
Abstract
Mesenchymal stem cells (MSCs) are multipotent cells that can differentiate into various cell types and have been widely used in tissue engineering application. In tissue engineering, a scaffold, MSCs and growth factors are used as essential components and their interactions have been regarded to be important for regeneration of tissues. A critical problem for MSCs in tissue engineering is their low survival ability and functionality. Most MSCs are going to be apoptotic after transplantation. Therefore, increasing MSC survival ability and functionalities is the key for potential applications of MSCs. Several approaches have been studied to increase MSC tissue forming capacity including application of growth factors, overexpression of stem cell regulatory genes, and improvement of biomaterials for scaffolds. The effects of these approaches on MSCs have been associated with activation of the phosphoinositide 3-kinase (PI3K)/Akt signaling pathway. The pathway plays central regulatory roles in MSC survival, proliferation, migration, angiogenesis, cytokine production, and differentiation. In this review, we summarize and discuss the literatures related to the roles of the PI3K/Akt pathway in the functionalities of MSCs and the involvement of the pathway in biomaterials-increased MSC functionalities. Biomaterials have been modified in their properties and surface structure and loaded with growth factors to increase MSC functionalities. Several studies demonstrated that the biomaterials-increased MSC functionalities are mediated by the activation of the PI3K/Akt pathway.
Introduction
Although MSCs were discovered from bone marrow (BMSCs), they have also been isolated from other tissues, including adipose tissue, umbilical cord blood, placenta, dental pulp, and pancreatic islets.32–36 All these MSCs have similar molecular markers with positive expression of mesenchymal markers (CD90, CD29, CD44, and CD105) and lack of CD34, CD45, CD11b, and HLA-DR.36,37 MSCs from adipose tissue (ASCs) are extensively studied to substitute BMSCs with several obvious advantages. First, the adipose tissues are easier to obtain than bone marrow. Second, MSCs in adipose tissue are much more abundant than that in bone marrow. Third, ASCs have higher ability to proliferate than BMSCs. 37
A major problem in the application of MSCs is that the differentiation capability of MSCs is decreased in the transplanted environment. 38 This decrease could be due to the new extracellular microenvironment, which is different from the natural environment of MSCs and is hostile to these cells. Therefore, most MSCs cannot survive long enough to generate terminally differentiated cells after transplantation. 38 For example, 90% of MSCs were shown to be apoptotic, a programmed cell death, within 3 days after transplantation into heart for cardiomyocyte infarct repair.39,40 However, cellular therapy using MSCs is still effective to a certain extent. The beneficial effects of MSCs are thought to be mediated by cytokines produced by MSCs rather than new tissues formed from MSCs.39,41 This may also be the case in tissue engineering. Therefore, there is room to further improve the efficacy of MSC-based treatments by increasing the survival ability and functionality of MSCs in the transplantation environment.
Many different approaches have been used to stimulate MSC proliferation and differentiation and to promote long-term survival and functionalities. These approaches include the delivery of bioactive materials or proteins, 42 the overexpression of genes in MSCs such as vascular endothelial growth factor (VEGF), Protein kinase B (Akt), bcl-2,43–48 and the improvement of biomaterials used for scaffolds. 49 Interestingly, all above approaches are involved in an intracellular signal pathway, phosphoinositide 3-kinase/Akt (PI3K/Akt). In this review, we will summarize the roles of the PI3K/Akt pathway in the functionalities of MSCs, how biomaterials affect MSC functionalities via the activation of the PI3K/Akt pathway, and discuss possible implications.
