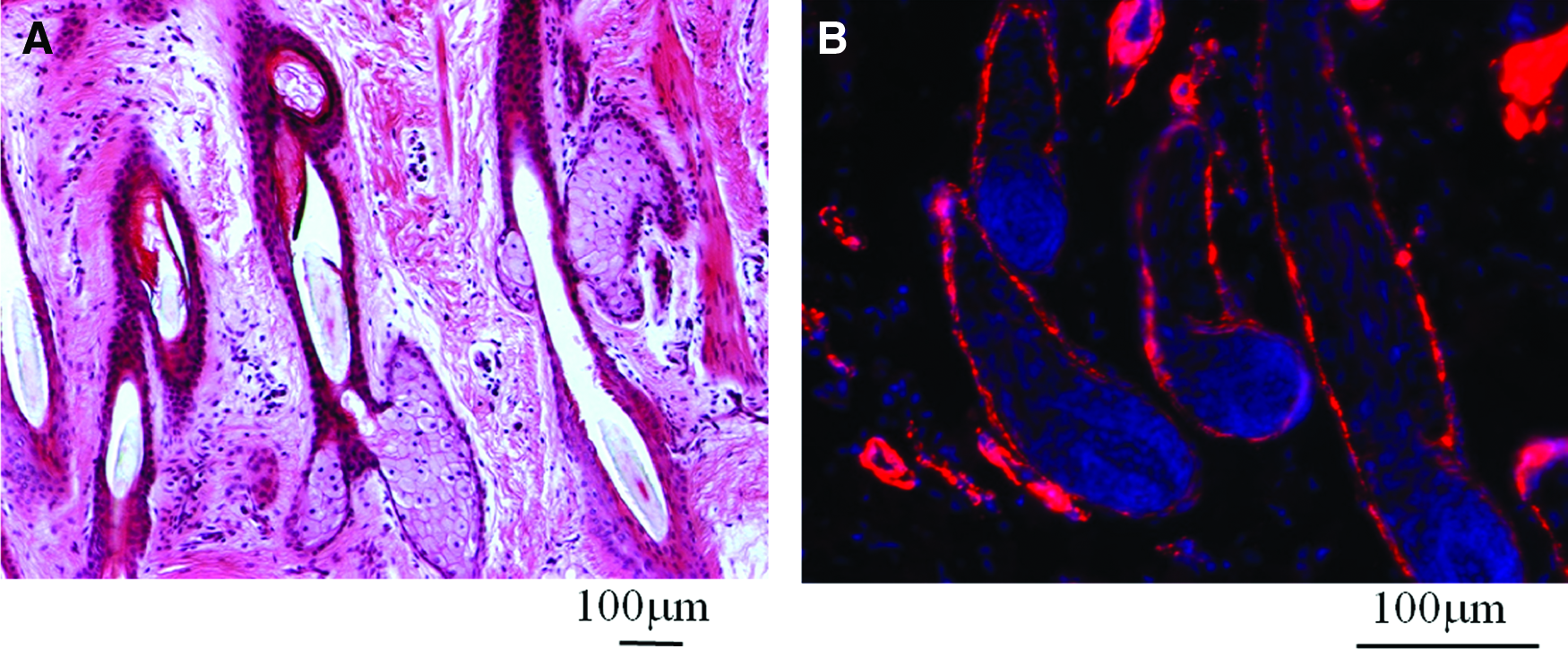
Cells comprising the DS of hair follicle are positive for smooth muscle alpha-actin (ACTA2).
Tissue engineering of cartilage, bone, and fat
In addition to myogenic differentiation, rodent DP/DS cells have the capacity to differentiate into the osteogenic, chondrogenic, and adipogenic lineage, similar to bone marrow-derived MSCs.39,40 Extending these studies, we demonstrated that human DS/DP cells also possess a multilineage differentiation potential. 4 We also showed that single clones gave rise to all four lineages, strongly indicating that human HF cells represent true mesenchymal stem cells—termed HF-MSC—and not a mixed population of unipotent progenitors. 5 These results suggest that the human HF can be an easily accessible source of true MSCs that could be employed for regeneration of bone and cartilage for the replacement of joints or for meniscus repair.
