
Abstract
The lymphatic system is involved in maintaining interstitial fluid homeostasis, fat absorption, and immune surveillance. Dysfunction of lymphatic fluid uptake can lead to lymphedema. Worldwide up to 250 million people are estimated to suffer from this disfiguring and disabling disease, which places a strain on the healthcare system as well as on the affected patients. The severity of lymphatic diseases calls for the establishment of new treatment methods. One approach is to replace dysfunctional lymphatic vessels with bioengineered ones. In this study, we mainly focus on hydrogels, scaffolds with cellular constructs, interstitial flow, and extracorporeal shockwave therapy. This review provides an overview on the current status of lymphatic biology and approaches of reconstruction and regeneration of lymphatic vascular tissues.
Introduction
M
Historically, the first discovery of lymphatic vessels was made by Nicolas Massa of Venice (1531–1569) and documented by Bartolomeo Eustachius in 1563. However, the function of these vessels remained unclear. The first differentiation between veins and lymphatics (chyliferous vessels) was documented for dogs by Gasparo Asselli in 1622. Lymphatic vessels were first discovered in humans and illustrated by Johann Vesling 2 years later. Olof Rudbeck clarified that the lymphatic network is a system with vessels throughout the body similar to the blood vascular system and discovered the fluid uptake by these vessels. 1 Despite these initial observations, the molecular mechanisms of the lymphatic system remained unclear until the last two decades due to the lack of reliable markers and experimental models.2,3 Research in lymphatic biology increased rapidly after identifying specific lymphatic markers, such as vascular endothelial growth factor receptor (VEGFR)-3, 4 podoplanin, 5 prospero-related homeobox protein 1 (Prox1) 6 and lymphatic vessel endothelial hyaluronan receptor (LYVE)-1. 7
The lymphatic vascular system plays a crucial role in regulating the interstitial fluid homeostasis, in absorbing dietary fats and in providing a conduit for immune cell trafficking. 8 This system is found in nearly all vascularized tissues, but was thought until recently to be absent in the central nervous system. In 2012, however, Iliff et al. revealed glial lymphatics in the dura mater of a mouse brain, shortly called “glymphatics”.9,10 Glymphatics absorb brain interstitial fluid and dural lymphatic vessels absorb the cerebrospinal fluid (CSF) from the adjacent subarachnoid space.11,12 The CSF drains into extracranial lymphatic vessels and lymph nodes. 13 Aside from disruption of lymphatic vessels, an impairment of the fluid uptake capacity can be due to increased vascular leakage in inflammation or injury leading to lymph accumulations within the interstitial space, namely lymphedema. 14 Lymphedema affects between 140 to 250 million people worldwide. 15 The globally most prevalent cause is the parasitic disease filariasis, which is common in South-East Asia. However, in many developed countries the major cause of disruption of the lymphatic pathways is the treatment of malignancies by surgery and/or radiation. Mainly, women with breast cancer are affected. 15 Patients with such postsurgical edema suffer from chronic and progressive swelling, recurrent infections, pain and significantly decreased quality of life. The currently available treatments include lymph drainage, compression garments, or microsurgery.16,17 Although these modalities can reduce the edema volume and prevent new fluid accumulation, patients require life-long treatment, challenging the healthcare system as well as the affected individuals and their families. 18 One approach to replace dysfunctional lymphatic vessels is microsurgical lymphatic vessel transplantation. Historically, transplantations were first performed by lymph–venous anastomoses.19–21 Nowadays, it is possible to harvest lymphatic grafts from the patient (e.g., anteromedial bundle at the inner aspect of the thigh) and anastomose these with lymphatic vessels at the defect site. Injury of the major (thoracic) duct can occur during surgical interventions and needs immediate surgical ligation. In case of extensive damage, replacement of this region might be preferable through anastomoses. However, due to redundancy in its conduit the need for thoracic duct replacement is small. The thin and fragile walls of the lymphatic vessels make anastomoses difficult.22,23 Moreover, three-dimensional (3D) constructs containing lymphatic vessels could be used for drug development or as a potential in vitro disease model for lymphedema and lymphoid hyperplasia. 24 A future-oriented approach is to create bioengineered lymphatic vessels. For instance, lymphatic endothelial cells (LECs) can be embedded into a 3D matrix to create lymphatic capillaries, and their proliferation can be increased by applying interstitial flow, using prolymphangiogenic growth factors or adapting shockwave therapy. These hydrogels could be implanted in vivo to improve wound healing and enhance lymphatic ingrowth after plastic surgery (e.g., skin flaps). This review summarizes current in vitro and in vivo approaches in lymphatic tissue engineering and the regeneration of lymphatic vessels.