The PI3K/Akt Pathway in MSCs
Overview of the PI3K/Akt pathway
The PI3K/Akt pathway is a survival pathway that regulates cell proliferation, apoptosis, differentiation, and migration. 50 The upstream stimulators of the pathway include various growth hormones, cytokines, and foreign molecules. The PI3K/Akt pathway has been shown to play key roles in the physiology and pathophysiology of many types of cells.51–54 Abnormal increase or decrease in the PI3K/Akt pathway activity is associated with many diseases such as cancer, diabetes, laminopathy, stroke, and neurodegenerative diseases and manipulation of the pathway has been proposed for the treatment of these diseases. 50 The PI3K/Akt also plays a key role in regulating neuronal function through protein ataxin-1, GABA receptor, and disruption of the pathway is associated with diseases such as Huntingtin disease. 55
The key enzyme of the pathway, PI3K, converts phosphatidylinositol 4,5-biphosphate into phosphatidylinositol 3,4,5-triphosphate, which binds both Akt and 3-phosphoinositide-dependent protein kinase 1 (PDK1), allowing PDK1 to phosphorylate Akt.54,56 Class 1A PI3K consists of two subunits: the catalytic subunit p110 and the regulatory subunit p85. There are four isoforms of p110: p110-alpha, beta, gamma, and sigma and three p85 isoforms: p85-alpha, beta, and gamma. The primary direct downstream target protein of PI3K is Akt. There are three Akt isoforms: Akt1, Akt2, and Akt3. 57 The activation of Akt causes a cascade of responses of downstream targets that regulate cellular functions (Fig. 1). For example, Akt regulates cell migration via Rac1 and rhoA, increases cell survival via bcl-2, enhances angiogenesis via VEGF, and increases cell proliferation via the activation of mammalian target of rapamycin (mTOR).50,54
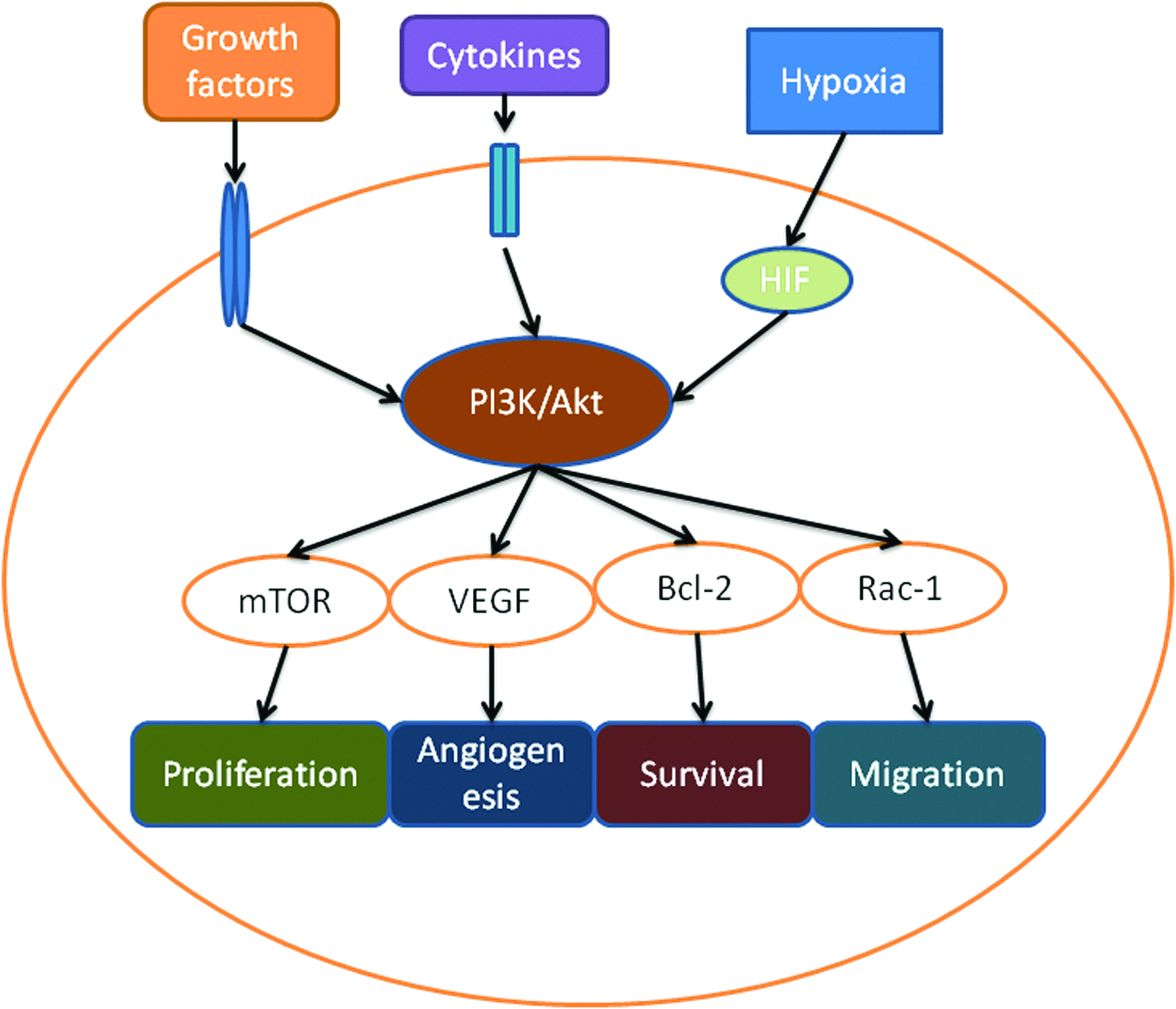
The phosphoinositide 3-kinase (PI3K)/ Protein kinase B (Akt) pathway in mesenchymal stem cells (MSCs). The activity of the PI3K/Akt pathway in MSCs can be stimulated by growth factors or hypoxia. Activated Akt promotes MSC proliferation, angiogenesis, survival, and migration via downstream target proteins such as mammalian target of rapamycin (mTOR), vascular endothelial growth factor (VEGF), Bcl-2, and Rac-1. Color images available online at www.liebertpub.com/teb
The activation of the PI3K/Akt pathway in MSCs has been applied in both cellular therapy and tissue engineering. In cellular therapy studies, stimulators of the pathway and the overexpression of some major components of the pathway have been used to induce the repair of infarcted cardiovascular cells and were demonstrated to increase the efficacy of MSCs.22,58,59 In tissue engineering studies, MSCs have been transplanted together with various growth factors, such as VEGF and fibroblast growth factor (FGF), to increase the functionality of MSCs. 14 However, the overexpression of key components of the PI3K/Akt pathway has not been applied in tissue engineering and warrants further studies.
The PI3K/Akt pathway and survival ability of MSCs
The role of the PI3K/Akt in the survival of MSCs is evidenced by the effects of both overexpression of key components of this pathway and chemical stimulation of the pathway (Fig. 2). Mangi et al. overexpressed Akt1 in MSCs to improve MSC survival after transplantation into the heart in rats. 58 The overexpression of Akt1 greatly improved the ability of MSCs to treat myocardial infarction. In Akt-overexpressing MSCs, the levels of target proteins downstream of Akt, such as anti-apoptotic protein bcl-2, were increased, whereas the level of the pro-apoptotic protein bax was decreased. Lim et al. extended the study to a large animal model, showing that Akt-overexpressing MSCs also exhibited increased efficacy in porcine myocardial infarction. 60
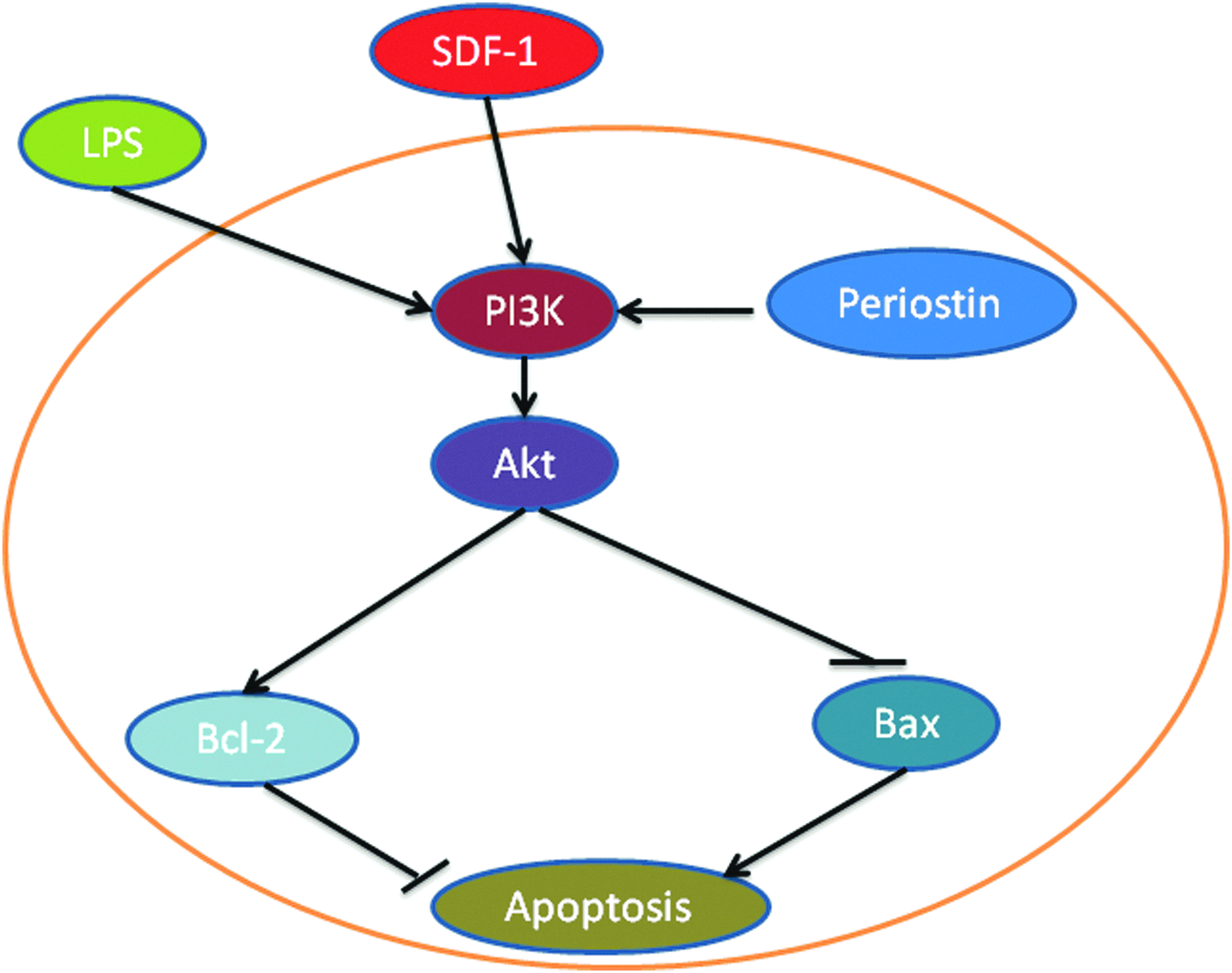
