
Abstract
In vitro culture of ovarian follicles is a promising bioengineering technique for preserving fecundity in reproductive-aged female by providing fertilizable oocytes. Successful clinical application should be preceded by developing the protocols that can efficiently overcome follicular cell apoptosis since the apoptosis is a critical phenomenon in in vivo folliculogenesis and in in vitro follicular maturation. Numerous prosurvival and antiapoptotic molecules, including follicular developmental regulators, have been reported to be involved in the intraovarian apoptosis. The authors searched literature and analyzed the current knowledge of these proteins and noncoding RNAs, and their antiapoptotic roles in the dynamics of follicular development in vivo and in vitro. Two-dimensional (2D) culture method has widely been used, however, with recent emergence of various biomaterials, three-dimensional (3D) culture is also considered a proper environment for maintenance of solid structure of ovarian follicles. The identification of candidate paracrine and endocrine intracellular effectors that are responsible for the coordination occurring between oocyte, granulosa, and theca cells during follicular development was explored in this review, to assess the possibility of their use as antiapoptotic factors in establishing more efficacious 2D or 3D in vitro follicular microenvironment. The retrieved information will provide an inventory and the insight for defining more sophisticated culture conditions that are essential for functional artificial ovarian bioengineering.
Introduction
I
A critical portion of follicular engineering comprises in vitro maturation of immature follicles retrieved from preserved ovarian tissue. To date, a great number of studies have been performed to search for optimized in vitro culture conditions and supportive scaffold biomaterials in the field of artificial ovary engineering. 6 However, the efficiency of in vitro follicular maturation protocols still needs to be improved before they are applied in clinical practice. To enhance follicular survival and growth, the potential efficacy of various antiapoptotic molecules should be evaluated.
An ovarian follicle is regarded as an independent intraovarian organ system that is composed of an oocyte and surrounding follicular cells such as theca and granulosa cells. 7 Folliculogenesis is the process in which recruited primordial follicles grow and one dominant one develops into a matured Graafian follicle with a potential to ovulate its egg to be fertilized. 8 Follicular growth is achieved by cell proliferation and complex combinational regulation of gonadotropin and growth factors. Gonadotropin regulation is controlled by the central nervous system, anterior pituitary. Specialized hypothalamic neurons secrete pulses of gonadotropin-releasing hormone into the portal blood vessels, which causes a pulsatile release of follicle-stimulating hormone (FSH) and luteinizing hormone (LH), which act on ovarian follicle cells to control folliculogenesis. 9 In menstrual cycles, only one follicle is selected as dominant follicle and developed, and others degenerate to follicle cellular apoptosis and finally follicular atresia. 10 At an early stage of follicular development, premature oocytes are at the germinal vesicle stage. To develop mature oocytes, supportive roles of theca and granulosa cells are indispensable.
Numerous factors play a role in in vivo follicular development. During follicular development, endocrine factors such as FSH and LH rescue follicles from apoptotic demise. Paracrine factors are also important in this process and they include epidermal growth factor (EGF), insulin-like growth factor I (IGF-I), basic fibroblast growth factor (bFGF), interleukin 1b (IL-1b), and activin. 11 In addition, noncoding RNAs such as microRNA play important roles during follicular development, and their mimic and inhibitors can potentially be used according to their antiapoptotic or proapoptotic actions (Fig. 1).
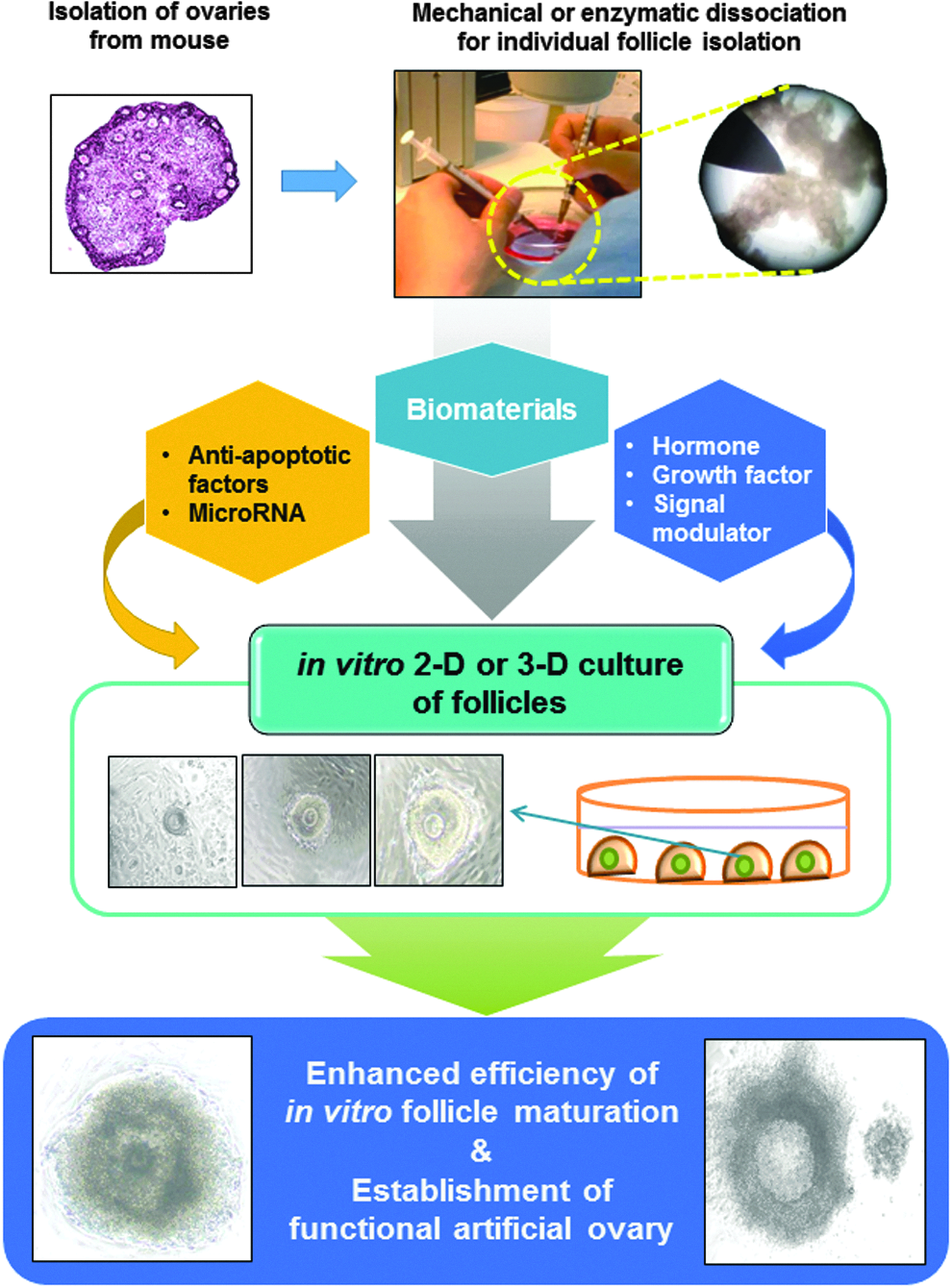
Applicability of potential antiapoptotic factors and noncoding RNAs for efficient 2D or 3D in vitro follicular maturation and artificial ovarian bioengineering. The use of hormones, growth factors, signal cascade regulators, and noncoding RNAs can act against the follicular cell apoptosis during conventional in vitro single follicular maturation procedures.
