
Abstract
Mechanical signals offer a promising way to control cell and tissue development. It has been established that cells constantly probe their mechanical microenvironment and employ force feedback mechanisms to modify themselves and when possible, their environment, to reach a homeostatic state. Thus, a correct mechanical microenvironment (external forces and mechanical properties and shapes of cellular surroundings) is necessary for the proper functioning of cells. In vitro or in the case of nonbiological implants in vivo, where cells are in an artificial environment, addition of the adequate mechanical signals can, therefore, enable the cells to function normally as in vivo. Hence, a wide variety of approaches have been developed to apply mechanical stimuli (such as substrate stretch, flow-induced shear stress, substrate stiffness, topography, and modulation of attachment area) to cells in vitro. These approaches have not just revealed the effects of the mechanical signals on cells but also provided ways for probing cellular molecules and structures that can provide a mechanistic understanding of the effects. However, they remain lower in complexity compared with the in vivo conditions, where the cellular mechanical microenvironment is the result of a combination of multiple mechanical signals. Therefore, combinations of mechanical stimuli have also been applied to cells in vitro. These studies have had varying focus—developing novel platforms to apply complex combinations of mechanical stimuli, observing the co-operation/competition between stimuli, combining benefits of multiple stimuli toward an application, or uncovering the underlying mechanisms of their action. In general, they provided new insights that could not have been predicted from previous knowledge. We present here a review of several such studies and the insights gained from them, thereby making a case for such studies to be continued and further developed.
Introduction
R
Mechanotransduction, or the mechanisms by which cells convert mechanical stimuli to biochemical change, in itself, is a field of intense study. Cells sense and respond to external forces and resistance to their internal forces. The underlying mechanism involves the forces changing the structure of molecules or molecular assemblies, leading to a change in their function. In principle, all large biomolecules can undergo such changes. However, a few key players have been identified—the cytoskeleton, nuclear lamina, adhesion proteins, surface processes, and stretch-activated channels. 1
The cell cytoskeleton is one of the most important structures. It consists of a network of filaments and tubules that are distributed over the entire cytoplasm. 5 Thus, any force transfer inside the cell tends to involve it. The cytoskeleton has been shown to be a tensegrity (tensional integrity) structure with elements under compression (microtubules) that are held together by elements under tension (microfilaments, intermediate filaments). 6 This structure enables it not just to bear loads and give the cells their shape but also to respond quickly to external forces. In addition, the cytoskeleton generates contractile forces (actomyosin fibers) that enable the cell, among other things, to move and to assess its environment by tugging at it. 5 Many cells also develop protrusions or surface processes, such as the primary cilia, that are structurally supported by cytoskeleton molecules but have associated specialized machinery (receptors, channels) that can be activated through the deflection of these processes by external forces. 7 To be sensed by the cells, external forces need to produce displacements and this requires the cells to be constrained in some way.
Two kinds of cell constraints exist for adherent cells, namely attachment to the extracellular matrix (ECM) and attachment to other cells. These constraints use specialized membrane proteins or molecular assemblies (cadherins 8 and gap junctions 9 for cell-cell attachment, integrins and focal adhesions for cell-ECM attachment 10 ) that are connected to the cytoskeleton. Biochemical changes resulting from mechanical stimuli can begin at the cell membrane itself via conformational changes in membrane proteins, or inside the cell as the forces are transmitted inside through the constraint points.
Longer-term biochemical changes involve changes in gene expression. This can happen not only due to downstream reactions resulting from the mechanical stimuli 11 but also by the direct transfer of forces to the nucleus. 12 The nucleus containing the chromatin (DNA and associated molecules) is surrounded by the nuclear lamina that shares connections with the cytoskeleton, providing a route for transfer of forces to the nucleus. 13 All these components are dynamically regulated by cells to maintain the mechanical microenvironment of the cells, and thus the tissue, in a state of mechanical homeostasis, a disruption of which can have pathological implications.14,15 An understanding of the actions of these key players as well as their regulation in response to various mechanical stimuli can provide a mechanistic understanding, and hence predictive ability, about the responses of cells to the stimuli. For example, cell alignment in response to stretch can be understood in terms of cytoskeleton energy minimization, 16 or the cytoskeleton contractility could explain the sensing of substrate stiffness by cells. 17
The main ways in which mechanical stimuli are applied to cells in vitro include stretching the substrate to which cells are attached, fluid flow over cells applying shear stress to the cells, compressive forces such as hydrostatic pressure, varying the substrate stiffness, varying the substrate topography, and controlling the cell attachment area by using patterning of the substrate. These stimuli are primarily inspired by the in vivo environment of cells and are applied to whole cells or cell populations. Other techniques such as magnetic tweezers, optical tweezers, and atomic force microscopy have been employed for subcellular mechanical stimulation along with force quantification to understand mechanotransduction mechanisms. 18 And yet more techniques, such as traction force microscopy 19 and micropillar arrays, 20 have been used to measure the forces exerted by cells on their surroundings.