Skin regeneration
Several studies suggested that HF cells migrate to the epidermis during homeostasis and to a larger extent following skin injury,29,30,35,38,103 suggesting that HF cells could be used to generate the epidermis and enhance wound healing. Indeed, Hoeller et al. reported generation of bioengineered skin by introducing fibroblasts and HF tissue into the dermis. Interestingly, epidermal keratinocytes migrated out of the HF and developed multiple layers of epidermis and stratum corneum. 104 In addition, HF-derived melanocytes have been used to develop a pigmented skin substitute. 105 Most important, transplantation of tissue-engineered skin from HF-derived stem cells was shown to enhance healing of ulcers and burns significantly.106–108 Notably, when hair buds were introduced into bioengineered skin before implantation, they sped up and guided nerve regeneration, suggesting that the HF may recover the lost sense of touch. 109
Nerve regeneration
Mouse HF-derived NES+/KRT15− stem cells have the capacity to differentiate into neurons in vitro suggesting a possible application to nerve regeneration in a variety of central and peripheral nervous system diseases. 26 Indeed, Amoh et al. transplanted mouse HF Nestin+ stem cells into a severed sciatic nerve or spinal cord, where they differentiated into Schwann cells and promoted nerve regeneration.59,110 The same group also reported that human HF stem cells have the capacity to restore the function of injured nerves.111,112 HF-derived neuronal and Schwann cells have also been introduced into an acellular sciatic nerve conduit, where they exhibited long-term survival and significant electrophysiological properties in vitro, but failed to induce repeated potentials. 113
Engineering functional HF
An important application of HF stem cells is the bioengineering of HFs to restore hair loss (alopecia). Common forms of alopecias include (1) the androgenetic alopecia, which results from the miniaturization of the hair; (2) the alopecia areata, which results from an autoimmune response that damages the HF; and (3) permanent alopecia, which can be caused, for example, from severe trauma. 1
Bioengineering HF has been a topic of intense scientific research over many years. To date, two strategies have been developed to achieve this goal. The first approach includes the transplantation of intact HFs from a HF-rich area into the bald area. This technique requires initially the surgical excision of a thin strip of scalp that contains dense HFs and subsequently the isolation of the individual follicles and implantation to the hairless scalp. 114 Although transplantation of whole follicles is considered as the gold standard for hair restoration, studies demonstrated that segments of the HF can also induce hair growth after transplantation.85,100,115–119 Transplantation of a truncated human HF after amputating the bulb resulted in hair regeneration, suggesting that the bulb compartment can be regenerated from the DS compartment.115,116,119 Interestingly, transplantation of intact DS into mouse85,117 or human dermal compartments 118 led to hair induction, possibly as a result of interactions between mesenchymal cells with the host epithelium. However, in contrast to human DS when human DP was transplanted into human skin, it failed to induce hair regeneration. 100
In severe cases of alopecias, the number of available HFs is not sufficient to restore the bald site. On the other hand, HF stem cells can be expanded in culture into large numbers that may be sufficient to cover the whole area and result in hair restoration. Jahoda et al. were the first to report that implantation of DP cells resulted in hair growth in mice. 62 Although the hair-inductive properties of DP cells were lost after long-term expansion in vitro, coculture with keratinocytes or in the keratinocyte conditioned medium could maintain the inductive properties of DP cells for almost 70 passages. 120 Similar to DP, DS cells were also found to induce HF growth. 121 Finally, HF restoration was enhanced by the mixture of bulge/hair germ stem cells from adult HFs with neonatal dermal cells.15,16,18,34,35 Notably, when mixed with embryonic mouse dermal and epidermal cells, mouse bone marrow-derived cells differentiated into HF cells, suggesting hair-inductive properties of bone marrow cells. 122 Although the results with mouse models are very encouraging, the significance of these findings in large animal models or humans has yet to be demonstrated.