In vitro support of follicular growth and survival has been a great challenge. Isolated follicles degenerate rapidly when cultured due to a loss of critical connections between the oocyte and granulosa cells. 12 Various protocols of ovarian follicular culture and tissue engineering have been developed for basic and clinical research on fertility preservation. An in vitro culture of isolated follicles or granulosa cells in the absence of serum resulted in spontaneous apoptosis, and the treatment of EGF and bFGF suppressed the spontaneous onset of apoptosis. 13
Since then, many antiapoptotic factors that have a critical role in in vitro single or multiple follicular maturation have been suggested (Tables 3 and 4). Their molecular mechanisms and related intracellular pathways were reported in some species. In addition, the importance of these factors was strongly suggested, which showed direct/indirect upregulation or downregulation of the genes or even epigenetic factors that possess a role in follicular survival. Identification of these antiapoptotic factors not only helps to design better culture media but also to select suitable two- or three-dimensional (2D, 3D) culture conditions that can increase reproductive efficiency.
This review summarizes current knowledge in regard to the roles of antiapoptotic pathways involved in the in vivo folliculogenesis. Currently reported and potential candidate factors that can meet the future needs of ovarian bioengineering will also be introduced. The retrieved information will provide an inventory of potential utilizable factors and the insight for defining the sophisticated culture conditions that are prerequisite of more efficacious ovarian follicular and tissue engineering.
The conceptual applicability of potential antiapoptotic proteins and noncoding RNAs for efficient in vitro follicular maturation and artificial ovarian bioengineering is depicted in Figure 1. The use of hormones, growth factors, signal cascade regulators, and noncoding RNAs can act against the follicular cell apoptosis during conventional in vitro single follicular maturation procedures. The potential use of paracrine and intracrine antiapoptotic factors for in vitro multifollicular maturation is presented in Figure 2. These factors can be used to overcome single follicular culture (Fig. 3) and enhance the yield through their application to simultaneous multifollicular bioengineering. Since the stage of the seeded follicles is important, candidate antiapoptotic factors acting on specific stage of follicular development should also be considered (Fig. 4). Numerous factors have been reported to have significant effects at primordial, primary, secondary, and tertiary follicle stages, respectively, that is, single or combined specific factor(s) should be considered according to the cultured follicle stage(s).
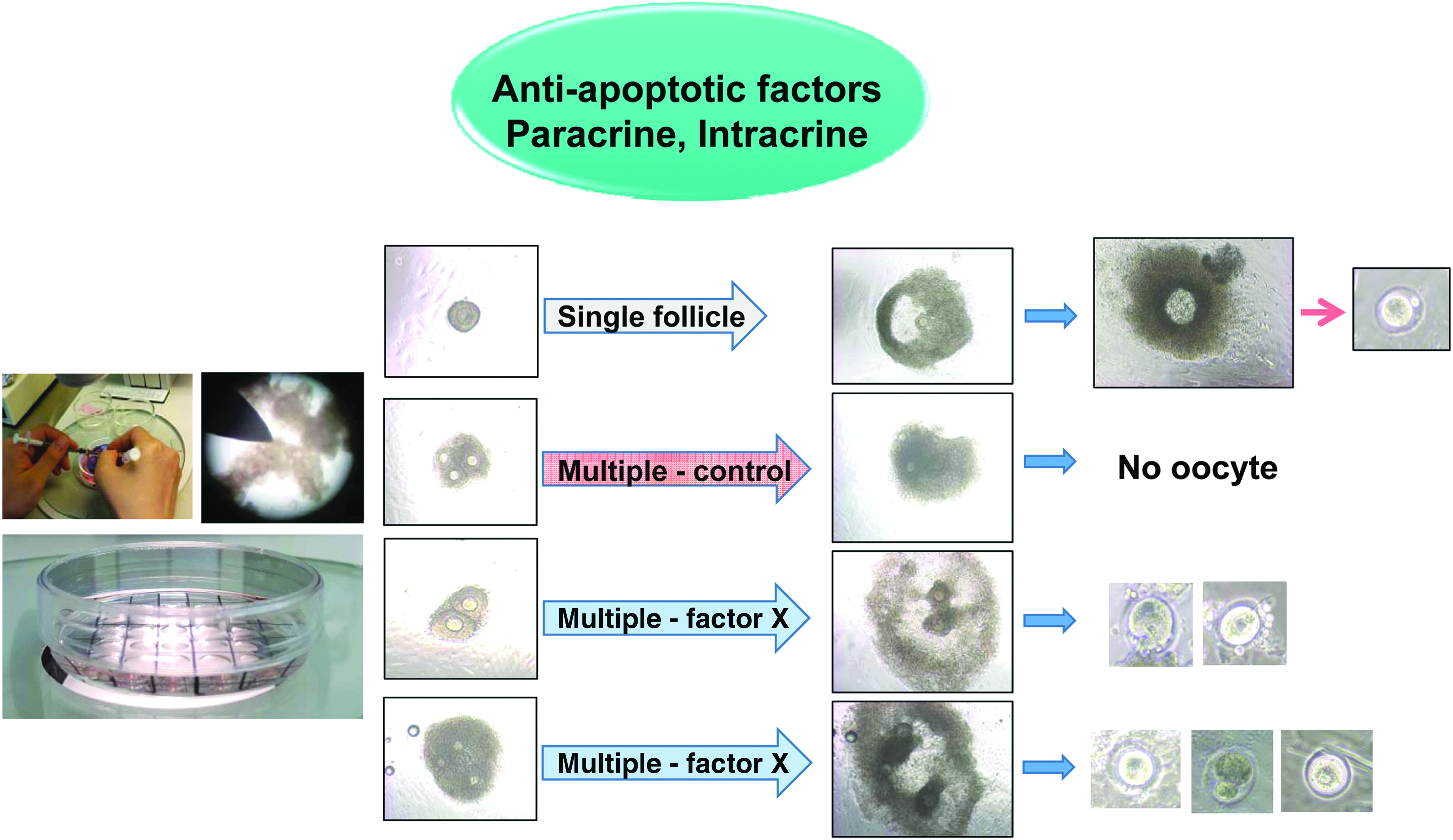
Potential use of paracrine and intracrine antiapoptotic factors for in vitro multifollicular maturation. In conventional in vitro follicle culture, isolated murine follicles were dissociated mechanically under stereo microscope and usually single follicles matured into mature follicles. Multifollicular maturation in vitro is known to be very difficult due to inhibitory signal from adjacent follicle(s). To achieve the multifollicular in vitro maturation, the reviewed factors can be used to overcome the current limitations and enhance the yield through their application to simultaneous multifollicular bioengineering (modified from Kim et al., 2016a). 37a

Development of ovarian follicles. Primordial follicles are derived by an assembly of oocytes from primordial germ cell and somatic cells. Primary follicles showed a layer of granulosa cells that started to proliferate. Secondary follicles have multilayer granulosa cells and proliferating theca cells that respond to FSH, LH, and estrogen compound. Graafian (matured) follicles possessing antrum that are filled with follicular fluid and factors such as inhibin that contribute to ovulation are produced. Mature follicles ovulate mature oocytes according to hormone stimulation. FSH, follicle-stimulating hormone; LH, luteinizing hormone.