Biology of the Lymphatic System
The lymphatic system (Fig. 1) consists of central/primary lymphoid organs, such as bone marrow and thymus, and the peripheral/secondary lymphoid organs, including lymphatic vessels, lymph nodes, spleen, adenoids, Peyer's patches, appendix and mucosa-, gut- and bronchial-associated lymphoid tissue. 25 The high pressure within the blood vascular system generates interstitial fluid by capillary filtration and the lymphatic system serves as a one-way drainage system for this intercellular surplus fluid. The interstitial fluid contains macromolecules, certain leukocytes, dissolved solutes, viruses, bacteria, and cell debris. 8 After it has been taken up by the lymphatic capillaries it is shortly called lymph. In contrast to blood, lymphatic capillaries are thin walled and have a relatively wide lumen (Fig. 2). 26 Moreover, the lymphatic capillaries are blind ended and consist of oak leaf-shaped endothelial cells (ECs), which lack a continuous basal lamina. 27 Discontinuous button-like junctions connect these cells and anchor the flap valve forming overlapping cell edges,28,29 through which the interstitial fluid enters the lymphatic vessels following the pressure gradient. 30 When the volume of the extracellular matrix (ECM) increases, the overlapping LEC junctions open. These overlaps act as one-way valves and prevent the fluid from returning into the surrounding tissue. Lymphatic capillaries lack smooth muscle cells (SMCs) and are connected to the ECM through anchoring filaments. These filaments get pulled apart when the interstitial volume increases assisting thereby in the valve opening.29,31
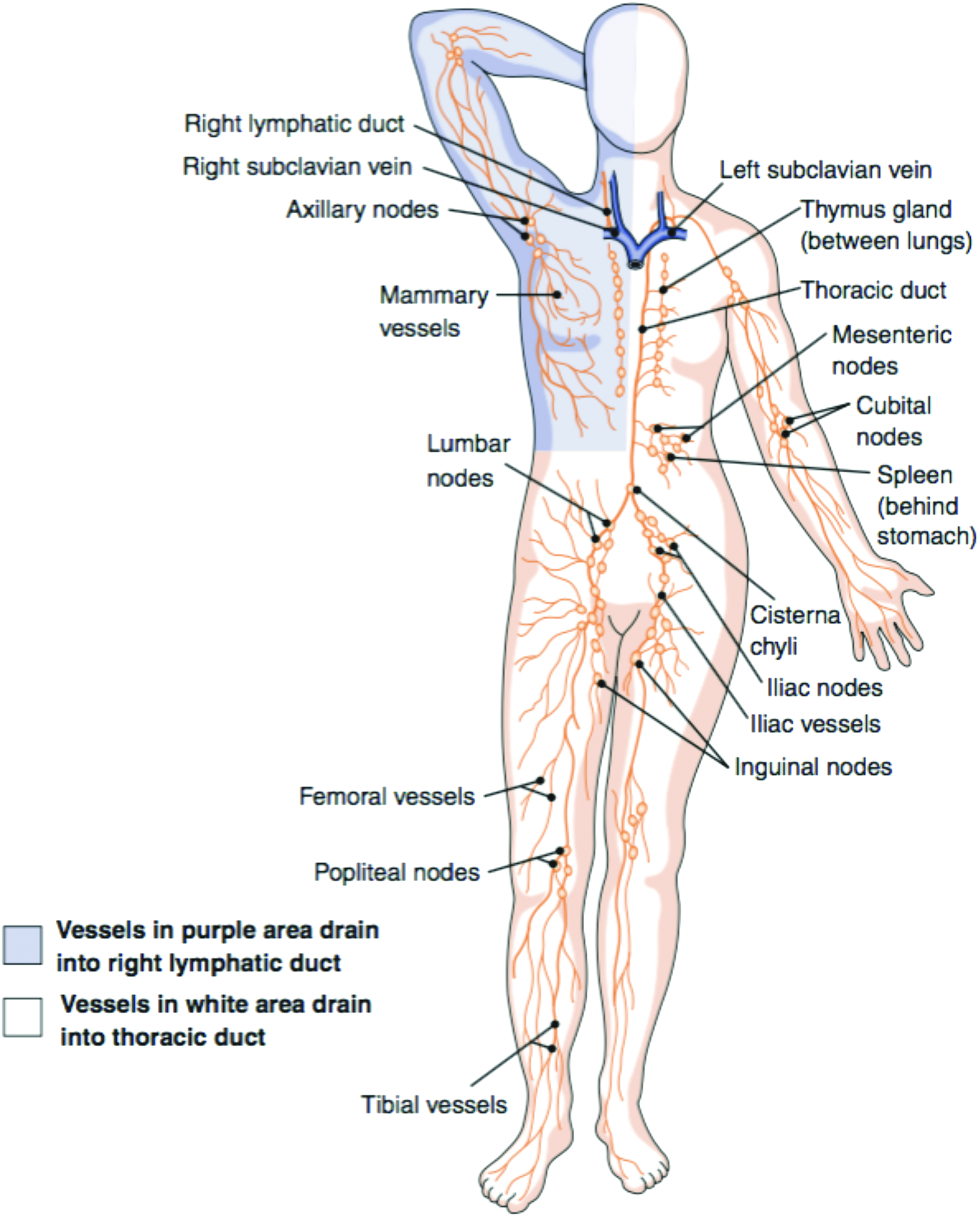