However, in all the techniques used, simplifications were used to study the effects of a particular stimulus. The application areas, namely tissue engineering and predictive cell cultures for pharmaceutical discovery, require providing an in vivo like environment for cells to elicit in vivo like behavior. In terms of mechanical stimuli, the in vivo situation is often complex, with multiple stimuli being simultaneously present. In the absence of a complete understanding of cellular mechanotransduction mechanisms (i.e., which molecular/structural players are involved and how they are regulated over time in response to the stimuli), it is difficult to predict the behavior of cells in a complex mechanical environment. Applying combinations of mechanical stimuli in vitro offers a way to further investigate the mechanotransduction mechanisms as well as to find optimal in vitro stimuli for the application areas. In addition, the limits of the mechanical stimuli that cells can withstand can be tested 21 as well as pathological conditions can be simulated by applying stimuli that disturb the cell/tissue mechanical homeostasis.22–24
Although macroscale mechanical bioreactors were explored in the past to apply tightly controlled combined mechanical stimulation to tissue constructs prepared from cells and materials, 25 most studies now employ micro or meso scale systems that not only allow for good control on the stimuli but also permit constant monitoring of the cells inside. Most reported studies so far have focused on the approaches and the development of the experimental setups to apply combinations of mechanical stimuli, often with an application in mind. As these experimental techniques become established, probing the mechanotransduction pathways is now also starting to be explored.
The aim of this review is to highlight the studies that have applied combinations of mechanical stimuli to cells, focusing on more recent examples. Micro and meso scale platforms to apply mechanical stimuli to cells have been reviewed before 26 but we will focus here on those that look at a combination of multiple stimuli.
Application of Combinations of Mechanical Stimuli
This section highlights examples of combined stimulation studies for various combinations of the five mechanical stimuli schematically shown in Figure 1. At the beginning of the sub-sections, a brief introduction to the known effects of each stimulus alone is also provided for context and comparison.
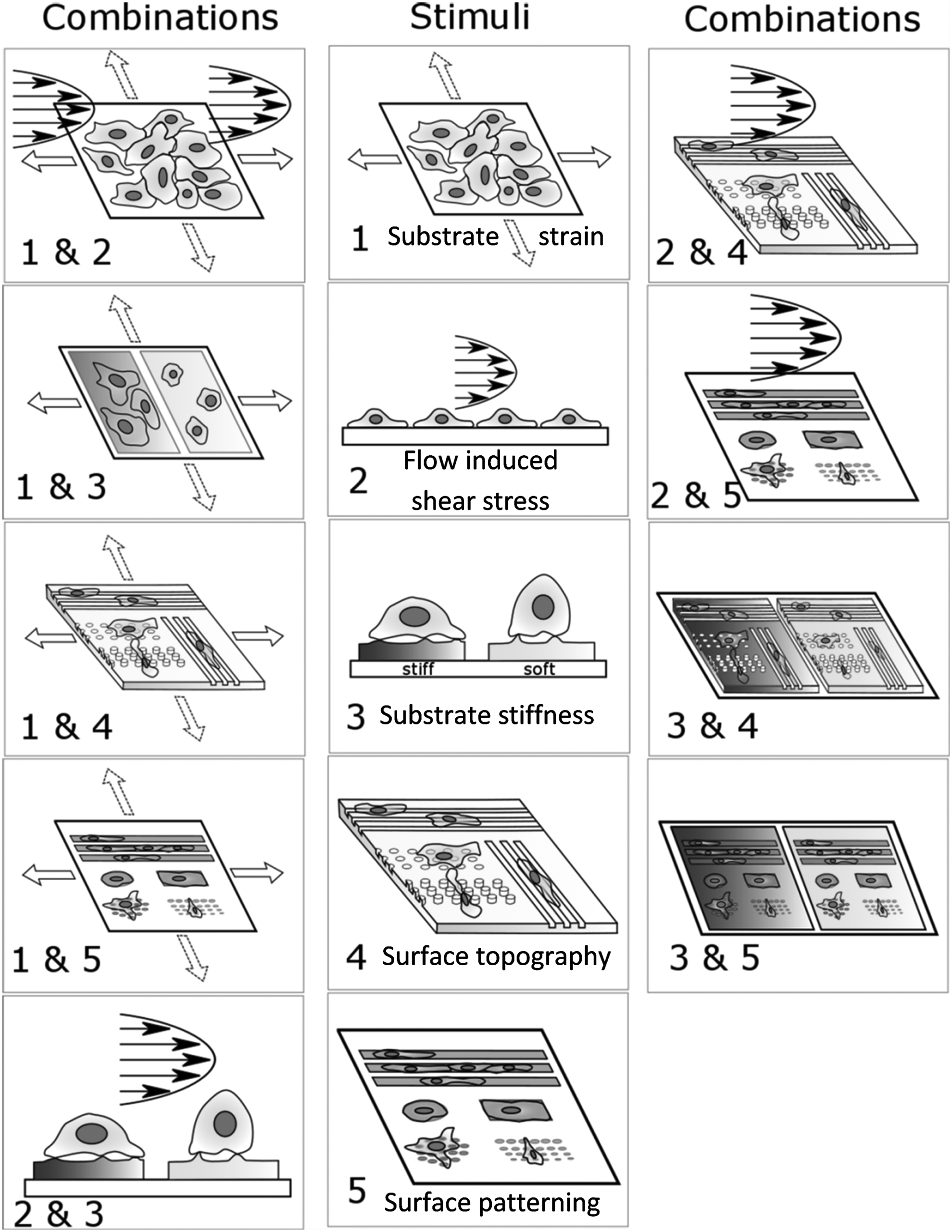
A schematic representation of various methods of applying mechanical stimuli to cells in vitro is shown here (middle column). Also shown are the schematic representations of the combinations of those mechanical stimuli covered in this review (side columns).