Representative schematic diagram depicting the action mechanism of antiapoptotic factors in follicular cells. Gonadotropins are released from pituitary, act on granulosa cells through their membrane receptors, and interact with the growth factors present in follicular fluid and with those secreted from adjacent theca/interstitial cells to promote follicular maturation. Ang-1–7, angiotensin-1–7; Apaf-1, apoptotic peptidase activating factor-1; Bcl-2, B cell/lymphoma-2 family; E2, estradiol; FasL, Fas ligand; FLIP, FLICE-like inhibitory protein; GnRH, gonadotropin-releasing hormone; HGF, hepatocyte growth factor; IGF-I, insulin-like growth factor I; KGF, keratinocyte growth factor; MAPK, mitogen-activated protein kinase; P4, progesterone; PRDX2, peroxiredoxin 2; Srx, sulfiredoxin; TGF-α, transforming growth factor-α; VEGF, vascular endothelial growth factor; XIAP, X-linked inhibitor of apoptosis protein.
Current Use of Antiapoptotic Proteins and Noncoding RNAs in 2D and 3D Follicular Bioengineering
Conventional ovarian follicle culture has been conducted using 2D system. In vitro follicular maturation using 2D system was reported in many studies of ovarian follicles from vertebrate animals, including rodent, pig, cattle, and sheep, 14 and offspring was born using the retrieved oocytes. 15 For this 2D system, multiwell culture 16 or medium droplet culture has been used.17–19 The authors recently reported successful in vitro follicular maturation and retrieval of mature oocytes from murine20–22 and primate ovaries 23 using droplet culture. Recently, using droplet culture, microRNAs (miRs) delivered into follicles altered the oocyte maturation in vitro 2D culture, for example, mmu-let-7c-, mmu-miR-27a-, and mmu-miR-322-inhibitor sequences improved the oocyte maturation rate in mice. 21
To improve the efficiency of development, many strategies have been employed. First, an extracellular matrix (ECM) pretreatment 2D culture was attempted by many researchers,24–27 which enhanced follicular attachment and provision of growth-promoting cytokines. Second, a 3D culture was used to mimic in vivo structural condition and overcome altered in vitro spatial arrangement of 2D system. 28 This mode of culture requires a scaffold made from various degradable biomaterials, including alginate, 28a matrigel, 29 and fibrin.30,31 Despite the improved follicular survival and maturation of 3D culture, some hurdles remain to be resolved, for example, highly variable follicular maturation in some scaffold groups, nonsignificant improvement of postfertilization development rates, 32 and even lower blastocyst developmental rates. 33 Currently reported antiapoptotic factors in 3D follicular bioengineering are summarized with detailed information on co-used scaffold and factors in Table 1.
bFGF, basic fibroblast growth factor; FSH, follicle-stimulating hormone; IGF-I, insulin-like growth factor I; MII, meiosis II; VEGF, vascular endothelial growth factor.
Despite all these strategies of previous reports, the yield still needs to be improved. Recent 2D in vitro murine follicle works using FSH with or without LH showed that the survival rate was 48–93% and ovulation rate was 21–82%, and mature oocytes were retrieved from 41% to 72% of cultured follicles.22,34–37 Recently, Kim et al. 37a successfully induced multifollicular ovulation from murine ovaries using angiotensin II agonist and significantly enhanced the oocyte maturation yields. Other 3D in vitro murine or human studies using FSH with alginate or fibrin showed survival rates of 32–100% and oocyte maturation rates of around 20%.34,38,39 Noteworthy is that the yield and culture protocols markedly varied among different cited studies, which necessitates the standardization of culture and analysis methods (Table 2).
