
Abstract
Tissue engineering and regenerative medicine (TERM) remains to be one of the fastest growing fields, which covers a wide scope of topics of both basic and applied biological researches. This overview article summarized the advancements in basic researches of TERM area, including stem cell biology, cell engineering, somatic nuclear transfer, genomic editing, discovery of new tissue progenitor/stem cells, and immunomodulation of stem cells and tissue regeneration. It reflects the cutting-edge achievements in basic researches, which will lay solid scientific foundation for future TERM translational researches.
The Aim and Methods
“Y
Advancements in Stem Cell Biology
Environmental factors contribute significantly to the regulation of stem cell functions. These factors can be generally divided into extrinsic and intrinsic categories. In last year literature, progresses were made in both areas.
Systemic hormone or growth factor levels are typical extrinsic factors, which are likely to affect stem cell functions. Nakada et al. reported an interesting finding that estrogen could increase hematopoietic stem cell (HSC) self-renewal in females and particularly during pregnancy. They demonstrated that HSCs of female mice divided significantly more frequently than those of male mice. Administration of estrogen increased HSC division in both males and females. Enhanced HSC division, frequency, increased cellularity, and splenic erythropoiesis were observed during pregnancy due to the increased estrogen level. They also demonstrated that the interaction between estrogen and its receptor provided the signaling for enhancing HSC self-renewal and promoting erythropoiesis during pregnancy. 3 In addition to sexual hormones, dietary and metabolic controls might also have an impact on stem cell functions in physiology and cancer (Fig. 1). 4
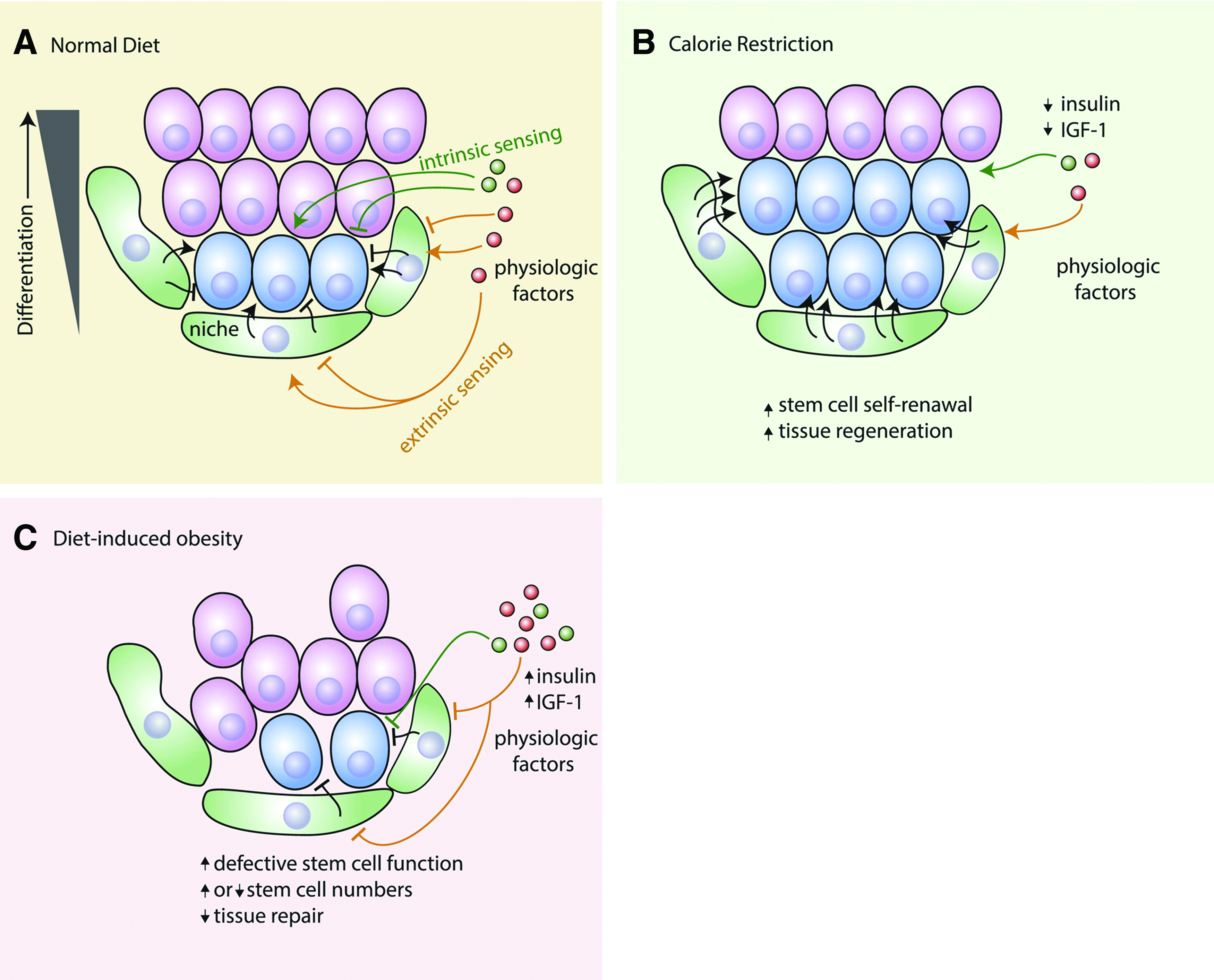
Dietary regulation of stem cells in tissue homeostasis.