Cyclic substrate strain and flow-induced shear stress
Cyclic substrate strain and flow-induced shear stress are probably the most widely studied mechanical stimuli for cells, both individually and in combination. Cyclic strains are known to affect various cellular processes, such as alignment, proliferation, apoptosis, and differentiation. 27 They can be presented to cells in several variations such as isotropic, anisotropic, various magnitudes, and various frequencies. 27 Each variation can affect cells differently, thus explaining the large number of experimental studies. In a smaller subset of studies, static strains have also been shown to affect cells28,29 and from a mechanotransduction point of view, it has been argued that static strains can affect cells by unfolding molecules or structures that require forces that are sustained for a long duration to undergo structural changes. 30
However, due to the mechanotransduction machinery being dynamic, the forces being sustained often constitute an unlikely scenario; for example, as the cells migrate, they would break substrate adhesions, and the static strain-induced forces would be dissipated. This reasoning also has some support in previous studies, although the studies are rather old and in a very different context (in vivo study). 31
Flow-induced shear stress has been mostly applied to endothelial cells since several pathological conditions of the vasculature were linked to disturbed blood flows. 32 Similar to cyclic strains, shear stress can also affect various cellular processes. Some well-known effects of shear stresses resembling those resulting from blood flow in healthy vasculature are alignment of endothelial cells along flow direction and improvement of endothelium barrier function.
The cyclic strain–shear stress combination represents the dynamic stimuli present inside blood vessels and, hence, has been applied most often to endothelial cells.22,33–39 One way in which such stimulation was initially achieved was by seeding cells on the inner lining of cylindrical elastomeric constructs and applying flow with a pressure pulse similar to blood flow.33,34 Most approaches developed later have used uniaxially stretched flat membranes that allow for easier microscopic imaging, with the uniaxial strains in such systems representing the circumferential strains in blood vessels.22,35,36,39 The flow is usually applied to the cells on these membranes perpendicular to the strain direction, also as in blood vessels.35,36,39 More recently, the application of these two dynamic stimuli has been achieved by using micro3,4,39 and meso37,38,40 scale platforms. The small size and low reagent requirement advantages of such platforms have also been put to use in increasing the throughput of the experimental platforms.37,38,40
The two stimuli have been found to have synergistic effects on endothelial cells—for example, the simultaneous application of the two stimuli have been found to improve the endothelium morphology, leading to elongated cells and long, aligned stress fibers.33,34 However, the effects of the two stimuli are not simply additive, as was shown by Breen et al. by comparing the regulation of Intracellular Adhesion Molecule 1 (ICAM-1) and Vascular Cell Adhesion Molecule 1 (VCAM-1) by the two stimuli alone and in combination. 36
More examples where strains and shear stresses have been found to have beneficial synergistic effects include a study where the stimuli were effective in increasing the intracellular calcium in tenocytes 41 and another study where they enhanced the differentiation of mesenchymal stem cells into endothelial cells with the aim of blood vessel tissue engineering. 42 Very interestingly, it has been shown in microfluidic platforms called lung-on-a-chip3 [Fig. 2(iii)] and gut-on-a-chip4 [Fig. 2(iv)] that the combined strain and shear stress stimuli can enable cells to assemble into in vivo like structures as well as to better predict the effects of pharmaceuticals.

The application of combinations of mechanical stimuli to cells in vitro has been put to several uses. (i) Aligning cardiomyocytes by using attachment area patterning combined with the application of pathological strains has been used to develop cardiovascular disease models. 23 (ii) A combination of substrate stiffness and attachment area patterning has been applied to control the generation of neural networks in vitro. 112 Strain–shear stress combinations have been utilized in (iii) lung-on-a-chip 3 and (iv) gut-on-a-chip 4 platforms, where they elicit more in vivo like behavior from the respective cell types and provide a useful way for pharmaceutical testing.