AH-R, aryl hydrocarbon receptor; AMH, anti-Müllerian hormone; Ang-1–7, angiotensin-1–7; Apaf-1, apoptotic protease-activating factor-1; Bcl-2, B cell/lymphoma-2 family; BMP, bone morphogenetic protein; eCG, equine gonadotropic hormone; EGF, epidermal growth factor; GDF-9, growth differentiation factor 9; GH, growth hormone; GHRH, growth hormone-releasing hormone; hCG, human chorionic gonadotropin; ICE, interleukin-1 β-converting enzyme; LH, luteinizing hormone; NF-κB, nuclear factor kappa B; Nobox, newborn ovary homeobox; rpS6, ribosomal protein S6; S6K1, p70 S6 kinase 1; SCF, stem cell factor; SMAD, similar to mothers against decapentaplegics; TGF, transforming growth factor; Tβ-RII, TGF-β receptor type II; VIP, vasoactive intestinal peptide; XIAP, X-linked inhibitor of apoptosis protein.
Most of aforementioned studies were performed using mouse model; however, the reproductive physiology of mice significantly differs from that of human. 8 To overcome this weakness, nonhuman primate ovaries, mostly Rhesus monkey ovaries, have been used for in vitro follicular maturation.40–42 Monkey follicles are larger and require a much longer culture duration, and many investigators attempted to find optimal conditions for in vitro maturation using 3D culture system seeking a shorter culture duration and enhanced maturation efficiency.43–45 Table 3 summarizes the primate follicle studies using a 3D culture system.
Taken together, the beneficial roles of antiapoptotic factors, either protein or noncoding RNAs, have been strongly suggested, however, the effects of a scaffold during in vitro follicular bioengineering should be considered simultaneously. Using this inventory, the mimics of antiapoptotic miRs and/or inhibitors of apoptotic miRs can be added to in vitro artificial ovary bioengineering microenvironment.
Candidate Antiapoptotic Molecules and their Action Mechanisms
Major antiapoptotic mechanisms are divided into the binding of endocrine and paracrine molecules to the receptors of cytoplasmic membrane and the signaling within the follicular cells. 46 A complex interaction between theca, interstitial, and granulosa cells mediates the hormonal regulation of apoptosis in granulosa cells during follicular atresia.47,48 Many studies reported that the endocrine factors such as estrogen, FSH and LH, and transforming growth factor (TGF) rescue follicles from apoptotic demise. These paracrine factors can induce activation or upregulate the expression of other molecules in the oocyte, theca, and/or granulosa cells indirectly, which prevent or suppress the apoptotic mechanisms.
Gonadotropins
Gonadotropins such as FSH and LH are secreted from pituitary gland and act as effective antiapoptotic agents through their receptors, respectively, on granulosa and theca cells. 49 Primordial follicles are at a gonadotropin-independent phase and thereafter follicles become responsive to gonadotropins. 50 Beyond this stage, all the processes of follicle growth, recruitment, selection, and ovulation are gonadotropin dependent. 49 Gonadotropins induce the expression of Bcl-2, FLIP, GATA, and XIAP (antiapoptotic molecules) and downregulate the expression of Bax, Fas/FasL, Apaf-1, and p53 (proapoptotic molecules) in in vitro and in vivo studies of mouse and rat.51–53 Intrafollicular IGF-I and theca cell-derived paracrine factors such as TGF-α, keratinocyte growth factor (KGF), and hepatocyte growth factor (HGF) mediate the gonadotropin-induced suppression of apoptosis in granulosa cells of antral follicles in vitro in rat.13,47,48,54 The prosurvival effects of gonadotropins on granulosa cells through direct or indirect upregulation of antiapoptotic molecules or downregulation of proapoptotic molecules are depicted in Figure 5.
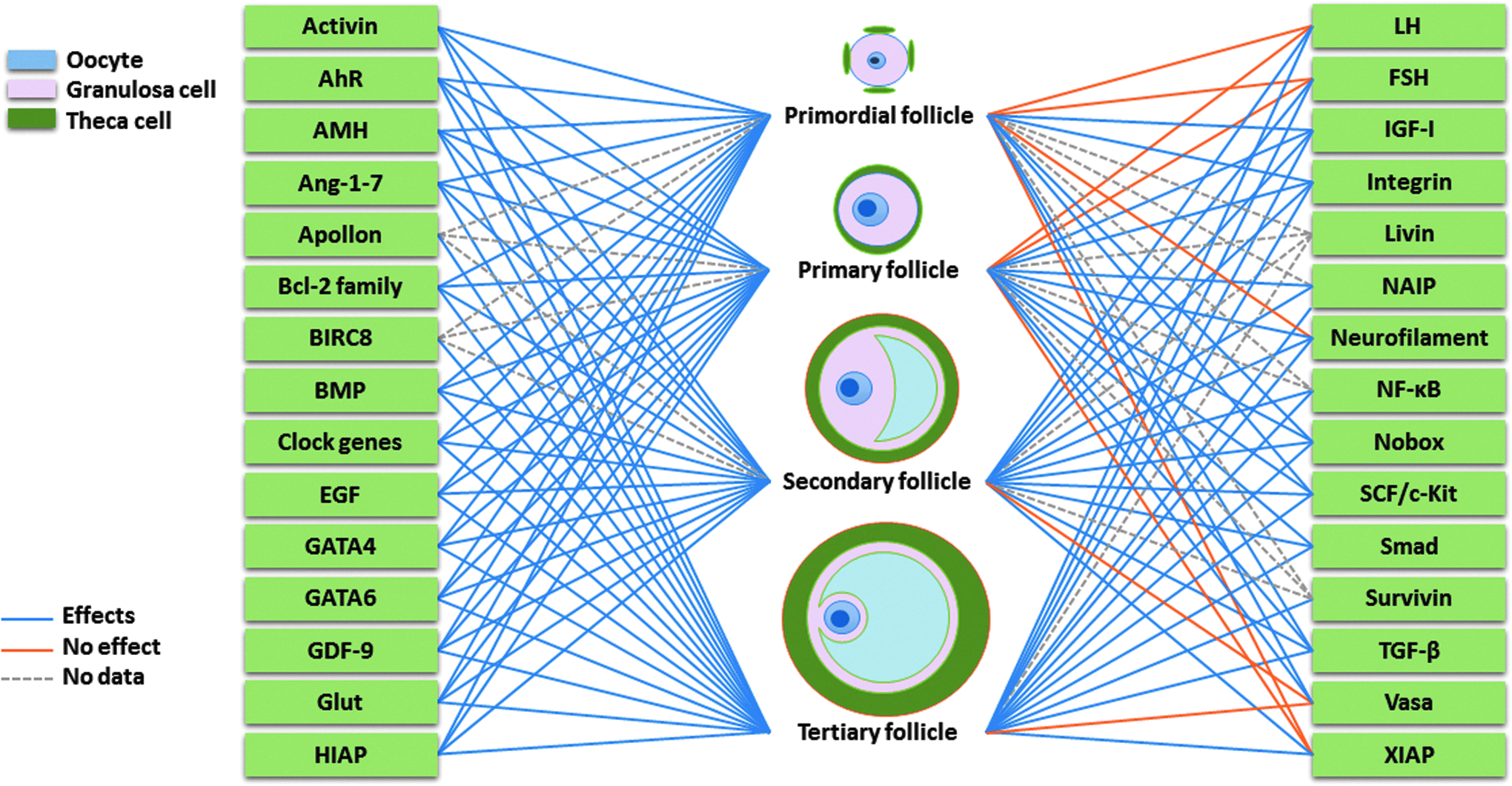
Candidate antiapoptotic factors acting on specific stage of follicular development. Listed factors have been reported to have significant effects at primordial, primary, secondary, and tertiary follicle stages, respectively, that is, single or combined specific factor(s) should be considered according to the cultured follicular stage(s). AhR, aryl hydrocarbon receptor; AMH, anti-Müllerian hormone; BIRC8, baculoviral IAP repeat containing 8; BMP, bone morphogenetic protein; EGF, epidermal growth factor; GDF-9, growth differentiation factor 9; Glut, glucose transporter proteins; HIAP, human inhibitor of apoptosis protein; NAIP, neuronal apoptosis inhibitory protein; NF-κB, nuclear factor kappa B; Nobox, newborn ovary homeobox; SCF, stem cell factor; TGF-β, transforming growth factor β.
TGF family