In stem cell biology research, much effort was made to define the signals that control proliferation and differentiation of tissue-specific adult stem cells or progenitor cells, but little is known about the substrates and the signals that guide migration of stem cells out of their niche. Using Drosophila tracheal outgrowth during metamorphosis as a model, Chen and Krasnow discovered that the embryonic tracheal inducer branchless fibroblast growth factor was expressed dynamically just ahead of progenitor outgrowth in decaying branches and thus guided the cell migration. By contrast, knockdown of this molecule abrogated cell outgrowth, whereas misexpression of this molecule could redirect the migration, suggesting that both tissue substrate and growth factor provided important signals to guide progenitor-mediated tissue development, which can potentially be used to guide stem cell-mediated tissue regeneration. 5
The mechanism studies of tissue development likewise provide clues to tissue regeneration such as neuronal regeneration. O'Donovan et al. used a fetal development model to demonstrate that activation of B-RAF kinase alone in mouse embryonic neurons was sufficient to drive the growth of long-range peripheral sensory axon projections in vivo even without the presence of upstream neurotrophin signaling. They further demonstrated that activated B-RAF signaling induced robust regeneration of sensory axons into the spinal cord after a dorsal root crush and also substantial axon regrowth in the crush-lesioned optic nerve. 6 The three-dimensional (3D) genome is another possible element to serve as an intrinsic factor for transcriptional regulation and pluripotency. Particularly, the relationship between genome organization and lineage-specific transcriptional regulation reflects an emerging trend. 7 DNA methylation is known to help maintaining stable gene regulatory programs, which are essential for the normal functions in multicellular organisms. It was found that there were differences between stem cells and somatic cells in their ways to stay in an epigenetic state. According to Shipony et al., human embryonic stem cells (ESCs) preserve their epigenetic state by balancing the antagonistic process that adds and removes DNA methylation. By contrast, somatic cells transmit considerable epigenetic information to their progenies through copying epigenetic information from mother to daughter cells. 8
In vitro differentiation of pluripotent stem cells into desired cell types is a subject deserving great attention, but remains a difficult task due to the requirement of complex undefined medium constituents, which may hinder further elucidation of the molecular mechanisms of a specific lineage differentiation. Last year, one surprising finding of stem cell differentiation was to define how few medium components were required to differentiate human-induced pluripotent stem cells (hiPSCs) into cardiomyocytes. Burridge et al. established a protocol to derive TNNT2+ cardiomyocytes from hiPSCs with a chemically defined medium consisting of only three components: the basal medium RPMI 1640, l-ascorbic acid 2-phosphate, and rice-derived recombinant human albumin. Also importantly, synthetic vitronectin peptides were needed as a specific substrate for hiPSC adhesion. This method was proven effective in 11 tested hiPSC lines and could reach a yield of 100 cardiomyocytes for every input pluripotent cell. 9
Recent studies indicate that vascular smooth muscle cells (SMCs) are no longer a homogenous cell population, but different subtypes arisen from diverse developmental origins. Naturally, derivation of different subtypes of vascular SMCs from different tissue origins is critical not only for the mechanism study of various vascular diseases but also for the regeneration of different types of tissues that contain their own subtypes of SMCs. Cheung et al. reported directed differentiation of embryonic origin-specific vascular smooth muscle subtypes from human pluripotent stem cells (hPSCs) through chemically defined induction of developmental stage-specific lineages, 10 indicating the potential of induced pluripotent stem cells (iPSCs) in vascular tissue regeneration.