PI3K and the survival of MSCs. In MSCs, the activity of the PI3K/Akt pathway is stimulated by stromal cell-derived factor-1 (SDF-1) and lipospolysaccharide (LPS) and by the overexpression of periostin. Activated Akt can increase MSC survival by blocking bax, which is a pro-apoptotic protein, and by increasing Bcl-2, which can reduce the level of apoptosis. Color images available online at www.liebertpub.com/teb
Periostin is a matricellular protein that can bind to integrin to activate the PI3K/Akt pathway.61,62 It has been overexpressed in MSCs to increase the survival of these cells. 59 In periostin-overexpressing MSCs, the PI3K/Akt pathway was activated, and the therapeutic efficiency was increased. Periostin-overexpressing MSCs and the conditioned medium from these cells were able to increase the survival of cardiomyocytes under hypoxic conditions in a coculture system.
The overexpression of bcl-2 has been shown to increase MSC survival and increase MSC function in the treatment of myocardial infarction. 63 In a rat model of myocardial infarction established by left anterior descending ligation, MSCs overexpressing bcl-2 were injected into damaged rat hearts. These MSCs increased the survival of the transplanted cells in vivo. In an in vitro experiment, bcl-2 reduced MSC apoptosis by 32% and increased VEGF secretion by 60% under hypoxic conditions. 63 The overexpression of bcl-2 in MSCs has also been shown to increase their ability to protect neurons. 64 These gene-overexpressed MSCs may be used in tissue engineering in a scaffold to increase tissue formation.
Pretreatment of MSCs with PI3K/Akt pathway stimulators has been shown to increase MSC survival. Several toll-like receptor stimulators have been used to increase MSC functionality in tissue engineering, and the PI3K/Akt pathway has been demonstrated to play a key role. Wang et al. showed that lipopolysaccharide (LPS), a TLR4 ligand, increased MSC survival. The pretreatment of MSCs with 1 mg/mL of LPS protected MSCs from oxidant stress-induced apoptosis. 65 The activation of the PI3K/Akt pathway was demonstrated by the increased phosphorylation of Akt at Ser 473 and phosphorylatopn of nuclear factor-kappaB (NF-κB) p65 at Ser 536. The effect of LPS was abolished in TLR4-deleted MSCs, indicating that the LPS/TLR4/PI3K pathway played a key role.
Stromal cell-derived factor-1 (SDF-1) has been shown to increase MSC survival by several studies.66–68 Liu et al. demonstrated that SDF-1 can increase the resistance of MSCs to apoptosis induced by H2O2. 39 The pretreatment of MSCs with SDF-1 significantly reduced apoptosis by activating the PI3K/Akt pathway. In these cells, the Bcl-2/bax ratio was increased, indicating a decreased level of apoptosis induced through the mitochondrial pathway. Yin et al. showed that addition of 0.5–2 μg/ml of SDF-1 inhibited hypoxia and serum deprivation-induced apoptosis of MSCs. 66 The effect was abolished by PI3K inhibitor wortmannin, indicating the important role of the PI3K/Akt pathway in SDF-1-increased MSC survival.
The PI3K/Akt pathway and MSC proliferation
MSCs must proliferate to be successfully applied in tissue engineering. First, MSCs must be enriched in vitro through cell proliferation because MSCs are rare in the bone marrow. MSCs only account for 0.001–0.01% of all bone marrow cells.5,22 Second, it could be helpful if MSCs proliferated after transplantation. It has been shown that the actual number of MSCs that form new tissue is far too small. 22 This could be caused by low survival rates and proliferation. Evidence showed that less than 3% of MSCs persist 2 weeks after transplantation.22,39 In in vitro cell culture, many growth factors have been used to stimulate MSC proliferation such as prostaglandin E2 (PGE2), hepatic growth factor (HGF), FGF, tumor necrosis factor-alpha (TNF-alpha), transforming growth factor-beta (TGF-beta), epidermal growth factor (EGF), and insulin.69–71 All these factors are stimulators of the PI3K/Akt pathway, indicating the importance of the activation of the pathway in the proliferation of MSCs.