TGF family members are signaling molecules that regulate the interaction between follicular cells and ECM. They include TGF-β, activin, inhibin, bone morphogenetic proteins (BMPs), growth differentiation factor-9 (GDF-9), anti-Müllerian hormone (AMH), IGF-I, TGF-β2, bFGF, epidermal growth factor receptor (EGF-R), and many other structurally related proteins.55,56
Transforming growth factor-β
TGF-β1, 2, and 3 are mammalian isoforms of TGF-β, which represent secreted ligands, and downstream signaling events are initiated by binding of TGF-β ligands and activation of their transmembrane receptors. TGF-β interacts with the TGF-β receptor-2 (TGF-β-R2), and the ligand–receptor complex subsequently binds to a TGF-β-R1, which makes a heterotetrameric receptor complex. The TGF-β2 receptor kinase activates TGF-β-R1, which activates the phosphorylation of TGF-β-R1 kinase and downstream signaling, including Smad phosphorylation.57–59
In the ovary, the ligands, receptors, and mediators of TGF-β have important roles during in vivo follicle maturation. 60 Likewise, TGF-β signaling has a physiological role in maintaining dormant primordial follicles, which functions through activation of p70 S6 kinase 1 (S6K1)/ribosomal protein S6 (rpS6) signaling in mouse ovaries. 61 Although stimulatory effect of TGF-β on survival of primordial follicles are shown, there is some evidence of its inhibitory effect on survival of primary follicle as well as on progression to the late antral stages. The downregulation of TGF-β-R1 expression or inhibition of the TGF-β1/TGF-β-R1 ligand activates the oocyte growth of early primordial follicles and the proliferation of granulosa cells in mouse in in vivo condition. 61
Activin
Activin and inhibin are closely related protein complexes with opposite biological effects. Inhibin downregulates FSH synthesis and inhibits FSH secretion. Conversely, activin enhances FSH biosynthesis and secretion, and participates in the regulation of ovarian cell proliferation, differentiation, and apoptosis. In rat ovary culture, activin induced primordial follicle development to preantral follicles and decreased the proportion of atretic follicles in the primary and preantral follicles. 62 In sheep follicle, activin B and activin A increased the follicle size and suppressed thecal androgen production in vitro. 63 Furthermore, the inhibitory effect of activin on androgen production was blocked by inhibin in antral follicle of sheep. 63 In in vitro culture, activin also suppressed production of progesterone and androgen in theca cells, while inhibin enhanced production of androgen in human 64 and cow. 65
Growth differentiation factor-9
GDFs, are the subfamily of proteins, belong to TGF-β superfamily that affects embryonic development. 66 GDF-1 functions left–right patterning and mesoderm induction during embryonic development. 67 GDF-2 induces and maintains the response to a neurotransmitter acetylcholine, and regulates iron metabolism.68,69 GDF-3 expression occurs in ossifying bone during embryonic development and in the thymus, spleen, bone marrow, brain, and adipose tissue of adults.70,71 GDF-5 has roles in the development of joints and the skeleton, and increasing the survival of neurons that respond to dopamine.72–74 GDF-6 interacts with BMPs to regulate ectoderm patterning, and controls eye development.75–77 GDF-8 controls the growth of muscle tissue. 78 GDF-10 has roles in head formation.79,80 GDF-11 controls anterior–posterior patterning by regulating Hox genes 81 and regulates the number of olfactory receptor neurons 82 and retinal ganglionic cells developing in the retina. 83
GDF-9 expression is limited to the ovaries, and endogenous GDF-9 plays an important role in the formation of primordial follicles and somatic cell differentiation.84,85 A high level of GDF-9 expression is observed solely in the oocytes of mice in vivo. 86 During oocyte maturation in vivo, the expression of GDF-9 and BMP-15 (or GDF-9B) changes dynamically after ovarian stimulation, with a decrease at the metaphase I (MI) stage and an increase to a peak level at the metaphase II (MII) stage in women. 87 Gonadotropins increase the expression of GDF-9 mRNA and protein, and introduction of GDF-9 siRNA suppresses the effect of gonadotropin in mouse and hamster follicles in both in vitro and in vivo conditions,88,89 whereas they have no effect on the expression of BMP-15 in mouse ovary. 88 GDF-9 deficiency blocks folliculogenesis and results in the loss of 17α-hydroxylase, c-Kit, and LH receptors on theca cell layers. Taken together, the role of GDF-9 in follicle development is indispensable.