In addition to vascular SMCs, endothelial cells (ECs) and pericytes are also the major cellular components of vascular structure. Orlova et al. developed a protocol to generate these two types of cells from hPSCs under defined conditions. 11 They further verified the functions by showing primary vascular plexus formation upon coculture with hPSC-derived pericytes and the incorporation of the cells into the vasculature of zebrafish xenografts in vivo. Therefore, hPSC-derived SMCs and ECs are possible to generate human vascular structures, and the incorporation of hPSC-derived pericytes may also generate an engineered human vessel with self-repair function.
Guan et al. performed biological characterization on human urine-derived stem cells (USCs) and found that USCs had similar surface antigen profiles and multiple differentiation potential. Particularly, they could be differentiated into neural lineage and survive after transplantation into rat brain. 12
To translate stem cell research into clinical trial, nonhuman primates provide optimal models for preclinical study and further development of stem cell therapies. Li et al. reported the first derivation of neural crest stem cells from cynomolgus monkey embryonic stem cells (CmNCSCs) at the neural rosette stage. They could be induced to differentiate toward peripheral nervous system lineages in vitro. After in vivo implantation into fetal brains of cynomolgus macaques, CmNCSCs survived, migrated, and differentiated into progeny consistent with a neural crest identity and, thus, offering a new tool for investigating neural crest development and neural crest-associated human disease. 13
Advancements in Cell Engineering
Somatic cell reprogramming by gene transfer or nuclear transfer, directed differentiation of pluripotent stem cells, and directed conversion between differentiated cell lineages represent powerful approaches to engineer cells for research purpose and for regenerative medicine. However, how engineered cells closely resemble the molecular and functional identities of original counterpart cells lacks the metrics for evaluation. CellNet, a novel network biology platform reported by Cahan et al., provides such a means to accurately assess the fidelity of engineered cells. 14
CellNet is a computational platform that evaluates gene regulatory networks (GRNs) of target cells. Ideally, a comprehensive catalog of GRNs can be established by chromatin immunoprecipitation followed by sequencing (ChIP-seq) for all cell types and all transcription factors. In this report, the authors developed a pipeline to reconstruct GRNs by using 3419 publicly available gene expression profiles of diverse cell types and tissues. With this platform, they analyzed the expression data of 56 published reports and found that cells derived through directed differentiation more closely resembled their in vivo counterparts than products of direct conversion using neurons and cardiomyocytes as the examples. Furthermore, directly converted cells failed to adequately silence expression programs of the starting population and that the establishment of unintended GRNs was common to almost every cellular engineering paradigm. 14
Another study from Morris et al. further indicates the value of CellNet to guide the practical application of stem cells in tissue regeneration. 15 They used CellNet to improve the conversion of B cells to macrophages transcriptionally and functionally through knocking down predicted B-cell regulators. In another example of converting fibroblasts to induced hepatocytes (iHEPs), CellNet revealed that iHEPs failed to classify as a liver. By contrast, CellNet analysis yielded the surprising discovery that iHEPs in fact represented an induced endoderm progenitor (iEP), which was capable of long-term functional engraftment of the mouse colon. These results further illustrate that CellNet can be employed to improve direct conversion and to uncover unappreciated properties of engineered cells.
In last year's literature, there were some progresses in achieving desired cell sources for tissue regeneration. Direct reprogramming from mature differentiated cells to specific tissue progenitor cells remained a hot topic of research.
Riddell et al. reported the reprogramming of committed lymphoid and myeloid progenitors as well as myeloid effector cells into induced hematopoietic stem cells (iHSCs) by transfection of six transcription factors Run1t1, Hlf, Lmo2, Prdm5, Pbx1, and Zfp37. In addition, inclusion of Mycn and Meis1 and the use of polycistronic viruses further increased the reprogramming efficacy. Functional evaluation with single-cell analysis also revealed that iHSCs derived under optimal conditions exhibited a gene expression profile that was highly similar to that of endogenous HSCs. 16
Generation of iPSCs by using Yamanaka factors to reprogram somatic cells is usually slow and inefficient because it is considered as a stochastic process. Guo et al. reported a new strategy to enhance iPSC reprogramming efficiency by identifying a privileged somatic cell state, from which acquisition of pluripotency could occur in a nonstochastic manner. The privileged cells were characterized by an ultrafast cell cycle around 8 h, and more than 99% of the bulk reprogramming activity occurred in this ultrafast cycling population. This study demonstrated that the stochastic nature of reprogramming could be overcome in a privileged somatic cell state, and the acceleration of cell cycle toward a critical threshold was a vital bottleneck for reprogramming. 17
The other way to enhance somatic cell reprogramming efficiency is to pick optimal reprogramming factors. Buganim et al. reported that the ectopic expression of Sall4, Nanog, Esrrb, and Lin28 (SNEL) in mouse embryonic fibroblasts (MEFs) could generate high-quality iPSCs more efficiently when compared to conventional Yamanak factors of Oct4, Sox2, Klf4, and Myc. 18
Conventionally, reprogramming of somatic cells is commonly achieved by overexpression of a few key factors through gene transfer. By contrast, Sancho et al. reported a different approach to reprogram adult pancreatic ductal cells into α, δ, and β cells. 19 From developmental biology point of view, Ngn3 is a key regulator of endocrine cell differentiation, whereas Fbw7 maintains adult ductal cell fate. Interestingly, their study showed that Fbw7 inactivation resulted in transdifferentiation of ductal cells into α and δ cells, and predominantly, β cells. More importantly, the newly formed β cells resembled islet β cells in term of their cellular morphology, marker gene expression, and insulin secretion in response to glucose challenge.