The lymphatic system in the human body. The lymph is pulled from collecting vessels into the five trunks, which drain the lymph into the ducts. Lymphatic vessels in the purple area drain into the right, and lymphatic vessels from the white area into the thoracic duct. Finally, the ducts drain the lymph into the venous circulation through the subclavian vein. 146 Color images available online at www.liebertpub.com/teb
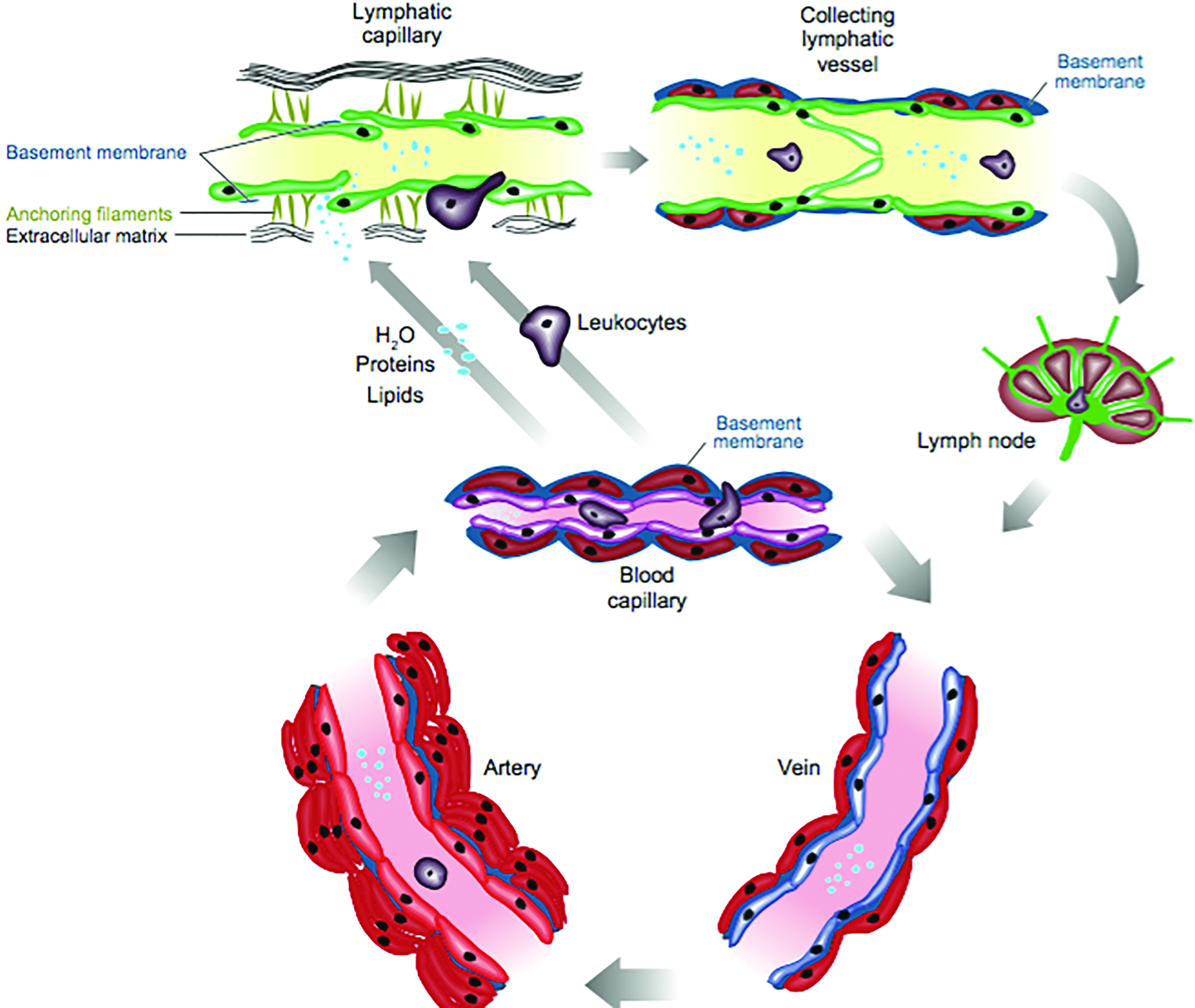
Structure of lymphatic vessels. Lymphatic capillaries (green) are thin walled, blind ended, and have a relatively wide lumen compared with blood vessels. Instead of having tight junction LECs partly overlap resulting in valve-like openings, through which interstitial fluid can enter the lymphatic vessels' lumen. These act as one-valve system preventing leakage of lymph into the surrounding tissue. Lymphatic capillaries lack SMC and are connected through anchoring filaments to the ECM. Increased interstitial volume pulls these filaments apart and consequently supports opening of the valves. After the lymph enters the capillaries, it is further transported to collecting lymphatic vessels and trunks. Finally, lymph drains into the ducts, which return the lymph back in the blood system. Collecting lymphatic vessels have a basement membrane, valves, and are surrounded by SMC (red) that promote lymph flow. In contrast, blood vessels have tight and adherence junctions, distinct basement membrane and multiple layers of SMCs. 26 ECM, extracellular matrix; LEC, lymphatic endothelial cell; SMC, smooth muscle cell. Color images available online at www.liebertpub.com/teb
In the small intestine of the digestive tract specialized lymphatic capillaries (lacteals) are responsible for transporting lipids. 32 However, the mechanisms regulating the uptake are still unclear.