BMP family
BMPs are multifunctional growth factors that belong to TGF-β superfamily. Smad-1, -5, and -8 are the downstream molecules of BMP receptors and play major roles and BMP-2A, -2B, -3, -3b, -4, -6, -7, and -15 are the ligand members of BMP family. 90
The BMP family gene expression affects follicular recruitment, selection, atresia, ovulation, luteinization, and luteolysis. In this regard, in a cell-specific manner, the expression of BMP mRNA is observed in the granulosa cells, oocytes, theca cells, theca externa, interstitial cells, and corpus luteum of rat in vivo.91,92 Genes such as Nog, Htra1, Fst, Bmper, and Vwc2S interact with BMPs, are positively associated with the emergence of developing follicles in mouse in vivo, 93 and treatment with anti-BMP-4 decreases the number of primary follicles and increases the numbers of primordial follicles in mouse ovary. 94 BMP-6 increases growth-regulated oncogene-α expression in human granulosa cells in vitro. 95 BMP-7 is the cytokine expressed on theca interna cells, oocytes, and granulosa cells of growing follicles, and is not expressed in primordial or primary follicles. It is expressed in both oocytes of secondary follicles and all the somatic cells of antral follicles, interstitial cells, corpora lutea, and ovarian surface epithelial cells of rabbit in vivo. 96 BMP-15 plays an important role in follicular development. BMP-15 protein is highly expressed on the oocytes of primary follicles, but not on those of primordial follicles in rabbit in vivo. 97 The expression of Bmp15 and Gdf9 is maintained during follicular maturation and then declines after ovulation in chicken in vivo. 98 Considering the known effects of BMPs on the folliculogenesis, the application of this factor in artificial ovarian engineering or 3D culture of follicles should be considered.
Anti-Müllerian hormone
AMH, a member of TGF-β superfamily, is secreted by the granulosa cells. Inhibition of gonadotropin secretion during cycle days 0–10 decreases the AMH expression on preantral and antral follicles. 99 In addition, in in vitro culture of mice ovarian follicle, AMH regulated the effect of FSH on follicular growth. 100 On the other hand, changes in the hormonal milieu during controlled ovarian hyperstimulation during in vitro fertilization cycles and after the LH surge may affect AMH secretion in women in vivo.101,102 In addition, inhibitory effect of recombinant AMH on early follicular development in vitro by suppressing the growth of human primordial follicle has been shown. 103 Taken together, gonadotropic hormones seem to be involved in the regulation of AMH expression. 99
Insulin growth factor-I
IGFs and IGF binding proteins (IGFBPs) play a major role in the autocrine and paracrine regulation of apoptosis. IGF-I has survival and stimulatory effects on primordial and primary follicles in goat in vitro. 104 The granulosa cells of antral follicles express IGF-I and its receptors (IGF-I-R), and the completion of follicular development in human and rats in vivo requires IGF-I.105,106 The expression of igf-I mRNA is increased in the primary to antral stage follicles and leads to a decrease in the caspase 3 activity in hamster in vitro.56,107 Theca and granulosa cells express IGF-I mRNA in vivo at all follicular developmental stages in rat. 108 Oocytes also express both IGF-I and its receptor in monkey, and androgen increased their expression in primordial follicles in vivo. 109 The expression of IGF-I in the follicle at every developmental stage is generally low throughout estrous cycles, and IGFBP-1 mRNA is not detected. In sheep, the expressions of both IGF-I-R and IGFBP-2, -3, -4, -5, and -6, decrease as follicles increase in diameter in vivo. 110 Taken together, IGF-I involved in the regulation of folliculogenesis in many species.
Epidermal growth factor
EGF plays an antiapoptotic role in the intrafollicular paracrine communication. 111 EGF is expressed on the oocytes, whereas EGF-R is expressed on the follicular cells and plays a role in the oocyte-to-follicular cell signaling pathway. EGF expresses in theca cells of preantral to preovulatory follicles, granulosa cells of preantral follicles, and oocytes of growing human follicles in vivo. 112
Bcl-2 family
A relationship is observed between follicular development and the expression of the apoptosis-regulating protein Bcl-2 and Bax in rat and mouse ovaries in vivo.113,114 The Bcl-2 gene has been found in vivo and in vitro in the ovary of many species, including human, rat, and drosophila.115–117 Knockout of Bcl-2 results in fewer oocytes and primordial follicles in the postnatal mouse ovaries in vivo. 118 Furthermore, the overexpression of Bcl-2 increased the primordial follicles in mice postnatal life in vivo. 119 Furthermore, in rat theca cells, gene products of bcl-2 and interleukin-1 beta-converting enzyme are involved in the apoptotic signal transduction pathway induced by TGF-α and TGF-β in vitro. 120 Also, it is shown in mouse that the expression of Bcl-2 has antiapoptotic effect in oocytes in vitro. 121 Bcl-2 is expressed mainly in developing follicles and plays an antiapoptotic role in granulosa cells of fetal and adult ovaries in mouse, rat, and quail in vivo.122–124
Stem cell factor and c-Kit
Stem cell factor (SCF) is a growth factor that activates c-Kit receptor and affects profoundly the proliferation, migration, differentiation, and survival of the ovary. Kit and Mgf gene express c-Kit receptor and ligand, and their interactions have a prosurvival effect on oocytes and theca cell differentiation of the primordial, primary, and antral follicles of pig in vivo. 125 C-Kit and SCF are expressed on oocytes of preantral follicles, weakly on granulosa and theca cells, and on oogonia of the primordial and primary follicles and on granulosa and theca cells of antral follicles in mice in vivo. 126 The expression of c-kit/SCF in oocytes is specific at all the follicular developmental steps, is gradually decreased as the follicles develop, negatively regulates Fas-mediated apoptosis in vivo, and is decreased after the follicles start to grow in monkey. 127 Moreover, SCF can downregulate the expression of BAX and therefore reduce the apoptosis of oocytes in mouse in both in vivo and in vitro condition. 128 In the antral follicles, the c-Kit receptor modulates the ability of the oocytes to enhance thecal androgen output and undergo cytoplasmic maturation. 129
Inhibitor of apoptosis family
The members of inhibitor of apoptosis protein (IAP) family, including baculovirus IAP repeat-containing protein 1 (BIRC1 or neuronal apoptosis inhibitory protein, NAIP), BIRC2 (c-IAP-1 or human inhibitor of apoptosis protein 2, HIAP-2), BIRC3 (HIAP-1 or c-IAP-2), BIRC4 (X-link inhibitor of apoptosis protein, XIAP), BIRC5 (Survivin), BIRC6 (Apollon or BRUCE), BIRC7 (livin, melanoma inhibitor of apoptosis, ML-IAP, or kidney inhibitor of apoptosis protein, K-IAP), and BIRC8 (inhibitor of apoptosis protein-like protein-2, ILP2 or testis-specific inhibitor of apoptosis protein, TsIAP), are the eight known antiapoptotic factors that inhibit the action of caspases. 130