The role of small molecules in somatic cell reprogramming is also a popular theme in stem cell research. Theunissen et al. reported that a combination of five kinase inhibitors could induce and maintain the OCT4 distal enhancer activity when applied directly to conventional human ESCs. Furthermore, the use of these inhibitors could generate human pluripotent cells with a distinct pluripotent state that closely resembled the state of mouse ESCs. 20 The established methodology presents a framework for defining the culture requirements of naive human pluripotent cells. Li et al. also reported that small molecules could facilitate the reprogramming of mouse fibroblasts into pancreatic lineages. 21 Besides small molecules, long noncoding RNAs have also been applied to program and reprogram cell fate, as reviewed by Flynn and Chang. 22
Sakurai et al. recently reported an interesting study using kinome-wide RNAi-based analysis to identify the kinases that regulate somatic cell reprogramming to iPSCs, and they eventually found that seven key kinases served as barriers to iPSC generation. 23 Interestingly, shRNA-mediated knockdown of serine/threonine kinases TESK1 or LIMK2 not only promoted mesenchymal-to-epithelial transition but also decreased COFILIN phosphorylation and disrupted actin filament structures during reprogramming of MEFs. In fact, reorganization of the actin cytoskeleton is critical in many cellular functions, including motility, adhesion, morphogenesis, and cytokinesis. The results of this particular study indicate that successfully reprogrammed cells must have actin depolymerization to achieve their well-established ES-like morphology and, thus, highlight the role of cytoskeletal remodeling in somatic cell reprogramming. 23
In last year's literature, progress was achieved as well in direct reprogramming from mature differentiated cells to different types of cells to generate functional cell source for tissue regeneration. Huang et al. reported the reprogramming of human fetal fibroblasts into human-induced hepatocytes (hiHEPs) through lentiviral transfection of transcription factors FOXA3, HNF1A, and HNF4A. The resulting hiHEPs not only displayed functional characteristics of mature hepatocytes in vitro but also could restore liver function and prolong survival upon in vivo implantation into an acute liver failure mouse model. 24 Another report by Du et al. demonstrated that functional hiHEPs could be generated from fibroblasts by overexpressing hepatic fate conversion factors HNF1A, HNF4A, and HNF6 along with the maturation factors ATF5, PROX1, and CEBPA. Additionally, these hiHEPs demonstrated their drug metabolism functions as primary hepatocytes did. 25
Kim et al. reported the direct conversion of mouse fibroblasts into induced neural stem cells (iNSCs) using four transcription factors Sox2, Klf4, Myc, and Pou3f4, and this combination did not generate full pluripotency status and thus to avoid the risk of tumor formation. These iNSCs could also differentiate into neurons, astrocytes, and oligodendrocytes with the silencing of retroviral transgene 2 months postreprogramming. 26 Gonzalez-Muñoz et al. also reported that histone chaperone ASF1A was required for maintenance of pluripotency and cellular reprogramming. 27
Reprogramming can also be applied to enhance expansion of purified stem cells. For example, CD34-positive cord blood stem cells have extensive hematopoietic capacity and usually lose their marrow-repopulating potential during in vitro expansion. Chaurasia et al. illustrated that epigenetic reprogramming of dividing CD34+ cord blood cells with the treatment of the most active histone deacetylase inhibitor valproic acid (VPA) followed by an initial 16-h cytokine priming could significantly increase the number of multipotent cells (CD34+CD90+). Compared to regularly expanded cells, VPA-treated CD34+ cells produced a greater number of SCID-repopulating cells and established multilineage hematopoiesis in primary and secondary immunodeficient recipient mice. 28
One major challenge in stem cell therapy is the difficulty in controlling cell fate and phenotype. Ankrum et al. introduced the development of an intracellular drug carrier, which was made of poly(lactic-co-glycolic acid) polymer generated by a single-emulsion evaporation technique. They further demonstrated that the particles with a size around 1 μm were able to be uptaken by mesenchymal stem cells and kept internalized within the cells for several weeks. This new platform can be used to upload drug or growth factors so as to well control stem cell proliferation, differentiation, phenotype, and other biological characters. 29