The activation of the PI3K/Akt pathway has been demonstrated to be a mediator of several factors that increase MSC proliferation. PGE2 has been shown to increase the proliferation of MSCs from human umbilical cord blood. 70 PGE2 phosphorylates Akt, which in turn phosphorylates GSK-3beta, leading to the accumulation of active beta-catenin in the nucleus (Fig. 3). Beta-catenin is a transcriptional factor that increases the expression of c-Myc to promote cell proliferation. 70 TNF-alpha can also increase MSC proliferation via the activation of downstream target proteins of the PI3K/Akt pathway, including NF-κB and cyclin D1.72,73 FGF-beta has been extensively studied to increase MSC proliferation and the activation of the PI3K/Akt pathway was shown to be important for the effect.74,75 The pathway is also involved in EGF-increased MSC proliferation.76,77 These factors may be studied in tissue engineering apparatus for their effects on the proliferation of MSCs and on the activity of the PI3K/Akt pathway.
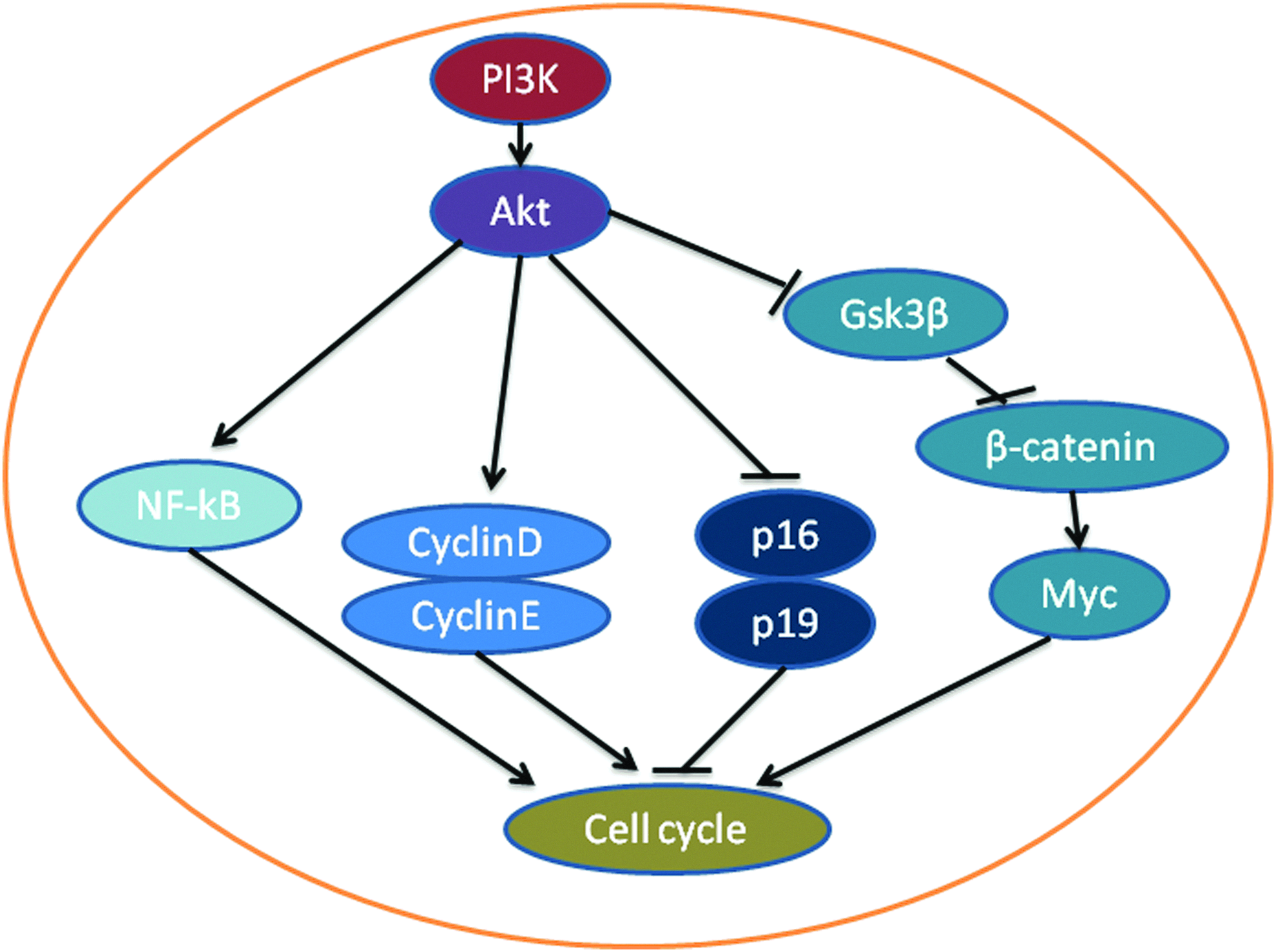
PI3K and the proliferation of MSCs. Phosphorylated Akt can block the p16 and p19 proteins and promote an increase in cell cycling of MSCs via cyclin D and E. Phosphorylated Akt also prevents GSK-3beta from forming a complex with beta-catenin, resulting in the accumulation of beta-catenin. Accumulated beta-catenin can increase the expression of myc, which is known to increase cell proliferation. Akt also increases proliferation via the nuclear factor-kappaB (NF-κB) pathway. Color images available online at www.liebertpub.com/teb
The role of the PI3K/Akt pathway in MSC proliferation has been demonstrated by activating the pathway by mutating the regulatory subunit of PI3K in MSCs. The subunit p85α negatively regulates the catalytic subunit p110. The introduction of p85α−/− into MSCs resulted in an increase in the level of pAkt. 78 It was observed that over a span of 10 continuous passages of cultured p85α−/− MSCs, the cell proliferation rate was maintained, as indicated by the increased number of cells and the increased formation of colonies. Increased cell cycling was indicated by a lower number of cells in G1 and a greater number of cells in G2. There were consistent changes in the proteins regulating the cell cycle, including increased levels of the cell cycle-promoting proteins cyclins D and E and reduced levels of the cell cycle blockers p16 and p19.
The PI3K/Akt and MSC migration
To be able to function properly, MSCs have to move to the appropriate place. MSCs are recruited into sites of injury by cytokines and chemokines so that these cells can help repair the injury. 79 MSCs also move along vessels sprouting from endothelial cells to facilitate vessel formation and maintenance. 80 Many studies have demonstrated that the migration ability of MSCs affects their efficacy.81,82 Therefore, a greater understanding of the mechanisms involved in MSCs migration will facilitate the application of these cells in tissue engineering.