X-linked inhibitor of apoptosis protein
XIAP is a member of IAP family that inhibits caspase in the ovary in vivo. 131 FSH upregulates XIAP in the rat ovary, which leads to granulosa cell apoptosis suppression and FSH-induced follicular growth in vitro. 53 Moreover, XIAP is a critical intracellular modulator of the death-inducing TNF-α signaling pathway in granulosa cells and is upregulated by a Nuclear factor kappa B (NF-κB)-mediated mechanism and the phosphatidylinositol 3-kinase (PI3-K) survival pathway in rat in vivo and in vitro.132–134 The presence of XIAP mRNA and protein is observed in isolated granulosa, theca, luteal, and stromal cells throughout the estrous cycle of sheep in vivo.135,136 An increasing number of active caspase-3-positive cells are associated with a reduction in XIAP mRNA expression in antral follicles of sheep in vivo. 135 In the sheep ovary in vivo, XIAP protein is present from the primary follicular stage onward, whereas its mRNA is not detected in primordial, primary, and secondary follicles. 136
Survivin
Survivin is a prosurvival molecule confined to the mitotic spindle and interacts with caspases. Survivin is encoded by the BIRC5 gene in humans in vitro. 137 Survivin is expressed at a high level on mitotically active avian granulosa cells during follicular development and regulates the cell cycle and inhibits apoptosis. 138 In African clawed frog, Xenopus laevis, survivin mRNA accumulates during oogenesis and is present in the early-stage oocytes in vitro. 139 Furthermore, in cultured human granulosa cells, human chorionic gonadotropin (hCG) increased survivin mRNA expression. 140 The pattern of survivin protein expression at various follicular development stages is not clearly defined yet.
Nuclear factor kappa B
NF-κB is a group of transcription factors which using DNA-binding proteins regulate NF-κB-related genes. NF-κB has an antiapoptotic effect by downregulation of TNFα-induced pathways and upregulation of Bcl-2 and IAP families.141–143 In rat granulosa cell culture, FSH induced activation of NF-κB and caused increase of XIAP gene transcription through the PI3K/Akt pathway. 144 In bovine oocytes, the level of NF-κB/p65 DNA binding increased by maturation of oocytes in vitro. 145 Although the presence of NF-κB in follicular cells of antral follicles in human and cow are shown, probable alterations of this factor during the follicular development is not clarified.
Integrins
In the ovary, ECM components of the follicular basement membrane and those in the follicular fluid mediate the regulation of follicular development, and, in this process, the role of integrins is important. 146 Integrins express on the primordial follicle surface of mouse ovary in vitro, link the follicular cells to the somatic and neighboring cells and ECM, as well as participate in signal transduction. 147 Integrin expression in ovarian follicles of monkey is associated with granulosa cell apoptosis of tertiary follicles in vivo. 148 The decrease or lack of integrin α6/β1 expression leads to the apoptosis of granulosa cells of tertiary follicles and atretic follicles in primate ovary in both in vivo and in vitro conditions.149,150
Clock genes
Clock genes, components of the circadian clock, interact with each other to generate gene expression oscillations in ovarian tissue in vivo and knocking out of these genes induces subfertility in mice. 151 In the rat ovary, the expression of clock genes becomes rhythmic following hCG stimulation in the granulosa cells of preovulatory follicles in vivo. Administration of gonadotropins or glucocorticoids can affect the expression on circadian clock genes in granulosa cells and corpus luteum cells in rat in vitro. 152 In luteal cells, inhibition of progesterone synthesis in apoptotic condition deregulated Per2 circadian oscillation and reduced transcription of Bmal1 and Clock. 152 Further studies are necessary to show the exact role of these genes and their interactions with gonadotropins in antiapoptotic processes of follicular cells.
Newborn ovary homeobox
The newborn ovary homeobox (Nobox) is encoded by eight exons and is an oocyte-specific homeobox gene. 56 High levels of Nobox mRNA expression occur in oocytes of primordial follicles through growing follicles, but not in granulosa and theca cells and corpora lutea in mice in vivo. 153 It is shown in mice oocyte that the expression of Nobox is highest in oocytes of antral follicles in normal estrus cycle, while gonadotropin administration increased its expression in oocytes of preantral follicles in vitro. 154 On the other hand, downregulation of GDF-9 in Nobox knockout mice increases postnatal oocyte loss and ovarian follicular fibrosis, and suppresses the transition from primordial to growing follicles in vivo.153,155
Renin–angiotensin system
The renin–angiotensin system that is expressed in the ovary and regulated by gonadotropins has a role in the follicular development. In rabbits, Ang II affects steroidogenesis and ovulation in vivo. 156 Both granulosa and theca cells produce angiotensin-1–7 (Ang-1–7) after stimulation by LH in rat, and also, treatment of oocytes with Ang-(1–7) in follicle culture resulted in maturation of oocytes in vivo. 157 In pig ovaries in vivo, Ang II type 1 and 2 receptors (AGT-R1 and AGT-R2) are detected in the granulosa cells of primordial, primary, and secondary follicles. 158 In periovulatory follicles of cow, AGT-R2 are mainly detected in the theca cells in vivo. 159 Recently, Kim et al. reported that intra-ovarian angiotensin system may be affected by estradiol treatment (2016) 159a and freezing–thawing (2016). 159b
Noncoding RNAs
Recent knowledge from genome-wide analysis shows that noncoding RNAs (ncRNAs) transcribe two thirds of the genome. Noncoding RNAs include transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), small nucleolar RNAs (snoRNAs), microRNAs (miRs), small interfering RNA (siRNA), small nuclear ribonucleic acid (snRNA), extracellular RNA (exRNA), piwi RNAs (piRNAs), and long ncRNAs (lncRNAs). According to the structure, size, and regulatory properties, they are classified into two categories. Long-size lncRNAs and short- and medium-size ncRNAs such as snoRNAs, piRNAs, and miRNAs. 160 Although not fully understood, they are reported to be involved in various biological and pathological phenomena.161,162
Long noncoding RNAs
Long non-coding RNAs (lncRNAs) play its role by regulating epigenetic effectors, microRNA-like molecules, and alternative splicing. 163 lncRNAs are negative apoptosis regulators of different types of cancer cells. 164 Estrogen modulates lncRNAs TC0100223, TC0101686, and TC0101441 expression in epithelial ovarian cancer cells in vitro. 165 Male hypermethylated (MHM) region is a Z chromosome-linked locus that is transcribed into an lncRNA in female chicken embryo in vivo and has an identified role in ovarian development. 166 Potential candidate lncRNAs are summarized in Table 4. In future, the mimic of apoptosis inhibitors and cell proliferating lncRNAs and/or the antagonist of apoptosis stimulators should be developed to enhance the efficiency of in vitro ovarian tissue bioengineering.