The central nervous system is also one of the focuses in the area of somatic cell reprogramming for generating neuronal progenitor cells to treat neuronal disease or regenerate tissues of central nervous system. Recent progress, challenge, and the future of induced neuronal cells were well summarized by Amamoto and Arlotta. 30
Advancements in Somatic Nuclear Transfer
Among the techniques of cell engineering, somatic cell nuclear transfer (SCNT) is a vital strategy to generate hPSCs, that is, isogenic ESCs to a patient, and thus, it is also called therapeutic cloning strategy. Nevertheless, the development of iPSCs seemed to override SCNT, because iPSCs are more efficient to generate and able to avoid the ethical issue of using human oocytes. Despite the apparent trend of using iPSCs to replace SCNT, whether iPSCs can fully match their native counterpart, ESCs, remains to be confirmed. As a result, SCNT research still continues and as a matter of fact, significant progress in this area is quite evident in last year's literature and several articles are embodied in this overview.
Although it was reported by the Mitalipov group that the SCNT approach finally produced stem cell lines from the DNA of human fetal and infant cells, it was not clear whether this technique equally worked for human adult cells. 31 In last year's literature, there are two reports of therapeutic cloning to generate human ESCs derived from adult human cells. One is the report by Chung et al. 32 They successfully generated human ESCs through SCNT using dermal fibroblasts from 35- and 75-year-old males. The generated human ESCs expressed OCT-4, SSEA-4, TRA-1-60, and TRA-1-81, had normal male karyotypes, nuclear DNA, and mitochondrial DNA genotyping demonstrated that nuclear DNA and mitochondrial DNA were exclusively derived, respectively, from patients' fibroblasts and donated oocytes.
In another report, Yamada et al. systematically investigated the parameters affecting the efficiency of blastocyst development and stem-cell derivation during the SCNT process. They showed that the use of both kinase and translation inhibitors, as well as the cell culture in the presence of histone deacetylase inhibitors, could significantly promote the development to blastocyst stage. They went a step further by generating ESCs from a 32-year-old woman who had type I diabetes. The SCNT-derived stem cells were pluripotent and could be differentiated into insulin-producing β cells and, therefore, could be potentially used for therapeutic cell replacement. 33 As also discussed in this article, iPSCs have been reported often as differentiation defective, containing aberrant patterns of cytosine methylation and hydroxymethylation, acquired somatic coding mutations, and showing biallelic expression of imprinted genes. The generated pluripotent human stem cells through the SCNT approach may serve as a good cell model to compare and evaluate the quality of iPSCs generated from two different strategies.
Genomic Editing
Because of the advancement in gene sequencing, whole-genome sequence is now available for manipulation. Genomic editing is a novel technology that uses engineered nucleases to generate defined genetic mutations or correct gene mutation. 34 Recently, this technology has been applied to stem cells too. Genovese et al. reported targeted genomic editing to correct IL2RG gene of HSCs derived from a patient of X-linked severe combined immunodeficiency (SCID-X1), and gene-edited HSCs could sustain normal hematopoiesis and give rise to functional lymphoid cells, indicating a new therapeutic strategy for treating SCID-X1 and other diseases. 35
hPSCs, including ESCs and iPSCs, are a good cell model to investigate the mechanism of a disease. In fact, iPSCs derived from patients are commonly used to establish cell models of various diseases. In last year literature, González et al. reported an iCRISPR platform that allows for rapid and versatile genome editing. 36 They showed that this platform was able to generate hPSCs with biallelic knockout for loss-of-function studies and homozygous knockin hPSCs with specific nucleotide alterations for precise modeling of disease conditions. Furthermore, the CRISPR/Cas system could efficiently in one-step generate double- and triple-gene knockout hPSC lines and stage-specific inducible gene knockout during hPSC differentiation.