After drainage into precollecting lymphatic vessels the lymph enters collecting lymphatic vessels. These are covered with a layer of SMC surrounding the endothelial layer that is connected with continuous zipper-like junctions,27,30 which prevent the leakage of lymph. Unidirectional flow in collecting vessels is ensured by intraluminal valves. From the collecting vessels the lymph is cleared in lymph nodes and further transported through lymphatic trunks to lymphatic ducts. 33 In total, there are four pairs of lymphatic trunks (lumbar, bronchomediastinal, subclavian, and jugular trunks) and one unpaired (interstinal trunk). All of these drain their content into two large collecting vessels called lymphatic ducts and these return the lymph back into the blood circulatory system. The accumulated lymph from the right side of the body superior to the diaphragm flows toward the right lymphatic duct and drains the lymph into the right subclavian vein. The larger, left lymphatic duct collects lymph from both sides of the body inferior to the diaphragm and from the left side of the body superior to the diaphragm. It returns the collected lymph into the left subclavian vein.33,34 In healthy adults, around 2–4 L of interstitial fluid with a protein content of 20–30 g/L return through the lymphatic system into the venous vasculature every day.2,8 This covers approximately 10% of the liquid that extravagates from the blood vessels into the interstitial space. The other 90% are reabsorbed by the venous capillary system. 2 Lymph transport from the initial capillaries into the collecting vessels is based on systemic forces (blood pressure, respiratory motion massage, peristaltic movement, and contractility of surrounding skeletal muscle) and interstitial fluid pressure.35–40 Furthermore, spontaneous contraction of SMCs and valves is crucial for lymph transport through collecting vessels. 40 In contrast to mammals, amphibians and reptiles feature additionally specialized pulsatile muscular organs (lymph hearts), which aid in the active lymph propulsion.41,42 This complex vascular architecture has to be taken into consideration for any attempt to create tissue-engineered lymphatic vessels.
Lymphangiogenesis
The development of the lymphatic system was described for the first time in 1902 by Florence Sabin, who suggested that ECs bud off from embryonic veins and form central primitive sacs from which ECs sprout out into the periphery by angiogenic mechanisms. 43 The concept of Sabin has been supported by an analysis of genetically altered mice.44–46 Recent data from Oliver et al. and Srinivasan et al. support the transdifferentiation of venous ECs into LECs as origin of the lymphatic system in mammals.47,48 However, this centrifugal model was early on challenged by Huntington (1908) and Kampmeier (1912), who alternatively suggested a centripetal model, where LECs arise de novo in the mesenchyme from precursor cells (lymphangioblasts).49–51 Recently, such contributions to the LEC population have been shown for specific organs and vascular beds: Nicenboim et al. proposed that LEC progenitors can arise from specialized mesoderm-derived angioblasts located within the venous niche, 52 Stanczuk et al. suggest that mesenteric lymphatic vessels develop from the c-Kit lineage cells of hemogenic endothelial origin, 53 and Yang et al. suggested that intersomitic vessels (ISVs) are an additional source of LECs in mice. 43 Hence, settling the century-old controversy between Sabin's and Huntington's models seems within reach. Both mechanisms are contributing to the embryonic lymphatic development in vertebrates; however, their relative individual contributions seem to be variable in different species. 54 The formation of the first lymphatic vessels starts in human in utero at around week 6–7 (mouse embryonic day (E) 9.5) when the blood circulatory system is already fully established and functional (Fig. 3a).2,55 Lymphangiogenesis requires proliferation, sprouting, migration, and tube formation. 56 The earliest evidence of lymphatic differentiation is the expression of the sex-determining region on the Y chromosome-related high-mobility group box group-F (SOXF) family transcription factor SOX18 in a subpopulation of blood vascular endothelial cells (BECs) in the dorsolateral wall of the cardinal vein (CV). 