MicroRNAs
MicroRNAs (miRs), short noncoding RNA molecules, play a negative regulatory role by binding to the 3'-untranslated region (3'-UTR) of the target mRNAs. 167 They are involved in the cellular proliferation, differentiation, metabolism, oncogenesis, and apoptosis. Cell proliferation inhibitors include let-7, miR-125b, miR-133, miR-143, miR-17-92, miR-221, miR-222, miR-224, miR-98, and miR-lat; and they affect endocrine and intracellular regulators of the cell cycle.168–171 The cell proliferation stimulator miRNAs, including miR-17, miR-372, and miR-373, affect the cell cycle inhibitors. 168 Some apoptosis promoter miRNAs such as miR-34a, miR-34b, miR-34c, and miR-214 activate the production of caspases, p21, p53, and bax.168,172–174
MiR-21 controls the apoptosis of granulosa cells: its absence increases the apoptosis and its increase protects against the apoptosis of granulosa cells in mice in vitro. 175 MiR-21 reduces apoptosis by affecting the activation of the antiapoptotic PTEN/protein kinase, interleukin/STAT3, PDCD4, and B/PI3 kinase.171,176–178 E2F transcription factor 1 (E2F1), regulated by miR-320, stimulates the apoptosis of granulosa cells in mice in vivo and in vitro through p53-dependent pathways. 179 Proliferation of granulosa cells is inhibited by miR-145 through the suppression of acvrib mRNA and protein expression in mice in vitro. 180 The upregulation of TGF-β-R2 and stimulation of Smad 3, antisense miR-145, promote follicular growth in mice in vivo. 181 MiR-181a inhibits granulosa cell proliferation by suppressing ACVR2A-induced Smad 2 phosphorylation in mice in vitro. 182
MicroRNAs affect oocyte maturation during ovarian follicle development in mice in vitro.20,21 MiR-23a decreases XIAP expression and induces apoptosis in human ovarian granulosa cells in vitro. 183 Transfection with an mmu-miR-27a-mimic sequence decreases the oocyte maturation rate, and transfection with mmu-let-7c-, mmu-miR-27a-, and mmu-miR-322-inhibitor sequences increases the oocyte maturation rate in mice in vitro.20,21 The expression of IGFBP-2 is higher in the adjacent granulosa cells of MII oocytes than in those of MI oocytes and is higher in miR inhibitor transfection groups than in miR mimic transfection groups and controls.20,21
The miRs involved in either upregulation or downregulation of apoptosis in ovarian granulosa cells are shown in Table 5. Mimics of antiapoptotic miRs and/or inhibitors of apoptotic miRs can be added to in vitro maturation or artificial ovary bioengineering microenvironment, for example, enhancing the oocyte maturation rate by transfecting granulosa cells with mmu-let-7c-, mmu-miR-27a-, and mmu-miR-322-inhibitor sequences. 21
cf. Mimics of antiapoptotic miRs and/or inhibitors of apoptotic miRs can be added to in vitro maturation or artificial ovary bioengineering microenvironment, for example, transfection with mmu-let-7c-, mmu-miR-27a-, and mmu-miR-322-inhibitor sequences increased the oocyte maturation rate (Kim et al., 2013).
Conclusions
Ovarian follicular maturation using in vitro culture is a promising bioengineering field for female fertility preservation for reproductive age cancer survivors and premature ovarian failure patients. Successful clinical application requires the protocols that can efficiently overcome follicular cell apoptosis since apoptosis is a critical phenomenon in in vitro follicular maturation and in in vivo folliculogenesis. The use of antiapoptotic agents in the in vitro follicular cell processing is at an early stage of establishment. The insight on their roles will contribute to the manufacturing of potential therapeutic cell/tissue products for the treatment of female reproductive organ failure.
Although various materials have been suggested to be utilized for 3D culture as ECM or scaffold, the reported maturation yields still need to be improved. The addition of aforementioned prosurvival and antiapoptotic factors can a feasible option in many 3D culture settings.
Conclusively, the authors searched literature and analyzed the current knowledge of these proteins and noncoding RNAs, and their antiapoptotic roles in the dynamics of follicular development in vivo and in vitro. Through this review, currently available antiapoptotic molecule directory was retrieved. It can provide a wide range of selection when establishing an optimal in vitro ovarian follicular culture condition and ovarian tissue engineering.
Footnotes
Acknowledgment
This study was supported by grants of the Korean Health Technology R&D Project, Ministry of Health & Welfare, Republic of Korea (HI14C2289 and HI14C2259).
Authors' Contribution
Y.Y.K participated in designing the study and proofreading. The systematic literature search and preparing the first draft were performed by A.T. S-Y.K. contributed substantially to the design of the literature review and article writing. All authors edited and approved the final draft.
Disclosure Statement
No competing financial interests exist.