In another article, Miyaoka et al. developed a method using droplet digital PCR and adapted iPSC growth conditions to efficiently isolate single-base genome-edited human iPSCs and, thus, to create desired mutated disease iPSCs with unprecedented efficiency. 37 Genomic editing through cell-autonomous correction of genomic abnormality can also occur naturally. For example, ring chromosomes are structural aberrations commonly associated with birth defects, mental disabilities, and growth retardation. In a study, Bershteyn et al. reported that during the process of generating human iPSCs from patient fibroblasts containing ring chromosomes with large deletions, they found that reprogrammed cells lost the abnormal chromosome and duplicated the wild-type homologue through the compensatory uniparental disomy mechanism. This interesting result implies that cellular reprogramming may serve as a means of chromosome therapy to reverse combined loss-of-function across many genes in cells with large-scale aberrations involving ring structures. 38
Discovery of New Tissue Progenitor Cells/Stem Cells
Tissue progenitor cells are an exceptional cell source for maintaining tissue structure and function, as well as for regenerating damaged tissues. In last year's literature, there were several reports of identifying new tissue/organ progenitor cells/stem cells with conventional or newly discovered biomarkers.
Isolation of endogenous cardiac stem and progenitor cells (eCSCs/eCPCs) has been reported previously using a very complicated marker cocktail. By contrast, Smith et al. reported the use of c-Kit+CD45−Tryp− as an effective marker to isolate and characterize the resident endogenous c-Kit+ cardiac stem cells from adult C57/BL6J mouse and Wistar rat hearts. 39 The key step was the retrograde coronary perfusion and digestion of the whole heart, which allow the collection of the highest number of eCSCs. This finding may provide a practical method for further investigation of eCSC-mediated heart regeneration.
Desai et al. illuminated the role of alveolar progenitor and stem cells in lung development and found that squamous alveolar type 1 (AT1) cells and cuboidal surfactant-secreting AT2 cells arose directly from a bipotent progenitor. AT2 cells functioned as the stem cells that contributed to alveolar renewal, repair, and cancer through the signal transduced by EGFR-KRAS. 40
HSCs probably are the first adult stem cells that have been identified. Conventionally, immunophenotype characterization with flow cytometry is the principle technique for identifying various types of stem cells, including HSCs. Gazit et al. reported the use of systems-wide microarray screen to reveal that Fgd5 expression in bone marrow was exclusively restricted to HSCs. They further used the Fgd5 reporter gene system to demonstrate that Fgd5 was not required for definitive hematopoiesis, but was able to identify bone marrow cells that had potent HSC activity. 41 Compared to the conventional stringent purification procedure of HSCs, this simple method may facilitate the rapid progress of HSC-related basic and translational researches.
Karthaus et al. reported the identification of multipotent luminal progenitor cells in human prostate organoid cultures. 42 They developed a 3D culture system that was able to support the growth of human prostate organoids composed of fully differentiated CK5+ basal and CK8+ luminal cells. Moreover, using this model, they discovered that single human luminal and basal cells gave rise to organoids, but luminal cell-derived organoids more closely resembled prostate glands, indicating the presence of luminal multilineage progenitor cells. The isolated cells as well as the robust scalable 3D culture system may allow for mechanistic studies of human prostate tissue development and regeneration.
In mammary gland, the existence of multipotent stem cells remains a controversial issue due to the apparent disparity in the research conclusions when using the transplantation model versus the lineage-tracing model. Rios et al. reported the discovery of bipotent stem cells as well as distinct long-lived progenitor cells in the mammary gland through clonal cell-fate mapping studies using a stochastic multicolor cre reporter and a new 3D imaging method. They concluded that the bipotent stem cells mainly coordinate ductal homeostasis and remodeling of the mouse adult gland, whereas both stem and progenitor cells are responsible for morphogenesis during puberty. 43
Wound regeneration probably is one of the best models to investigate tissue regeneration mechanism. Dermal fibroblasts are considered as connective tissue supporting cells with simple functions. However, Driskell et al. discovered that there are two distinct types of dermal fibroblasts, which play different functions in skin. 44 The one from upper part of the dermis forms the upper dermis, including the dermal papilla, which regulates hair growth and arrector pili muscle, whereas the other one forms the lower dermis, which is the major cell type for producing bulk of the fibrillar extracellular matrix. Hair follicle regeneration requires the presence of upper part dermal fibroblasts, whereas the hypodermis fibroblasts cause fibrosis upon skin injury.