Drug delivery through the HF
Skin is an easily accessible organ that has been widely considered as a unique target for drug delivery. In contrast to the conventional delivery methods (oral, injections), the transdermal route allows drug administration to the circulation through the dermal vasculature and may increase drug bioavailability while avoiding painful injections. However, the presence of stratum corneum, the outermost layer of the skin, severely limits the penetration of hydrophilic and high molecular weight substances. 123 To bypass this drawback, microscale devices have been developed to enable transdermal delivery, including liquid jet injectors, microneedles, and thermal ablation devices. 123
Alternatively, scientists have focused on drug administration via the follicular route. The HF disrupts the stratum corneum and provides an opening to the epidermis. In certain areas, such as the scalp or the face, the total area of pores can reach up to 10% of the skin area, contributing significantly to solute permeation.124–126 In addition, the dense network of blood vessels that are associated with the HF suggests that drug release to the circulation may be feasible. 127 The heterogeneity of the HF resident cells (stem cells, gland cells, immune cells, etc.) may enable cell-specific drug targeting for treatment of skin diseases or vaccination.128–130 Last, but not least, the relatively large volume of infundibulum renders the HF a reservoir for sustained drug release to the circulation, further highlighting the importance of follicular delivery. 127
Several studies highlighted the contribution of follicular penetration during drug delivery through the skin. Mitragotri modeled the permeability of hydrophilic and hydrophobic compounds in skin, assuming that the solutes can transport through one or more of the following mechanisms: free-volume diffusion, lateral diffusion of the lipids, diffusion through pores, or diffusion through shunts (HFs and glands). The model predicted that high molecular weight and highly hydrophilic molecules penetrate the skin preferentially through the shunts. 131 Others suggested that there is a critical value of octanol/water partition coefficient beyond which the flux through the follicle is greatly diminished. 132 However, most studies omit the significance of sebum (a lipophilic product of the sebaceous gland) during drug delivery due to lack of representative experimental models. The presence of sebum in the HF and its upward flow may hinder the delivery of hydrophilic compounds and may favor the delivery of hydrophobic compounds. Indeed, apart from molecular weight and molecular orientation, diffusion through the sebum was found to be affected by compound lipophilicity. 133
To further improve tissue targeting and drug delivery via the HF, studies have incorporated particle-based formulations. Lademann et al. demonstrated that nanoparticle containing dye could penetrate up to 1400 μm into the follicle of porcine skin, whereas the nonparticle formulation reached only 500 μm. Interestingly, the nanoparticles prolonged the storage of the dye into the follicle. 134 The nanoparticle size was shown to play a critical role in follicular penetration, which was optimal for particles between 750–1500 nm and decreased for larger particles. 135 In addition, Vogt et al. demonstrated that the particle size affected cellular uptake. They reported that only the 40-nm size nanoparticles could enter Langerhans cells that were localized around the HF, indicating that size-specific particle formulations can be designed to target antigen-presenting cells and deliver vaccines via the follicular route. 129 Finally, systemic delivery of a chemical through the HF has also been examined in vivo.127,136 Caffeine was introduced into a shampoo formulation and its delivery into the circulation via the skin was examined in human subjects. Interestingly, the follicular route not only accelerated the delivery, but it also prolonged detection of caffeine in the blood indicating that HFs may act as a reservoir of chemical compounds.
Cell and gene therapy using HF stem cells
The goal of gene therapy is to restore the lost tissue function by introducing the correct gene copy at the sites where the gene is missing or mutated. 137 Application of gene therapy for hair restoration has been attempted and showed promising results. Transduction of rat bulge-derived HF stem cells with lacZ-encoding retrovirus showed stable expression of the transgene in the HF epithelial compartments for at least 6 months after implantation of transduced cells in an immunodeficient mouse model. 138 Retroviral gene transfer of the streptomyces tyrosinase gene was used to treat albinism. Specifically, transduction of ex vivo cultured skin from albino mice restored melanin production from the skin HF. 139 Direct gene transfer into the skin in vivo has also been reported to restore hair growth. Intradermal administration of the Sonic Hedgehog gene into C57BL/6 mice using an adenovirus resulted into anagen induction and subsequently enhanced hair growth. 140 More recently, in vivo transfection of the human telomerase reverse transcriptase DNA complexed with polyethylenimine induced telogen to anagen transition in the rat dorsal skin. 141 In addition to the treatment of hair- or skin-related disorders, gene transfer to the HF could be used for delivery of proteins into the systemic circulation through the vascular plexus surrounding the follicles. As we demonstrated previously, human bioengineered skin could be engineered to produce insulin in a regulatable manner by administration of a small molecule (doxycycline), reversing hyperglycemia in a diabetic mouse model. 142 Engineering HFs into the dermal compartment of bioengineered skin might increase insulin production and facilitate transport to the system circulation further enhancing the potential of this technology.