57 SOX18 activates Prox1, which induces the emigration of LEC progenitors from the veins in a polarized manner.2,43 This process requires the prolymphangiogenic factor vascular growth factor (VEGF)-C.44,58 At approximately E10.5, the venous-derived LECs merge together with LECs that budded off from ISVs. At this stage, LECs do not yet express podoplanin. Podoplanin is only detected after LECs have fully exited from the CV or ISVs. The fact that, regardless of their origin, both Prox1-expressing LEC populations become podoplanin-positive suggests that they have the same molecular identity and that the emigration from the veins is linked with the LEC differentiation program.43,59 Cytoplasmic tyrosine kinase (Syk)-expressing myeloid cells potentiate lymphangiogenesis by producing chemokines and growth factors, including VEGF-C and VEGF-D.60,61 In the separation process of blood and lymph vasculature, platelets play a crucial role and require the Syk/SLP-76/PLCγ signaling pathway in hematopoietic cells.62–64 This pathway is activated in platelets by the transmembrane podoplanin receptor C-type lectin receptor (CLEC)-2 and induces platelet aggregation at the adhesion junctions between developing lymph sacs and CV,61,65–69 thereby closing off the primary connection between blood and lymphatic systems. 65 The emergence of the intraluminal valves in the collecting vessels depends on forkhead box protein C2 (Foxc2) expression in prospective valve-forming cells (Fig. 3b). 70 Foxc2 downregulates the expression of lymphatic markers such as Prox1, VEGFR-3, or LYVE-1.2,5–7,71 Once the primitive lymphatic networks are formed, remodeling and specialization leads to the development of two predominate types of vessels: small lymph capillaries and larger collecting vessels. 2 In adults, under normal physiological conditions, lymphangiogenesis occurs only in the corpus luteum formation during menstruation and in the endometrium during pregnancy.72,73 Finally, lymph nodes appear at E12.5 and the development is driven by the tumor necrosis factor family cytokine TRANCE and interleukin 7 (IL-7) (Fig. 3c). 14
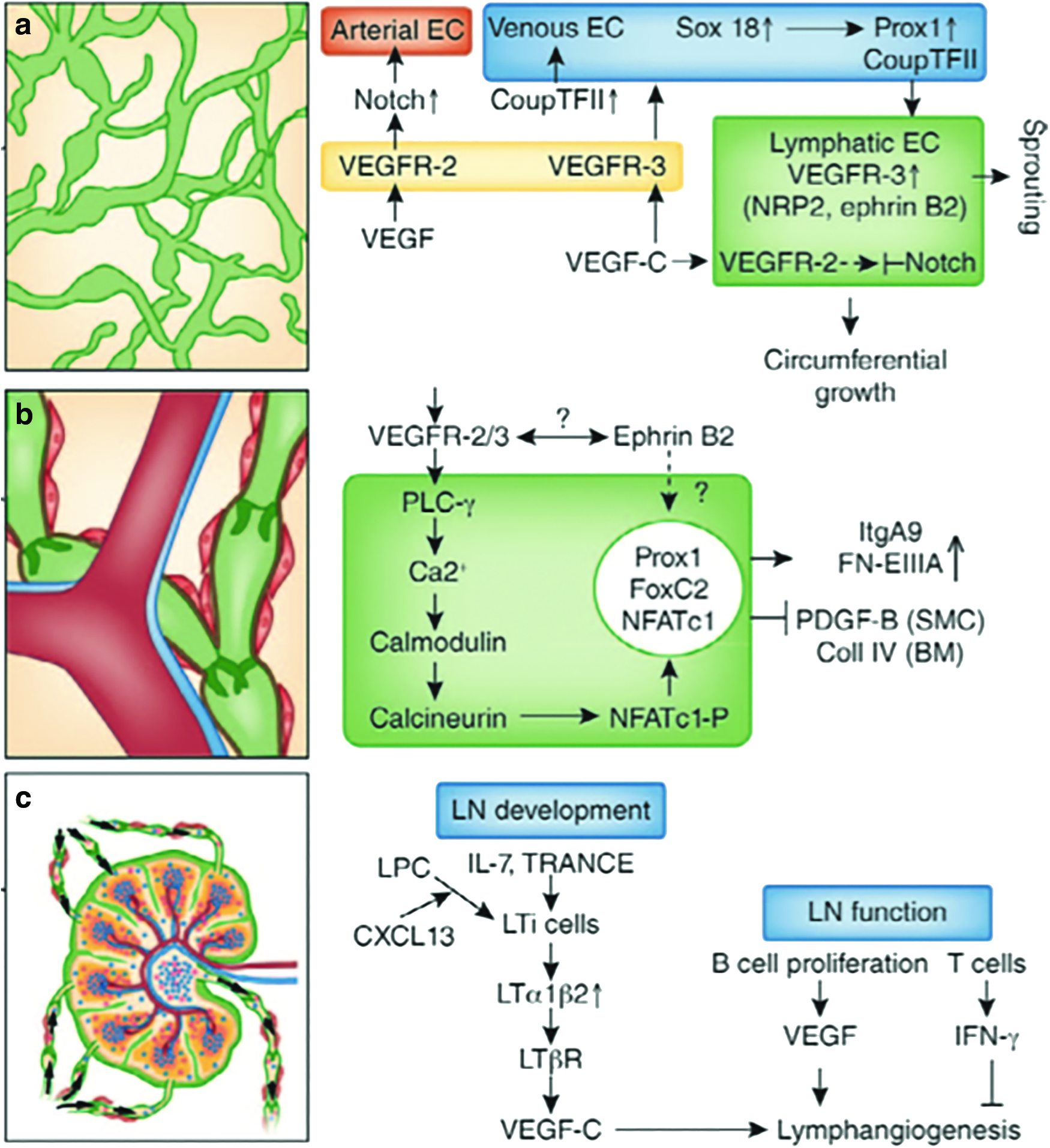
The lymphatic architecture and signals involved in the formation of
LECs and Specific Lymphatic Endothelial Markers
LECs express pan-endothelial markers such as platelet endothelial cell adhesion molecule(PECAM)-1, also known as CD31, VEGFR-2, and specific lymphatic markers, including Prox1, podoplanin, LYVE-1, VEGFR-3, and chemokine C-C motif ligand (CCL)-21. 3 Prox1 determines lymphatic identity and induces ECs with lymphatic fate to bud from the CV to give rise to the lymphatic system.6,74 In Prox1 null mice, venous ECs cannot successfully differentiate into LECs and all subsequent lymphatic development is arrested by E11.5-E.12, leading to a lack of the lymphatic system.74,75 The fms-like tyrosine kinase VEGFR-3, also called Flt-4, is important for lymphatic development and serves as receptor for VEGF-C and VEGF-D. VEGFR-3 is expressed on all ECs during the early stages of embryonic development, however is restricted to LECs in the adult organism.4,76 Deletion of VEGF-C in mouse embryos leads to a complete absence of the lymphatic vasculature. LECs do differentiate in VEGF-C null mice, but fail to emigrate from the CV. 77 After the budding of LEC progenitor cells has completed by E11.5 in mice, they express high levels of podoplanin.2,5 Podoplanin-deficient mice display lymphatic defects and die shortly after birth due to respiratory failure. 44 Podoplanin can be used to distinguish LECs and BECs, because it is never expressed by BECs. Podoplanin can bind to CLEC-2, which is highly expressed by immune cells and platelets. 78 The binding of podoplanin to CLEC-2 increases platelet aggregation and activation as well as motility and protrusion formation in immune cells. 78 The interaction of CLEC-2 and podoplanin is crucial to lymphatic vessel maintenance.67,68 Another predominant cell surface receptor that is almost exclusively confined to lymphatics and lymph node endothelium is LYVE-1. It plays a role in lymphatic intercellular junctions and is not only expressed on lymphatic vessels, but also by liver cells and in the spleen.79,80
Coculture of LECs with ASCs or fibroblasts lead to lymphatic capillary-like structures that can be enhanced by applying interstitial flow and/or adding growth factors, such as VEGF-C. Moreover, the choice of matrix is crucial. Regeneration and reduction of lymphedema can be achieved through gene therapy and shockwave treatment.