Tissue/organ regeneration after various injuries is generally thought to be mediated by localized tissue progenitor cells or stem cells from a remote area such as bone marrow. The liver is a unique organ that is characterized by its potent capacity of self-regeneration upon injury. In response to liver damage such as toxin-induced injuries, an accumulation of atypical ductal cells (ADCs), also commonly called oval cells, could be observed in rodents or human livers. Oval cells are considered as liver stem cells and thus were thought as the major cell source for regenerating liver tissue based on in vitro assays. However, a report from Yanger et al. subverted the conventional concept that tissue has to be regenerated from stem cells. 45 They used both direct genetic and unbiased nucleoside analog-based lineage-labeling tools to demonstrate that ADCs did not exhibit progenitor cell activity in vivo. More importantly, most of in vivo regenerated hepatocytes were actually derived from pre-existing hepatocytes. Whether other mature tissue cells have functional roles similar to hepatocytes in tissue regeneration is certainly worth investigating. Interestingly, Hu et al. reported that liver regeneration also depended on spatially and temporally precisely coordinated proliferation of both hepatocytes and liver sinusoidal endothelial cells to reconstitute liver structure and function, and this was accomplished by EC-derived angiopoietin-2 control of liver regeneration as a spatiotemporal rheostat. 46
Immune Modulation of Stem Cells and Tissue Regeneration
Stem cell-mediated tissue regeneration depends on the proper interplay between stem cells and their niche environment, which provides a proper “Soil” for stem cells (“Seed”) to grow and function. 47 Last year, the issue of immune modulation on stem cells and tissue regeneration was brought to attention in several prestigious journals. Aurora et al. reported an interesting finding that myocardial infarction could be fully regenerated in neonatal mice. However, this insult resulted in scar formation in adult mice due to the triggered immune response that not only aimed to clear debris and restore tissue integrity but also led to an evitable tissue scarring with compromised cardiac function. The critical time point was postnatal day 7, after that, the mice lost the regenerative ability. By comparing the hearts between postnatal day 1 and 14, they further discovered the presence of regenerative macrophages in neonatal heart, which had a unique polarization phenotype and secreted numerous soluble factors that were likely to facilitate myocardial regeneration. Furthermore, deletion of the regenerative macrophages led to the failure in neonatal heart regeneration, 48 indicating that a subset of macrophages could potentially be exploited to recapitulate regenerative function in adult heart. 49 In addition to macrophage, a complement may also exert its significant effect on tissue regeneration. Using posthepatectomy liver regeneration as a model, Marshall et al. reported an interesting finding that application of a novel site-targeted murine complement inhibitor, CR2-CD59 which specifically inhibits the terminal membrane attack complex, could remarkably enhance regeneration after 90% hepatectomy and improve long-term survival of the animals from 0 to 70%. 50 These evidences indicate that inflammatory cells and their secreted factors may have influence on tissue replacement after injury through stem cells and other reparative mechanisms, as summarized in Figure 2 51 and Table 1. 52