Reprogramming of HF stem cells
In a breakthrough study in 2006, Yamanaka and colleagues demonstrated that introduction of four transcription factors (OCT4, SOX2, KLF4, and MYC) into mouse embryonic fibroblasts or adult fibroblasts endowed them with enhanced proliferation capacity and potential for differentiation into all three germ layers, similar to ESCs.143–146 The Thomson group demonstrated that two of the transcription factors (KLF4 and MYC) could be replaced by NANOG and LIN28 with a similar outcome. 147 The resulting cells were designated as induced pluripotent stem cells (iPSCs). An explosion of studies that followed demonstrated that iPSCs could be generated from many human cells, including blood cells,148,149 MSCs, 150 fetal 150 and neonatal fibroblasts,147,150 adipose-derived stem cells, 151 adult testis, 152 β-pancreatic cells, 153 and T lymphocytes. 154 Interestingly, HF-derived primary keratinocytes could be reprogrammed with 100-fold higher efficiency than fibroblasts. 155 Finally, DP cells were shown to reprogram using only two factors (OCT4, KLF4), 156 suggesting the presence of endogenous factors that might facilitate reprogramming.
A higher reprogramming efficiency with fewer transcription factors suggests that HF cell-derived iPSCs may be useful for regenerative medicine applications as well as for development of models to study the genetics and pathophysiology of human disease. Indeed, HF-derived MSCs were also reprogrammed and used to understand the feedback loops that sustain self-renewal using global genomic and proteomic strategies. 157 In our laboratory, we developed a robust, stage-wise strategy to differentiate human HF-MSC-derived iPSCs (HF-iPSCs) into functional SMCs through an intermediate stage of multipotent and highly proliferative MSC that could be propagated for more that 45 population doubling. Interestingly, vascular constructs engineered from HF-iPSC-derived SMCs exhibited high levels of contractility in response to receptor- and nonreceptor-mediated agonists. 158 These results are encouraging and suggest that our strategy may facilitate acquisition of large numbers of cells required for regenerative medicine or for engineering vascular tissues that could serve as models for studying mechanisms of vascular development or disease.
Conclusions and Future Directions
In summary, HF stem cells have a great potential for tissue engineering and regenerative medicine applications. The ease of accessibility along with the broad differentiation capacity of HF stem cells makes the HF an ideal stem cell source. However, human HF stem cells remain relatively unexplored as compared to their mouse counterparts or other human adult stem cells. As a result, more studies are required to address a number of challenges that hinder application of these cells in regenerative medicine. To this end, identification of reliable HF stem cell markers is urgently needed to facilitate HF stem cell isolation. More studies are also needed to evaluate the differentiation potential of human HF stem cells and establish culture conditions for efficient differentiation. The ease of reprogramming should be further explored to identify potential small molecules that may induce reprogramming even in the absence of genetic modification. 159 Finally, more studies are necessary to establish the HF as a site for drug and gene/protein delivery, for treatment of skin diseases and wound healing or to the blood circulation for treatment of systemic disorders.
Footnotes
Acknowledgments
This work was supported by grants from the National Institutes of Health (R01 HL086582) and the New York State Stem Cell Science (NYSTEM Contract #C024316) to S.T. Andreadis. The authors would also like to thank Anna Mistriotis for her help with the schematic representation of the HF.
Disclosure Statement
No competing financial interests exist.