Cell migration is closely related to changes in the cytoskeleton and to the formation of movement structures. The present theory of cell migration is summarized in several reviews.83,84 In brief, actin plays a key role in cell movement, 85 although several other structural proteins are involved in the cytoskeleton. During migration, cells become asymmetrical, forming front and back ends to direct cell movement. At the front, there are filopodia and lamellipodia. At the back, stress fibers, which are involved in cell adhesion, are formed or deformed to facilitate cell migration.
The PI3K/Akt pathway is well known to affect cytoskeleton changes in many cells, particularly in cancer cells. p70(S6K), a downstream target of the PI3K/Akt pathway, can regulate cytoskeleton dynamics via actin filament cross-linking proteins (Fig. 4). The overexpression of p70(S6K) in ovarian cancer cells promoted directional cell migration. 86 p70(S6K) stimulated the rapid activation of Rac-1 and CDC42 and their downstream effector, p21-activated kinase (PAK1). 86 Inhibition of p70(S6K) activity resulted in the decreased expression of these proteins and in actin cytoskeleton reorganization. In embryonic fibroblasts, Rac-1-null cells were defective in lamellipodia formation, cell spreading, cell-fibronectin adhesion, and focal contact formation when stimulated with platelet-derived growth factor (PDGF). 87 The phosphorylation of PAK1 was reduced, and the reconstitution of active PAK1 counteracted this effect. Girdin, a newly found actin-binding protein, is also a substrate of Akt. The phosphorylation of girdin by Akt is important to girdin's function in cell migration. Girdin is essential for the formation of stress fibers and lamellipodia in fibroblasts and endothelial cells.88,89 The RNAi knockdown of girdin was found to disrupt the rearrangement of the actin cytoskeleton in vascular smooth muscle cells. 90 The girdin-related migration of endothelial cells is involved in angiogenesis. 91 However, the role of girdin and its related signal pathways in MSC migration has not been studied.
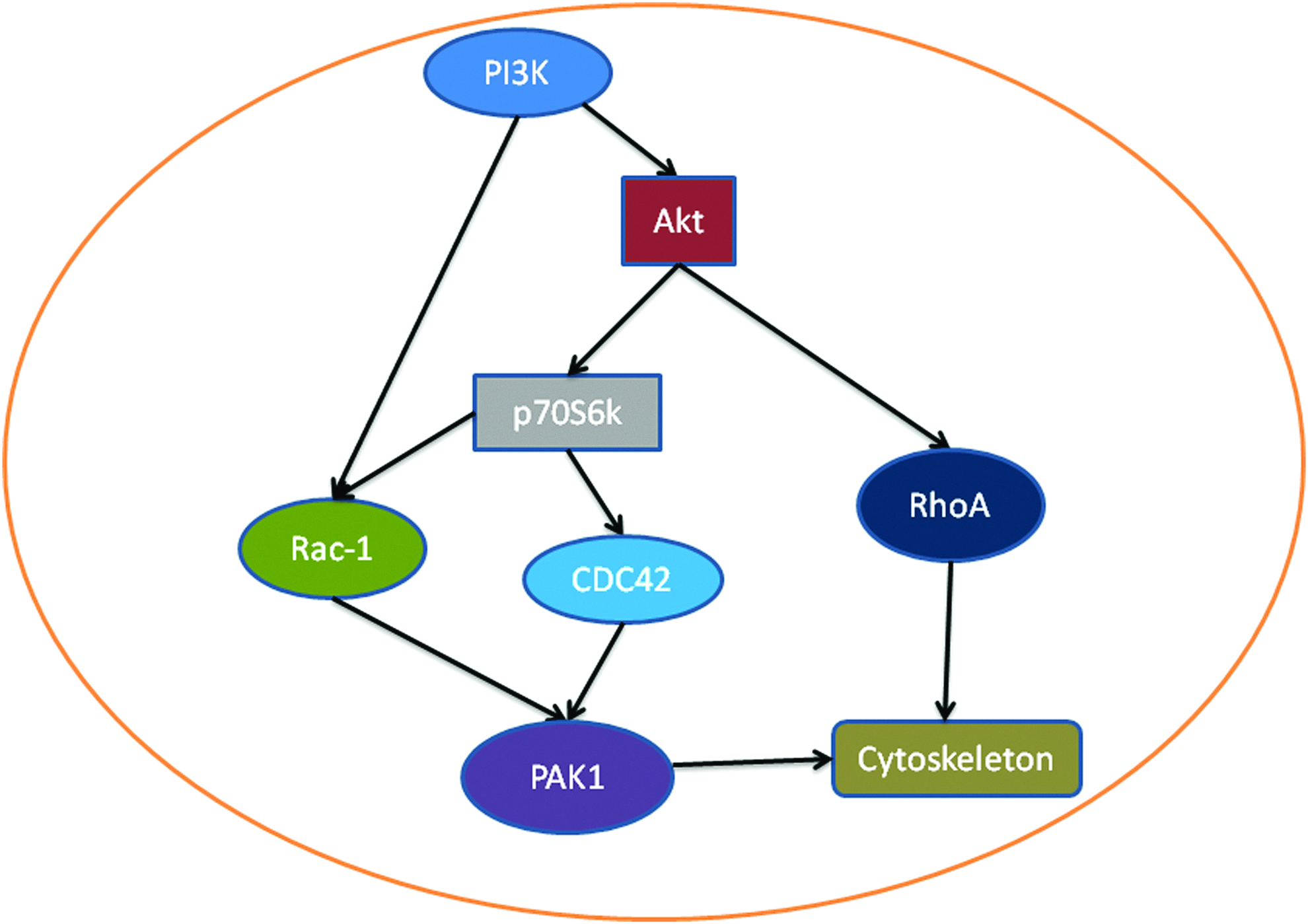
MSC migration promoted by the PI3K/Akt pathway. PI3K can activate Akt, which in turn increases the expression of RhoA, thus changing the cytoskeleton. Akt also phosphorylates p70S6K, which increases Rac-1 and CDC42 expression, leading to PAK1 activation and cytoskeleton changes. PI3K also directly acts on Rac-1. Color images available online at www.liebertpub.com/teb
In MSCs, several PI3K/Akt stimulators have been shown to increase MSC migration. The role of SDF-1, one such stimulator, in MSC migration has been well studied. 79 A study has shown that SDF-1 promoted MSC migration in a bone regeneration scaffold. 92 SDF-1-increased MSC migration was mediated by the activation of the PI3K/Akt pathway. 39 Hypoxia conditioning has also been shown to increase MSC migration via the PI3K/Akt pathway, and this effect was abolished by the PI3K inhibitors wortmannin and LY294002.93,94 However, the downstream target proteins of the PI3K/Akt pathway in MSC migration is not well understood and warrant further studies.