Combined flow-strain stimulation platforms have been used to study pathological conditions as well—Amaya et al. found an upregulation of atherogenic genes in endothelial cells when the cyclic substrate strains and the pulsatile flow were not synchronized as they are in vivo. 22 Combining complex variations of the two stimuli have also shown interesting results, for example, combined application of anisotropic biaxial strains and shear stress at various orientations to the strains showed that a 0.55 Pa shear stress dominated over cyclic substrate strains in terms of endothelial cell alignment response. 37
A challenge that is often encountered in designing test platforms for combined strain-flow stimulation is the interaction between the stimuli. When applying only strains to cells, a reactive shear stress is always applied to the cells due to the relative motion between the substrate and the culture medium. 43 When strain and shear stress are applied simultaneously, the strain actuation generally also changes the confining volume of the flowing fluid, thus adding to the unintended shear stress. Computational modeling has been used to design systems to reduce these unintended effects of the strain actuation on the shear stress, or at least to quantify and characterize them well. 40
Cyclic substrate strain and hydrogel stiffness
Many cell types respond to cyclic substrate strain by aligning along a preferred direction, generally the lowest strain direction. This effect has been attributed to the cells' attempt to minimize the stress, strain, or elastic energy. 16 However, these studies were done on substrates that are much stiffer than natural tissue, such as lung or vasculature, which experience strains in vivo. It is known that the substrate stiffness is extremely important for cells as they constantly gauge their environment. 44 Gradients in stiffness are also known to determine the direction of cell migration, a process known as durotaxis. 45 Hence, applying tissue-specific stiffness and strain stimuli simultaneously to cells is important.
Applying strains to cells that are attached to substrates that are closer in stiffness to natural soft tissue has been achieved, in general, by placing cells on or in hydrogels of an appropriate stiffness that are attached to an elastic substrate. For cells cultured on hydrogels, ECM-coated polyacrylamide hydrogels have been commonly used46–48 ; whereas for culture within hydrogels, ECM hydrogels such as collagen49–51 or fibrin 52 gels have been generally used.
When cultured within hydrogels, cells see a significantly different attachment milieu compared with a substrate surface and this has been suggested to be responsible for the observed significant differences in cell behavior. 53 The nutrient transport to the cells from the culture media is also modified. 53 However, it is difficult to separate the effects of the stiffness and the additional factors. For simplicity, the studies where cells were cultured inside hydrogels and a strain was applied are considered in this section. For strain-stimulating cells within hydrogels, increasing the throughput of the experimental platform has also been reported.54–56
For cells cultured on hydrogels and stimulated with cyclic substrate strains, the general observation in multiple studies has been that cells respond quite differently to the strains when compared with cells cultured on much stiffer silicone surfaces.47,57,58 On stiff silicone membranes, cells usually readily align perpendicular to the major strain direction; whereas on hydrogels, the response is less prominent for a low stiffness47,58 and even contrary in certain instances (cells align along the major strain direction). 57 It has also been shown that strain stimulation can make cells display a more spread morphology on low stiffness hydrogels, where they show low spreading under static conditions. 47 As hydrogels are stretched and relaxed, water moves out and back into the hydrogel network. The forces caused by this movement have been shown to be responsible for the rupture of epithelial cell layers, which was earlier believed to be due to the breaking of cell-cell or cell-ECM bonds as a result of the stretching 48 [Fig. 3(iii)].