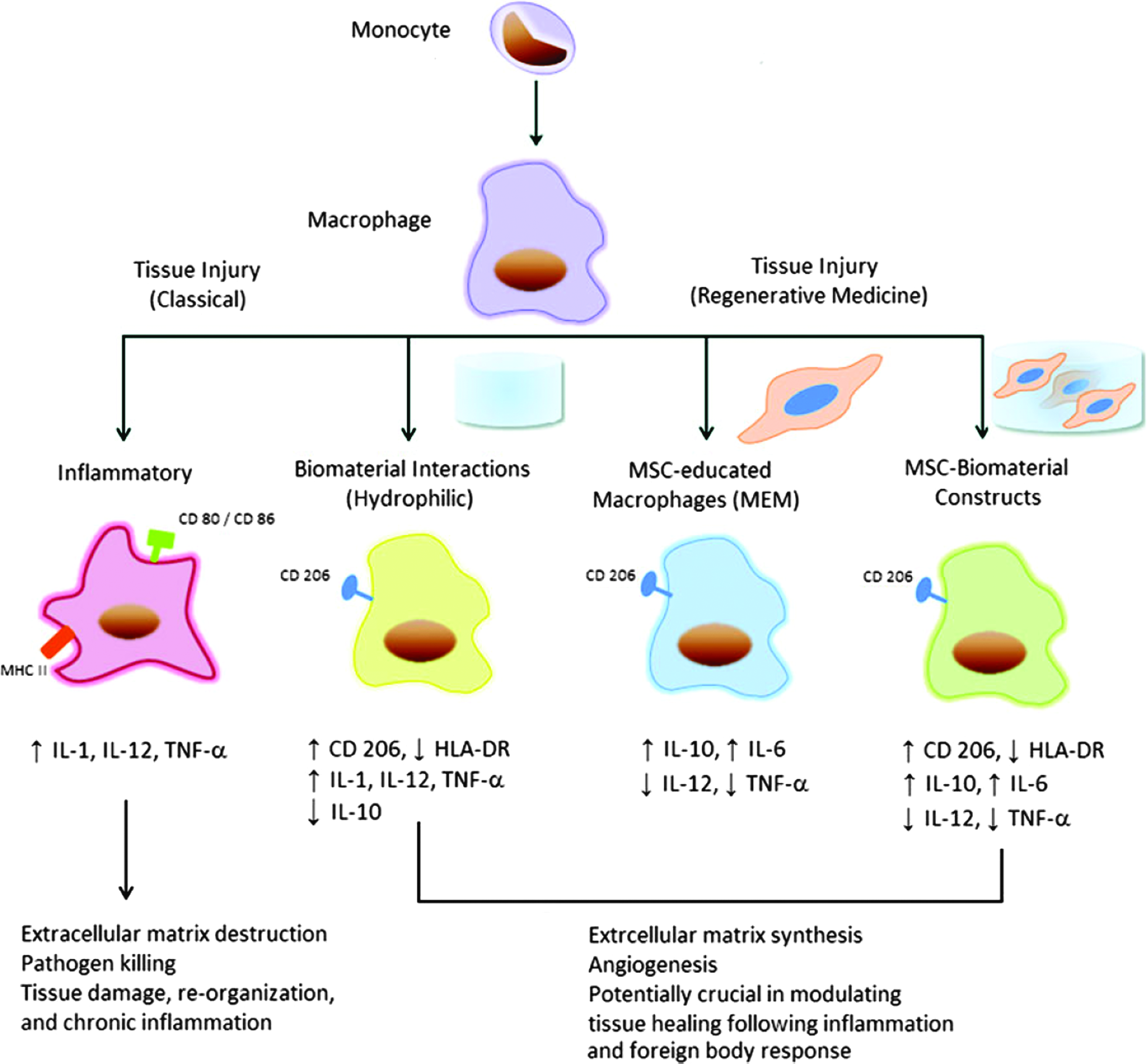
Illustration of the immunomodulatory effects of biomaterials and mesenchymal stem cells (MSCs) on macrophage phenotype and function in vitro. Upon stimulation such as tissue injury, macrophages can acquire a classically activated phenotype, which is associated with proinflammatory cytokines (i.e., IL-1, IL-12, tumor necrosis factor-α [TNF-α]), debris removal, continued inflammation, and tissue injury. These M1 macrophages bear surface markers such as CD80, CD86, and MHC class II molecules. Alternatively, there is evidence to support an alternatively activated anti-inflammatory immunophenotype, which can be acquired when monocytes are cultured on biomaterials with specific surface chemistry (i.e., hydrophilic) or cocultured with MSCs, with or without a biomaterial scaffold. These alternatively activated macrophages, M2, display surface markers such as CD206 and are associated with an increase in anti-inflammatory cytokines (i.e., IL-10, IL-6, and TGF-b), extracellular matrix production, and tissue remodeling. IL, interleukin; TGF-β, transforming growth factor-β (permitted reprinting from Hanson et al. 51 ). Color images available online at www.liebertpub.com/teb
The process of regeneration is summarized for six different tissues, each with varying regenerative capacities: skin, heart, skeletal muscle, CNS, liver, and bone. For each, steps that have the potential to be influenced by immunity are listed as well as the disease state and summary of immune-targeted therapeutics (modified from Table 3 of Aurora and Olson 52 with permission).
CNS, central nervous system; EGF, epidermal growth factor; IFN, interferon; IGF, insulin-like growth factor; IL, interleukin; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-α; VEGF, vascular endothelial growth factor; n/a, not applicable.
Conclusion Remark
This article serves as an overview of basic research advancement in TERM field with particular focus on stem cell-related subjects and also immune modulation on stem cell function. The impressive achievements can be summarized in these aspects. (1) Significant advancements were achieved in the area of iPSCs, in which various types of regenerative cells were able to be derived from iPSCs with their regenerative functional confirmation both in vitro and in vivo. Many new tissue-specific stem cells/progenitor cells were discovered as well and impressive progress was also made in SCNT to generate hPSCs. (2) New concepts were also proposed and proven, such as regulation of stem cell behavior through sexual hormones and dietary/metabolic controls; immune modulation of stem cell and tissue regeneration and nonstem cell-mediated tissue regeneration. (3) Application of genomic editing to stem cell biology, which is likely to correct genomic disorder of the stem cells derived from a patient with genetic disease. All these achievements will provide a solid scientific foundation for future translation work in TERM field.
Footnotes
Acknowledgment
This was supported by the National Natural Science Foundation (31170937, 81071590).
Disclosure Statement
No competing financial interests exist.