The PI3K/Akt pathway and MSC-induced angiogenesis
Angiogenesis is necessary for the survival of new tissues formed by tissue engineering because blood vessels are required to supply nutrients and transport out wastes. MSCs can help vessels to grow and invade from neighboring tissues by promoting the formation of tubes by endothelial cells. MSCs can recruit pericytes and smooth muscle cells to promote the maturation of newly formed blood vessels. 80 In addition, MSCs themselves can also differentiate into endothelial cells to increase vascularization. 95
Several studies have demonstrated that the activation of the PI3K/Akt pathway increases the angiogenesis ability of MSCs. Several stimulators of the PI3K/Akt pathway, such as SDF-1 and cobalt chloride (CoCl2), have been shown to increase angiogenesis. The overexpression of SDF-1 was also shown to increase angiogenesis. 26 MiR-126-overexpressing MSCs increased the level of angiogenesis in an infarcted area in mice subjected to ligation of the coronary artery by increasing the levels of pAkt and pErk. 96 The delivery of angiopoietin-1 with a Sendai virus-based vector to MSCs increased the pAkt levels and resulted in increased angiogenesis. 97 Adrenomedullin, an activator of the PI3K/Akt pathway, was shown to increase angiogenesis when it was infused together with transplanted MSCs. 25 The effect of MSCs on endothelial cells with respect to angiogenesis was also demonstrated using conditioned medium from MSC cultures. This medium increased the level of angiogenesis by increasing the level pAkt and the effect was inhibited by a PI3K/Akt inhibitor. 98
The PI3K/Akt and cytokine production of MSCs
MSCs can secrete many cytokines that regulate other cells and facilitate tissue repair; these cytokines include VEGF, FGF, monocyte chemoattractant protein-1, HGF, insulin-like growth factor-I (IGF-I), SDF-1, and thrombopoietin.99–101 The cytokines secreted by MSCs are required for MSCs to exert their effects in infarcted heart tissue because the numbers of surviving MSCs and differentiated cells derived from MSCs are low. 102 The secreted cytokines can promote the recruitment of local cells for injury repair. The role of cytokines secreted by MSCs as local regulators was further demonstrated by the ability of conditioned MSC medium to ameliorate myocardial infarction. 103 Conditioned medium from MSC culture also increased wound healing indicated by recruitment of macrophages and endothelial cells. 104 It has been shown that systemic delivery of BMSCs decreased neuropathy and this effect was reduced by neutralizing antibodies against brain-derived neurotrophic factor (BDNF), indicating importance of MSC-secreted factors. Secreted neuron growth factors such as nerve growth factor, BDNF, and glial cell line-derived neurotrophic factor also facilitate the recovery from Alzheimer's disease, Huntington's disease, Parkinson's disease, multiple sclerosis, and Krabbe's disease. 105
Evidence has shown that the MSC's ability to secrete cytokines is regulated by PI3K/Akt activity. SDF-1 has been shown to increase the secretion of cytokines by MSCs, and this effect is mediated by the PI3K/Akt pathway. 39 The overexpression of Akt1 in MSCs increased the gene expression of VEGF, FGF-2, HGF, and IGF-1 and thus increased the secretion of these cytokines. 103 Akt1-induced cytokine secretion has been linked to the increased survival of infarcted myocardial cells. interleukin-6 and TNF-alpha can synergistically costimulate MSCs to produce VEGF via the PI3K/Akt pathway. 106 NF-κB, a downstream protein in the PI3K/Akt pathway, was also shown to play an important role in the TNF-alpha-induced production of VEGF, FGF, HGF, and IGF-1. 107
These cytokines can also act on MSCs to affect their functionalities. For example, MSCs can express HGF receptor, c-Met.108,109 HGF-expressing MSCs have a greater ability to repair myocardial infarction, whereas HGF-null MSCs are unable to repair ischemic limbs.110,111 This effect of HGF was demonstrated to be mediated by the PI3K/Akt pathway. HGF at a concentration of 20 ng/mL promoted osteogenic differentiation via the activation of c-Met, Akt pathway, and the increased expression of cell cycle inhibitor p27 and osteogenic-associated transcription factors Runx2 and osterix, whereas a high concentration of HGF (100 ng/mL) promoted proliferation and suppressed osteogenic differentiation by activating Erk1/2 pathway and inhibiting Akt pathway.112,113
The PI3K/Akt pathway and MSC differentiation