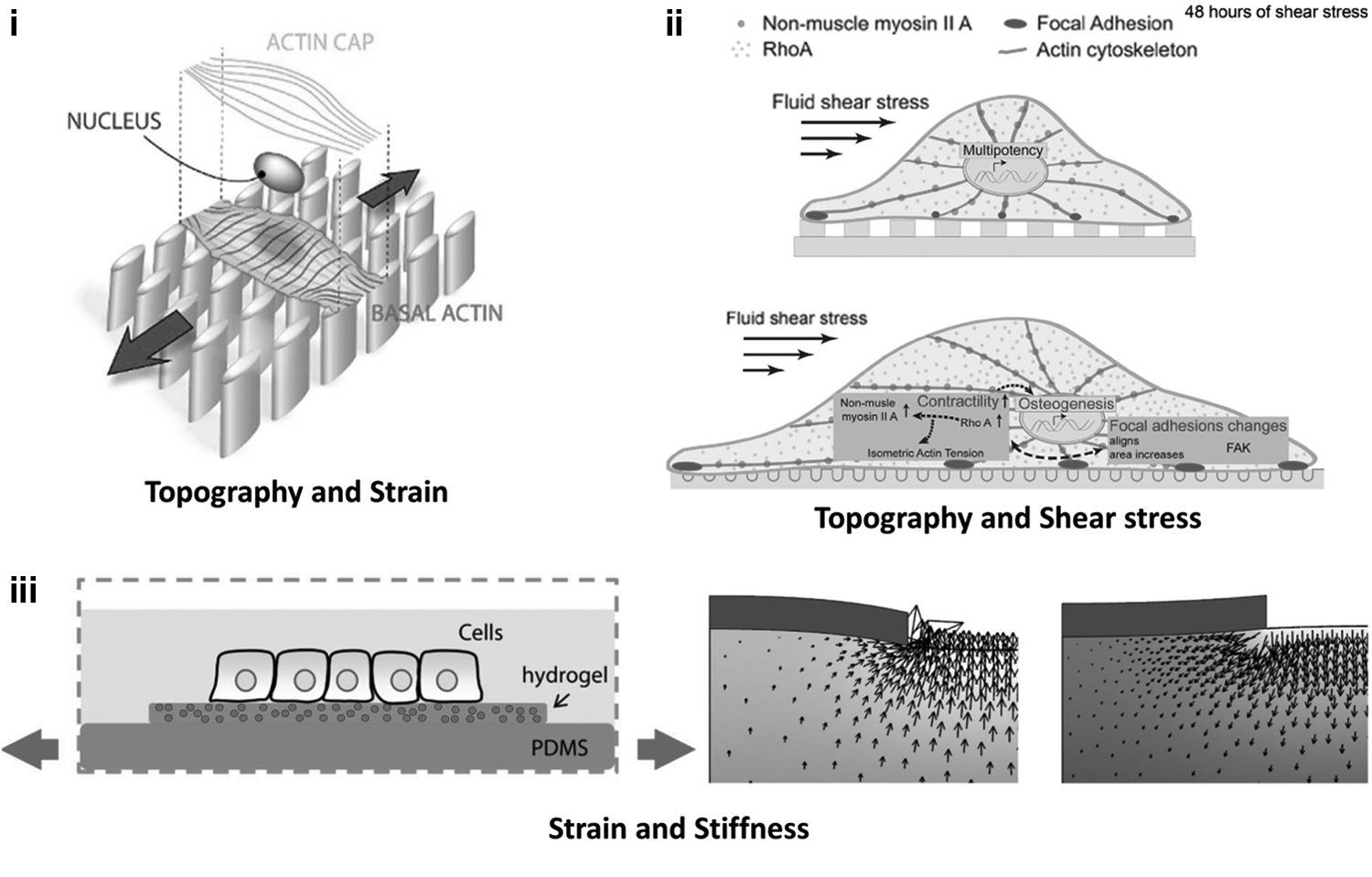
Even though the responses of cells to individual signals are often well understood, the combination of different mechanical stimuli can lead to conflicting cellular responses. Stimulating cells in vitro with combinations of mechanical stimuli has provided important insights. Examples include: (i) Topography combined with strains has been used to show that basal actin responds to topography; whereas the actin cap, which goes over the nucleus, responds to strains. 75 (ii) Topography-modulated cell attachment area and the corresponding modulation of cell contractility have been shown to have the potential to regulate mesenchymal stem cells (MSC) differentiation in the presence of flow-induced shear stress. Flow-induced shear stress increases the contractility of MSC and drives them toward osteogenesis on a topography where cells are more spread and more contractile (bottom), whereas a topography that reduces their attachment area and contractility can keep them multipotent in the presence of shear stress (top). 99 (iii) Strains applied to a hydrogel providing a physiological stiffness for cells revealed that the hydrodynamic forces from the fluid movement caused by the strains can have significant effects on cells. 48
In the case of cells cultured within hydrogels, the application of strains generally leads to better tissue-specific phenotypes and arrangements. For example, osteogenic differentiation was shown to be induced in mesenchymal stem cells, 59 valve interstitial cells were found to respond to anisotropies of strain and take on an active myofibroblast phenotype at high strain anisotropies, 50 contractility of smooth muscle cells was found to increase, 51 endothelial cells were found to form structures with lumen at the cost of sprout formation, 52 and tendon/ligament markers were shown to be upregulated in mesenchymal stem cells. 60 Another effect commonly observed was the higher mechanical strength of strained cell hydrogel constructs.51,56 When nonphysiological strains were applied, such as equibiaxial strains applied to smooth muscle cells that reside in an anisotropically strained environment in vivo, the loss of the healthy phenotype was observed. 49
In the studies with cells within hydrogels, the experiments were designed around providing a three-dimensional environment around the cells, resembling their in vivo environment. The stiffness was not really controlled, although the stiffness probably played a much more significant role as evidenced from the high interaction observed between the cells and the hydrogels in the form of hydrogel compaction 61 and rearrangement 50 in several studies. This would not have been possible in a three-dimensional porous scaffold of a much higher stiffness.