ASC, adipose tissue-derived stem cell; BEC, blood vascular endothelial cell; HDMEC, human dermal microvascular endothelial cell; ESWT, extracorporeal shockwave therapy; IGF, insulin-like growth factor; LEC, lymphatic endothelial cell; LV, lymphatic vessels; HGF, hepatocyte growth factor; FGF, fibroblast growth factor; PGA, polyglycolic acid; TE, tissue engineering; VEGF, vascular endothelial growth factor.
Sources of LECs and In Vitro Culturing
For lymphatic tissue engineering the key prerequisite is the availability of LECs. Until recently, the isolation of LECs from different sources, such as dermis, intestine, and lymph nodes, was the only option.81,82 However, a recent study describes the differentiation of induced pluripotent stem cells (iPSCs) into the lymphatic lineage. 83 Garrafa et al. isolated LECs from tissues such as lymph nodes, thymus, spleen, palatine tonsils, and iliac lymphatic vessels, using first Ulex Europaeus Agglutinin 1 (UEA I)-coated beads to isolate ECs followed by a purification step using a monoclonal anti-podoplanin antibody D2-40.84–86 For the isolation of LECs from dermis, skin can be obtained from adults who underwent elective surgery (abdominoplasty or breast reduction). After the epidermis has been mechanically removed, human dermal microvascular endothelial cells (HDMECs) are retrieved by enzymatic digestion. Thereafter, the dermal cell suspension was subjected to fluorescence-activated cell sorting to obtain podoplanin+CD34+CD45− cells. 86 Another possible source to obtain LECs from human individuals and mice are the lymph nodes. First, the lymph nodes are disrupted with a needle and then enzymatic digested. Afterward, LECs are purified by podoplanin, CD31, and LYVE-1 positivity.87–89 Yet, another source for LECs is the intestine. The mucosa from the jejunum is enzymatically digested to harvest interstitial microvascular cells for further isolation,90,91 which are then seeded on fibronectin-coated plates. Transcriptomic analysis of the cells revealed that the gene expression of HDMECs and interstitial LECs (iLECs) is similar. 92 Since LECs are generally plastic compared with other cell lines, it is expected that they display a certain degree of heterogeneity. However, there is yet not enough data to clarify whether LECs derived from certain tissue can be used to create artificial lymphatic vessels for other tissues.
Freshly isolated cells are often grown in fibronectin- or collagen-coated flasks.93,94 However, coating appears not to be necessary for further culture. 94 Many variants of culture medium are possible as long as EC growth medium is used.44,95–97 Growth of LECs have been reported to be stimulated by growth factors, including VEGF-C, VEGF-A, human fibroblast growth factor (hFGF)-B, FGF-2, epidermal- and insulin-like growth factors (EGF and IGF)-1, IGF-2, hepatocyte growth factor (HGF), and platelet-derived growth factor-BB.97–103 Primary cells have a limited growth potential. Thus, to keep them proliferating for longer periods, they can be for example, infected with a retrovirus, which contains a coding region for human telomerase reverse transcriptase.104,105 Such cells can retain the typical cobblestone morphology and lymphatic endothelial marker expression beyond 40 passages. 104 Taken together, there are approaches available to obtain adequate cell number, including iPSC reprogrammed LECs and telomerase-immortalized LECs; however these are limited by the lack of knowledge of long-term consequences as well as safety issues.
Engineering Lymphatic Vessels
In contrast to the generation of lymphatic vessels, the engineering of blood vessels has been understood for several years, for example, from results obtained by coculturing of different EC types with mesenchymal stem cells, which results in branching, long tubes, and capillary-like blood vessels.106,107 The advances made in tissue-engineered blood vessels through scaffold (e.g., acellular scaffolds incorporated with biomolecules) and scaffold-free techniques can guide the engineering efforts to generate large lymphatic vessels. Scaffold-free approaches use coculture systems, decellularization methods, cell sheet engineering, fluid shear stress through bioreactor, and bioprinting.108–110 Over the last decades and after identifying the main lymphatic markers, research in the lymphatic field has gained a boost and promising approaches have been developed. Advances in tissue engineering have been made in bone and avascular tissue, such as cartilage.96,111–113 However, Marino et al. were able to engineer prevascularized dermoepidermal human skin grafts containing both human blood and lymphatic capillaries. 114 For this purpose, HDMECs (which consist of both BECs and LECs) were cocultured with fibroblasts in fibrin hydrogels for 3 weeks to achieve a complete vascular plexus in vitro. Fibroblasts were a prerequisite for the successful development of bioengineered human lumen-forming lymphatic capillaries in both fibrin and type I collagen hydrogel in these experiments. Lymphatic and blood vascular capillaries never anastomosed, and in vivo the lymphatic capillaries were able to drain injected Evans Blue dye, thereby, indicating perfusion. The skin graft, a fibrin hydrogel containing HDMECs, fibroblasts, and keratinocytes, was transplanted into rats, where the bioengineered lymph and blood capillaries maintained their lumen and established connections to the host rat vessels. 114 However, although significant progress has been achieved, the vascularization of larger tissues and 3D models is still a problem and remains challenging.