The PI3K/Akt pathway has been shown to be important in the differentiation of MSCs into various cell types. However, the roles of this pathway are complicated by controversial effects. For example, the levels of components of the PI3K/Akt pathway and its downstream proteins mTOR, FOXO1, p27(kip1), and p70S6K are increased during the differentiation of MSCs into adipocytes.114–116 The inhibition of this pathway with the PI3K-specific inhibitor LY294002 resulted in decreased adipogenesis, indicating that the PI3K/Akt pathway plays an important role in adipogenesis. 114 The activation of the downstream protein mTOR is sufficient to promote adipogenesis, and this process is inhibited by rapamycin. It has been demonstrated that mTOR upregulates peroxisome proliferator-activated receptor gamma and CCAAT-enhancer binding protein alpha, two master adipogenic transcriptional factors 115 (Fig. 5). Further, the protein Lar (leukocyte common antigen-related tyrosine phosphatase) negatively regulates the adipogenesis of MSCs by inhibiting the PI3K/Akt pathway. 116
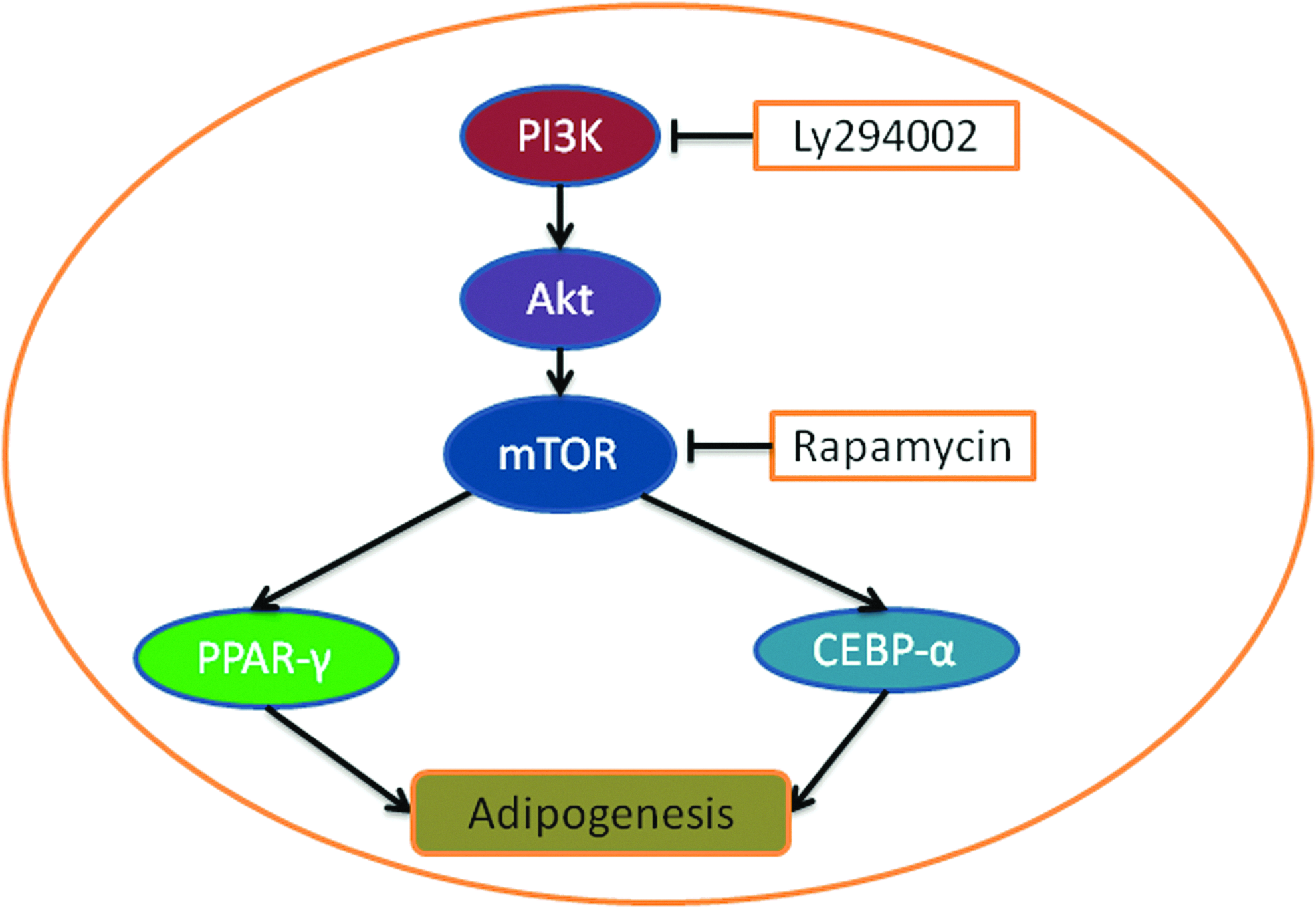
PI3K and adipogenesis. The PI3K/Akt pathway can regulate adipogenesis via mTOR, which can increase the levels of peroxisome proliferator-activated receptor gamma (PPAR-gamma) and CCAAT-enhancer binding protein alpha (CEBP-alpha). The PI3K-specific inhibitor LY294002 and the mTOR inhibitor rapamycin decrease the level of adipogenesis. Color images available online at www.liebertpub.com/teb
IGF-I has been expressed in MSCs to increase osteogenesis for bone repair, and the mechanism shown involved the activation of IRS1-PI3K. 106 Although IGF-1 can also activate mitogen-activated protein kinase (MAPK) pathway, several studies have shown that inhibition of MAPKs increases rather than decreases the rate of osteogenesis,117–120 indicating the PI3K/Akt pathway plays the key role in IGF-1-induced osteogenesis. TNF-alpha was shown to increase osteogenesis by activating NF-κB in MSCs from human adipose tissue; this induction of osteogenesis by TNF-alpha was blocked by an NF-κB inhibitor. 121 The mechanically induced activation of Akt has also been shown to increase osteogenesis.122,123
Several studies have yielded controversial results about the role of the PI3K/Akt pathway in adipogenesis and osteogenesis. Fitter et al. showed that imatinib promoted adipogenesis by inhibiting PDGF-induced PI3K activity. 124 Controversial results regarding the role of the PI3K/Akt pathway in osteogenesis have also been presented.125–131 Studies have shown that imatinib stimulates osteogenesis by inhibiting PDGFR/PI3K/Akt signaling in MSCs.132–134 The reason for this effect is not well understood. It is possible that PDGF induces adipogenesis via signaling pathways different from those involved in the stimulation by special adipogenic medium (1 μM dexamethasone, 0.2 mM indomethacin, 0.5 mM 3-Isobutyl-1-methylxanthine, 0.01 mg/mL insulin, and 10% fetal bovine serum). It is now known that multiple signal pathways are involved in lineage determination. Canonical and non-canonical Wnts are involved in determining whether MSCs differentiate into osteocytes or adipocytes.135,136 FGF was demonstrated to increase osteogenesis via FGFR1 and 2, which activate the PI3K/Akt and Erk1/2 pathways. 137 The role of the interaction of multiple signal pathways in MSC differentiation has also been studied. 138 However, no conclusive explanation has been revealed by the results.
The importance of PI3K/Akt pathway in chondrogenesis has also been demonstrated. Insulin was shown to increase chondrogenesis via activation of PI3K/Akt pathway. 139 IGF-1 also promoted chondrogenesis via the same pathway and its downstream target protein NF-κB. 140 Delta-like 1 protein, which inhibits Akt but not Erk1/2 and p38 MAPK reduced insulin-induced chondrogenic differentiation as demonstrated by a reduction of cartilage pellet formation and expression of genes involved in chondrogenesis such as aggrecan, collagen Type II, and X. 139 Addition of Delta-like 1 protein into chondrogenic medium produced a dose-dependent inhibition of chondrogenesis that was reversed by fibronectin, an activator of the PI3K/Akt pathway. 139
MSCs may also differentiate into other cells types such as neurons, cardiomyocytes, hepatocytes, and pancreatic beta-cells. 35 However, the role of PI3K/Akt pathway in the differentiation of these cells has not been studied well. Further exploration of the pathway may help to promote the potential use of MSCs in the regeneration of these cells.