The studies combining hydrogel stiffness and strain show that cell response to strain on hydrogels can vary significantly from response on stiff elastomeric substrates that were normally used previously. New forces from fluid squeezing in and out of the hydrogel network due to strains need to be considered as well. 48 Since cells are able to deform the hydrogels, techniques such as traction force microscopy offer a way to quantify the cell-substrate interactions. 48 Putting cells within the hydrogels permits even greater interaction with the ECM network. When comparing the effects of strains on cells on hydrogel with those on stiff silicone sheets, an important consideration is the transfer of the strains to the cells, 46 although it is not always reported.
Cyclic substrate strain and surface topography
Substrate surface topography has long been known to affect cell attachment. Surface topography can increase the surface area for better cell interaction and attachment 62 or can prevent microorganisms from attaching (anti fouling), 63 organized structures such as grooves can orient cells a certain way (contact guidance), 64 or topography can direct stem cell differentiation.65,66 Even though surface topography has many applications for the control of cellular behavior, not enough is known to always be able to predict how a topography would affect certain cells.
High-throughput techniques are now being used to explore the effects of a large number of topographies. By accumulating large quantities of data, hopes are that a predictive link between topography and cellular action can be made. 66 When combined with strains, topography can affect the way substrate strains are transferred to the cells and in the case of contact guidance of alignment, oppose the alignment response of cells to the strain. To test for these effects, topographical features have been created on stretchable silicone sheets.
The general observation has been that microgrooves (1–20 μm width) can keep cells aligned along them even when the strain is along the groove direction, hence trying to make the cells align perpendicular to the grooves. 67 This has, in fact, been used to apply tensile strains to cells such as ligament fibroblasts in the direction of alignment, which resembles their in vivo condition, but is otherwise not attainable in vitro since the strains direct them to align perpendicularly. 68 The strain along the alignment direction stimulus also resembles the in vivo condition of smooth muscle cells and has been found to increase smooth muscle cell markers in mesenchymal 69 and neural crest 70 stem cells. However, it has been suggested that there is a strain threshold after which the strains dominate the alignment response—for example, Loesberg et al. found this threshold to be at 8% strains. 71
In contrast, nanogrooves (300–600 nm width), which can also guide cells to align along them, could not sustain the alignment if the strain was applied along the grooves and directed the cells to align perpendicular to the grooves.72,73 The strain threshold for dominating topography-directed alignment along nanogrooves was found to be much lower (3%) than for microgrooves. 72 Wang et al. showed that as the nanogroove sizes increase, they are able to resist the strain stimulus for a longer duration, but given sufficient time, strains can always dominate topography-driven alignment. 74
In all the studies, if strain and topography were applied perpendicular to each other, they had a synergistic effect on the alignment of the cells. A mechanistic understanding of the competition between strain and contact guidance has only recently started to emerge with the demonstration that the basal actin responds to the contact guidance and the perinuclear actin cap, which is directly connected to the nucleus, follows the strain avoidance response to strain and guides the nucleus alignment with it 75 [Fig. 3(i)].
Few studies have combined strain and topography where the topography was not used to getting contact guided alignment. In one such example, when the topographical cue was micropillars, the addition of strain changed how the cells attached to pillars and regions between pillars. 76 The strain also caused larger changes in the expression levels of genes related to the cellular mechanotransduction machinery than the presence of the pillars alone. 76 In another example, the topography presented to cells was micropillars and the strain was applied only to the pillars by magnetically actuating them. It was shown that the pillar strains slowed down the cell migration. 77
Cyclic substrate strain and attachment area patterning
In addition to topography, another method that has been used to guide cell attachment is the chemical patterning of the substrate surface area into parts where cells can attach and parts where they cannot attach. This has generally been achieved by microcontact printing, where a silicone stamp of the pattern is obtained by soft lithography microfabrication and the pattern is stamped on the substrate after adsorption of the patterning molecule (usually an ECM protein) on the stamp. 78 By patterning the attachment area for individual cells, it has been shown how the pattern shape that determined cell shape can determine cell death 79 or differentiation of stem cells. 80
Arrays of cell adhesive islands that are smaller than cells have been used to differentiate between the effects of attachment area and cell shape 79 and also to determine the limits of attachment point size and distance between them that are needed for cells to attach and spread. 81 Rows of attachment areas have also been used to align cells in a way that is similar to contact guidance by microgroove topographies. 82 Gradients in the availability of attachment areas have been used to study the corresponding determination of cell migration, a process known as haptotaxis. 83
The number of studies stimulating cells with both attachment area patterning and cyclic substrate strain is few. Two of these studies used parallel patterned lines along which cells showed alignment. On the application of strains at various angles to the patterned lines (0°, 45° and 90°), the cells mostly ignored the alignment stimulus of the strain and remained aligned along the patterned lines.82,84 However, in one of these studies, when the strains were applied along the patterned lines, the cells aligned along an intermediate angle between the dictated alignment by the strains and the surface patterning. 82
Two other studies used patterning to align and assemble cells into arrangements resembling their in vivo condition and then applied strains either along the same direction as the in vivo strain direction for them or along the perpendicular direction, which represented a pathological condition23,24 [Fig. 2(i)]. In one study, the cells used were valvular endothelial cells and it was shown that the application of a pathological condition of a 20% strain perpendicular to the cell alignment direction drove the cells to a mesenchymal phenotype (endothelial to mesenchymal transition). 24 In the other study, using cardiomyocytes, the application of pathological strains perpendicular to the alignment direction led to a disruption of the cell layer architecture. 23
And lastly, in a study using patterning to control individual cell shapes, on the application of strains, 85 it was found that strain increases the amount of stress fibers and focal adhesions in radially symmetric cell shapes (circular, square); whereas in bipolar (rectangular) cells, there is a higher rearrangement of the stress fibers and focal adhesions. 85
Flow-induced shear stress and hydrogel stiffness
This combination of stimuli, just like strain-flow combinations, is very relevant for blood vessels and has mostly been applied to endothelial cells. The effects of shear stress have been widely studied for endothelial cells, but these studies were generally performed on cells that attached to very stiff substrates such as glass. 86 It is known that endothelial cells are affected by the stiffness of their environment. 87 It is also known that blood vessels stiffen with age and in some pathological conditions. 88 Hence, it is likely that substrate stiffness will have an effect on the response of endothelial cells to shear stress.