Approaches for Scaffold-Based Tissue Engineering of Lymphatic Vessels
In the field of tissue engineering, the last decade has seen tremendous technological advancements. With the exception of the lymphatic system, nearly every major organ has been targeted, including cardiac, cartilage, nervous, bone, lung, liver, and kidney tissue.115–121 Nevertheless, inclusion of lymphatics and blood system in these organs is necessary for successfully in vivo application.117,122 One of the fundamental challenges in creating tissue-engineered lymphatic vessels is to replicate the morphology of the cell structure and its organization. Few attempts have been made to generate lymphatic vessels in 3D structures.96,123 One approach uses polyglycolic acid (PGA) tubes as matrix, which were successfully implanted into mice after seeding with LECs. However, instead of establishing tube-like structures, LECs formed sheets around the PGA tubes. 124 Another method cocultures LECs onto a feeder sheet of fibroblasts. These constructs spontaneously organized into a stable 3D lymphatic capillary network without requiring any exogenous biomaterials or growth factors; LEC proliferation and tube formation are induced by fibroblast-derived VEGF–C and HGF. 125 Helm et al. established a 3D VEGF-fibrin-collagen matrix construct and applied interstitial flow. 126 In this study, BECs and LECs were cultured in an interstitial flow culture chamber using different setups.79,126 The data suggested that for assembly soft matrices are preferred by both BECs and LECs. In contrast to BECs, LECs preferred less permeable matrices. Thus, LECs organized more efficiently in fibrin-only matrices and emerged as long and overlapping structures, whereas BECs preferred an equal mixture of fibrin and collagen and formed densely branched tubes that had a wider lumen than lymphatic vessels. 126 These studies demonstrated the importance of 3D matrix composition and interstitial flow for in vitro vascularization. Moreover, the used multichamber fluidic device allows simultaneous 3D fluidic experiments for long-term culture. The lymphatic system is required for clearance of interstitial fluid and is therefore constantly exposed to and stimulated by fluid flow and pressure. Interstitial flow can affect capillary morphogenesis, fibroblast remodeling of the ECM, and tumor cell migration. 95 Boardman and Swartz hypothesized that interstitial fluid flow may play a crucial role in lymphangiogenesis. They investigated the correlation of interstitial fluid flow and LEC migration. A 2 mm circumferential band of dermal tissue was removed from a mouse tail and an acellular collagen dermal equivalent (CDE) was inserted into the gap through which the lymph moves in a proximal direction. By creating this collagen window, an observation of lymphatic architecture and evaluation of tissue fluid balance parameters has been enabled. The removal of the dermal tissue band did not destroy the underlying major blood vessels, muscles, or bone. Thus, only the lymphatic flow was affected and resulted in edema of the tail without a fluid-bridging treatment such as the CDE. 127 The authors evaluated the lymphangiogenic processes with cellular, molecular, and fluid channel markers over a period of 60 days. In the initial phase of 10 days, the fluid moves diffusely and without channeling through the CDE and large amount of VEGF-C was seen at the distal edge of the CDE. After 25 days, crude but distinct LYVE-1 and VEGFR-3-positive fluid channels could be observed that further assembled into an intact, continuous lymphatic network after 60 days. This network of functional lymphatic vessels assembled from single cells detected primarily within the CDE on the distal site. The expression of VEGF-C and the sprouting of LECs correlated with the direction of the interstitial flow. Interestingly, VEGF-C expression was barely detectable after 25 days and after 60 days not detectable at all. 127 In another model system, the significant effect of interstitial flow on lymphangiogenesis and angiogenesis was demonstrated by using a multichamber fluidic device for 3D culture. This fluidic 9-chamber radial flow device allows long-term culture with live imaging capabilities, and due to the multiple chambers, different experiments can be performed simultaneously. In this device the LEC cultures that were exposed to flow spread and formed multicellular, lumenized structures similar to natural lymphatic networks, and the vessel density and organization of these structures could be improved by adding an engineered variant of VEGF that bind fibrin through factor XIII. 95 In addition, Ng et al. demonstrate that LECs form large vacuoles and long extensions under interstitial flow. 128 Taken together, several tissue engineering approaches for lymphatics have been reported, yet further research is needed in this field.