The roles of PI3K/Akt pathway in the functionalities of ASCs have also been studied but in a much less extent than that of BMSCs. It has been demonstrated that stimulation of the activity of Akt promoted the differentiation of ASCs into adipose tissue while inhibition of the pathway by Wartmannin reduced adipose tissue formation. 141 Hypoxia condition has been shown to increase the survival ability, proliferation, angiogenesis, growth factor secretion, and migration of ASCs via HIF-alpha/VEGF-A/PI3K/Akt pathway.142–146 Both neutralizing antibodies against VEGF-A and inhibitor of PI3K may abolish the effect of hypoxia condition. These studies indicate that the pathway may also play a key role in ASCs in all aspects described above. More studies about the roles of the PI3K/Akt pathway in ASCs are warranted.
The Effect of Biomaterials on the Activity of PI3K/Akt Pathway in MSCs
Effective tissue engineering such as bone regeneration required optimal interactions of three components including MSC cells, biomaterials used for scaffolds, and bioactive molecules.147–151 Biomaterials are used not only for mechanical support but also stimulation of MSCs. At present many different kinds of biomaterials have been used to make scaffolds including polymers, calcium phosphate (CaP), and bioglass etc. Current studies in biomaterials attempt to increase their capability to stimulate MSCs to form new tissues. For the purpose, biomaterials with new properties have been made, surface structure of biomaterials has been modified, and growth factors have been loaded to scaffolds. Biomaterials-increased MSC functionalities have been associated with the activation of the PI3K/Akt pathway as described in the following sections.
Change of nature of biomaterials
Novel biomaterials have been made so that they have increased capability to increase MSC functionalities. CaP such as β-tricalcium phosphate and hydroxyapatite (HAP) possesses good mechanical properties, biocompatibility, and osteoconductivity, but it lacks biodegradability. 12 Polymers are more popular for making scaffolds due to their property of biodegradation. However, the disadvantage of polymers is that their osteoconductivity is very low. Woo et al. developed a novel material using a mixture of HAP and polymer PLLA called PLLA/HAP. 152 In an in vitro culture system, this novel biomaterial has been shown to increase MSC cell survival as indicated by DNA content and decrease apoptosis detected by Tunel assay. The mechanism was demonstrated that PLLA/HAP could absorb more fibronectin and vitonectin, which increased the activity of the PI3K/Akt as indicated by increased levels of pAkt and its downstream target bcl-2. This study provides a good example that manipulation of biomaterials can increase MSC functionalities via the activation of PI3K/Akt pathway. At present, many biomaterials have been studied to increase MSC functionalities. However, the involvement of the PI3K/Akt is not extensively studied. It will be interesting to investigate how biomaterials affect MSC different functionalities through the PI3K/Akt and its downstream target proteins. Our studies have shown that CoCl2 can increase MSC proliferation and differentiation. 49 It also increased MSC secretion of VEGF and thus increased vascularization in animal model. 45 The PI3K/Akt pathway is likely to be involved in these effects. Several studies have shown that CoCl2 induced hypoxia, which in turn activated the PI3K/Akt pathway to mediate CoCl2 biological effects.153–155 Further studies are warranted to characterize the direct effect of CoCl2 on the activity of the PI3K/Akt pathway and its downstream target proteins in MSCs.
Modification of biomaterial surface
Modification of the surface of biomaterials has also been used to increase MSC functionalities. Attachment of peptides to the surface of biomaterials has been shown to increase their stimulatory effect on MSCs. 151 Several studies demonstrated that peptide Arg-Gly-Asp (RGD) attachment increased MSC survival, proliferation, and differentiation.156–160 Zhang et al. demonstrated that RGD attached poly (caprolactone) (PCL) increased MSC proliferation and this was mediated by the activation of the PI3K/Akt pathway. 161 RGD activated an integrin that in turn increased Akt activity.
Control of growth factor release in scaffolds
Several growth factors such as FGF, TGF-beta, and bone morphogenetic protein-2 (BMP-2) have been loaded on scaffold biomaterials to increase the efficacy of bone formation.15,162 BMP-2 has been incorporated into the biomaterial surface by several methods such as indirect adsorption, affinity binding, or chemically conjugating.163,164 The loading of BMP-2 in scaffold biomaterials CaP, tricalcium phosphate, and PCL increased bone formation.165–168 Release pattern of BMP-2 from scaffolds is recognized to be important and scaffold geometry has been used to achieve gradient release of BMP-2.169–172 BMP-2 is known to increase osteogenesis via the activation of PI3K/Akt pathway. 173 It will be interesting to study how BMP2 and other growth factors loaded in scaffolds and their release pattern affect the PI3K/Akt pathway in MSCs and thus alter MSC functionalities in tissue engineering apparatus.
Conclusions and Future directions
The bottleneck for MSCs in tissue engineering application is that their survival ability and functionality are very low. 38 This could be overcome by activating the PI3K/Akt pathway in MSCs, which plays key roles in the functionalities of MSCs including cell survival ability, cell proliferation, migration, angiogenesis, cytokine production, and differentiation as discussed above. Therefore, activation of the pathway will increase the performance of MSCs in tissue engineering. Indeed, several approaches have been applied to increase MSC functionalities including overexpression of genes, addition of growth factors, and manipulation of biomaterials.43–48 Several studies showed that novel biomaterials, use of CoCl2, incorporation of peptides on the surface of biomaterials, and loading growth factors in scaffolds can increase MSC functionalities.42,49 Biomaterial-increased MSC functionalities have been associated with the activation of the PI3K/Akt pathway.
In future, the ability of a biomaterial to activate PI3K/Akt in MSCs in vitro system could be used for selection criteria for the use in tissue engineering. Activation of the PI3K/Akt pathway could indicate the effect of biomaterials on MSC functionalities. It will be interesting to study how biomaterials affect the downstream target proteins of the PI3K/Akt pathway associated with different functionalities of MSCs. How loaded growth factors in scaffolds activate the PI3K/Akt and its downstream targets in MSCs could be studied. MSCs have been genetically engineered to overexpress regulatory genes to increase their functionalities in cellular therapy. It is interesting to investigate whether biomaterials have synergistic effect with these MSCs. Combinational applications of genetic engineered MSCs, growth factors, and bioactive biomaterials could be an effective approach for successful tissue regeneration.
Acknowledgment
Y.X. is supported by grants from the National Health and Medical Research Council (NHMRC). C.C. is also supported by an NHMRC grant.
Footnotes
Disclosure Statement
No competing financial interests exist.