This was found to be the case from multiple studies where shear stress was applied to endothelial cells that were cultured on substrates having a more tissue-like stiffness. Galie et al. cultured endothelial cells on polyacrylamide gels of various stiffnesses and applied various shear stresses to them. 89 They observed that on lower stiffness hydrogels, the cells were more sensitive to shear stress in terms of spreading, although they were less sensitive in terms of alignment. 89 They also observed that the cells could better use shear stress to modulate the effect of the inflammatory cytokine tumor necrosis factor alpha on a lower stiffness hydrogel. 89 Kohn et al. observed that endothelial cells responded to shear stress with a more elongated morphology and tight cell-cell contacts on substrates with lower stiffness when compared with a glass substrate. 90
Flow-induced shear stress and substrate surface topography
This combination of stimuli has been applied to cells in vitro to study various useful processes, such as the competition between the alignment responses to each stimulus, achieving tightly bound cell layers that resist detachment under high shear flows, directed cell migration, preparing cell layers predictive of the in vivo condition, and uncovering mechanotransduction pathways.
In a way similar to the topography-strain competition in aligning cells, a larger-sized groove topography (>800 nm pitch) was found to be able to retain endothelial cell alignment when opposed by shear stress. 91 However, unlike in the strain-topography case, smaller topographies were not completely dominated by the shear stress and resulted in a loss of alignment to either stimulus. 91 When the two stimuli directed the cells to align in the same direction, the effects were synergistic. 91 Tightly bound endothelial cell layers resisting detachment by shear stress are of utility for cardiovascular implants such as vascular grafts92,93 or devices with an antithrombotic surface.94,95 Topography-induced alignment along the flow92,95 and even just the presence of topographical features aligned along the flow direction 93 have been shown to achieve this, although topography aligning cells perpendicular to the flow were found to be more useful in withstanding very high shear stresses (8 Pa). 94
Aligned topographies can be used to provide direction to migrating endothelial cells, irrespective of the flow direction, 96 and such directionality can be used to advantage in enhancing wound healing. 97 Combined topography and shear stimulation has also been used to improve the alignment and tight junction formation in kidney epithelial cells, thus providing a better in vitro model of renal tissue. 98 And lastly, using topography cues to control the cell shape-modulated contractility of mesenchymal stem cells (MSC) and applying shear stress to further affect their contractility, the regulation of the differentiation of MSC has been shown 99 [Fig. 3(ii)].
A challenge in combined topography-shear stress stimulation is the estimation of the actual shear stresses experienced by the cells since topographical features can potentially disturb the flow. Hence, computational modeling has been used to study shear stress variations caused by topography as well as by the experimentally determined surface of a cell layer resting on the topographical features. 100
Flow-induced shear stress and attachment area patterning
Only a few studies have been reported where combinations of these two stimuli were applied to cells. You et al. studied the intracellular calcium signal of osteoblast cells attached to an array of adhesive islands in response to applied shear stress. Based on the calcium signals, they observed that the cells that attached to the islands were more sensitive to the shear stress than those that attached to nonpatterned substrates and that increasing the cell spreading area by moving the islands apart reduced this sensitivity to shear stress. 101 Hsu et al. showed by using a combination of collagen patterning and shear stress that up to a certain shear stress, cells move in the direction of higher collagen density (haptotaxis) even if it is against the flow, but at a higher shear stress mechanotaxis takes over and cells migrate in the direction of the flow, ignoring the collagen densities. 102 In addition, shear stress has been used to determine the strength of binding between cells and patterned adhesive islands.103,104
Hydrogel stiffness and substrate surface topography
The number of studies reported for this combination of stimuli is quite small as well. Schulte et al. varied stiffness and topography for a polyethylene glycol hydrogel and found that the presence of microgroove topography turns the normally cell nonadhesive hydrogel slightly cell adhesive, and this adhesion is stronger for hydrogels of a lower stiffness. 105 Lu et al. explored various topography and stiffness combinations to find an optimum substrate for maintaining the stemness of mouse embryonic stem cells. 106 Wong et al., 107 Wu et al., 108 and Jeon et al. 109 also looked at stiffness and topography combinations but they used materials of a nonphysiological stiffness. Still, differences in the cellular responses were observed.