Regenerative Approaches
In addition to several in vitro approaches for the construction of a lymphatic vasculature, the regeneration of damaged lymphatics in situ has gained more interest recently. Regeneration of lymphatic networks and reduction in edema can be achieved by VEGF-C gene therapy.44,129–133 Transient overexpression of VEGF-C leads to a functional lymphatic vessels network with valves and SMC coverage, due to induced growth, differentiation, and maturation. 131 Moreover, the calcium-binding epidermal growth factor domain 1 (CCBE1) protein enhances lymphangiogenesis by promoting VEGF-C activation through proteolytic cleavage. These results suggest CCBE1 as potential therapeutic candidate for inducing lymphangiogenesis. 134
Regeneration of lymphatic vessels has been demonstrated recently by using physical treatment methods such as extracorporeal shockwave therapy (ESWT). This method has been originally used for the removal of kidney stones. Recently, in addition, beneficial effects on various musculoskeletal pathologies, including stress fractures, nonunion fractures, and chronic tendinopathy have been described. 135 EWST leads to increased cell permeability and expression of growth factors, such as VEGF-C, which is associated with promoting lymphangiogenesis and accelerating capillary morphogenesis.126,136
ESWT-induced promotion of lymphangiogenesis was shown by Kubo et al. in a rabbit model. Disrupted, edema-causing lymphatic vessels within the ear were treated with or without low-energy shockwaves. Significantly enhanced expression of VEGF-C and VEGFR-3was detected in the treated ears. Moreover, the lymphedema thickness was reduced. 137 In addition, ESWT was tested in a tail model of lymphedema in rats. Low-energy ESWT led to an increased expression of VEGF-C and basic FGF (bFGF) and decreased the lymphedema. 138 Kim et al. used mice suffering from lymphedema at the mid-thigh to investigate the effect of gelatine hydrogels containing VEGF-C and/or ESWT. The combination of treatment methods (VEGF-C hydrogels and ESWT) showed the best results in terms of lymphatic vessel formation, improvement of lymphedema, and enhanced expression of VEGF-C and VEGFR-3. 139 In this context, it is important to note that VEGF-C concentrations well beyond physiological levels induce lymphatic hyperplasia while inhibiting an increase of lymphatic vessel density.140,141 In another study, Rohringer et al. investigated the effects of ESWT on LECs regarding morphology, migration, proliferation, marker profiles, and gene expression. The results revealed a positive effect of ESWT on LEC proliferation in vitro, which was critically dependent on the energy flux. By coculturing LECs with adipose tissue-derived stem cells in fibrin hydrogels and consecutive treatments with ESWT, the formation of tube-like structures was enhanced. Podoplanin was significantly upregulated by treated cells and biological properties were altered in terms of proliferation, migration, morphology, marker profiles, and gene expression. 142 To conclude, gene therapy and ESWT enhance LEC proliferation and regeneration of the lymphatic network.
The thorough evaluation of engineered lymphatic tissue constructs critically depends on accurate and robust methods for the detection and visualization of lymphatic vessels, for example, a new model using transgenic Prox1-Cre-tdTomato reporter mice. 143 This reporter is well suited for in vivo imaging as it shows bright red fluorescence in lymphatic vessels allowing deep imaging. In comparisons, tdTomato is more photostable than tandem-dimer red fluorescent protein and leads to less photobleaching during intravital microscopy (IVM). Other Cre-lines that have been previously used revealed Cre expression in some non-LECs, for example, LYVE-1-Cre identifies a subset of leukocytes and BECs, 144 and in experiments with podoplanin-Cre lines, expression could be seen in secondary lymphoid organs as well as in stromal cells of lymph nodes. 145 Therefore, the use of LYVE-1-Cre- or podoplanin-Cre-expressing mouse lines is only of limited value for lymphatic vessel imaging. A further advantage of the new Prox1-Cre-tdTomato reporter is the possibility of direct monitoring and long-term imaging by IVM as well as repeated imaging of the same mouse. Thus, morphological changes can be visualized, while reducing the amount of animals used for the study. Using this Prox1-Cre-tdTomato mouse strain, Roberta Bianchi et al. demonstrated that dendritic cells (DCs) can actively migrate inside lymphatic vessels after entry, as opposed to the older view that DCs get passively transported toward lymph nodes after lymphatic vessel entry. 143 Hence, this reporter mouse provides an accurate tool for visualization of lymphatic vessels.
Conclusion
The presence of lymphatic vessels in nearly all organs and the fact that they are essential for human survival clearly demonstrates the necessity of their integration in tissue engineering approaches in addition to the blood vascular system. Since scientific attention on the molecular basics of lymphatics was focused yet roughly in the last two decades, unsurprisingly, the development of novel effective therapies to regenerate or replace lymphatic vessels still lags behind. Learning from blood vascular tissue engineering one can partly translate this knowledge to the lymphatic counterpart. Nevertheless, the unique features and characteristics of lymphatic vessels (e.g., the unidirectional flow, the special valves, or the microarchitecture of the lymphatic microcapillaries) demand for special approaches to accompany blood vascular with lymphatic tissue engineering. Based on the availability of solid data on lymphatic molecular biology on the one hand, and on the more and more sophisticated methods to engineer tissues on the other hand, the development of strategies which reflect the intimate relationship of the blood vascular and lymphatic systems will arise soon.
Footnotes
Acknowledgments
The authors thank Simon Sperger, Severin Mühleder, and Carina Egger for critical reading of the article.
Disclosure Statement
No competing financial interests exist.