Hydrogel stiffness and attachment area patterning
Combined stiffness and attachment patterning has been used to identify the forces that are generated by cells on spreading 110 and on being exposed to a stiffness gradient, 111 both using the detection of hydrogel deformations produced. The combination of these two stimuli has also been investigated for application in neural tissue engineering, where the patterning of islands and lines was used to organize neurons into networks (with cell bodies localizing to the islands and the dendrites localizing to the lines) and the stiffness was found to control the speed and quality of network formation (faster arrangement on a lower stiffness substrate, but more synapses and action potentials on a stiffer substrate) 112 [Fig. 2(ii)].
Another application of the combined control of stiffness and patterning was shown in assembling breast adenocarcinoma cells into hollow structures resembling epithelial structures in vivo. 113 This was achieved by comparing the combined effects of adhesive islands and substrate stiffness, which showed that lower substrate stiffness and far apart adhesive islands reduced the cell-ECM interaction.
Combinations of more than two stimuli
Since many mechanical stimuli are coupled, it is not always possible to apply a particular mechanical stimulus without applying another—for example, when strain is applied to cells that are attached on a substrate surface, reactive shear stress is applied to them as well and this is caused by the relative motion between the substrate and the medium. So, when applying combinations of two mechanical stimuli, more stimuli can actually be present. However, it is not always straightforward to vary or control these unintended stimuli. A platform has been reported where multiple mechanical stimuli can be intentionally and controllably varied. Galie and Stegemann reported a platform to apply interstitial flow-induced shear stresses and cyclic substrate strains simultaneously to cells cultured in a compliant hydrogel (stiffness stimulus). 114 They later used it to study the effects of the stimuli on cardiac fibroblast activation. 115
Closely mimicking the in vivo condition would require combining still more stimuli. The cells need to be put on or in a substrate with the native tissue stiffness, provided attachment cues to keep them aligned as in the tissue, it needs to be ensured that the same levels of cell-cell and cell-ECM interactions are present as in the native tissue, and finally, the appropriate dynamic mechanical stimuli of stretch and shear stress have to be applied. In addition, the substrate should be modifiable (degraded or rearranged) by the cells, allow movement of nutrient and waste products, and offer migration or positioning cues that are similar to the native tissue. Controlling all these parameters at the scale of the cells might be challenging due to the limits of fabrication, but the better the cellular interpretation of the mechanical cues is understood, the better are the chances to substitute a harder-to-implement stimulus by a simpler one.
Discussion and conclusion
The application of combinations of mechanical stimuli to cells has provided a number of insights on how opposing stimuli compete, how supporting stimuli produce synergistic effects that might not be directly additive, how one stimulus can sensitize cells toward another stimulus, how unintended stimuli might appear as side effects of the intended ones, and how the combined stimuli can produce more in vivo like behavior from cells in vitro. These effects would have been impossible to predict due to our incomplete understanding of the mechanical workings of cells. In addition, the combinations of stimuli have been used to better understand the workings of the mechanotransduction machinery in complex mechanical environments.
In future, many more new insights can be expected as systematic studies combining multiple stimuli are continued to test the effects of various magnitudes, directionality, frequencies, and dosage of the stimuli. Increased application of optimized, combined mechanical stimuli for tissue engineering and pharmaceutical discovery can also be expected.
High-throughput platforms can speed up this discovery process, and, hence, more of them could be expected to be employed. With increasing knowledge, the simplest in vitro systems for each application could be identified, for example, if a topography and substrate stiffness can elicit the same response from cells as putting the cells within a hydrogel and applying strains, the former stimulation could be selected for its ease of application. A better understanding of mechanotransduction mechanisms is also likely to emerge from such studies. Mathematical and computational modeling have already been used to predict the mechanical stimuli in complex conditions and to predict cell response to simple stimuli. With the advanced knowledge from combined stimulation studies, the models can be updated to provide them with higher predictive ability. Thus, we conclude that the in vitro stimulation of cells with combinations of mechanical stimuli is a useful approach and systematic studies should be continued.
Footnotes
Disclosure Statement
No competing financial interests exist.