
Abstract
A major hurdle in the field of tissue engineering and regenerative medicine remains the design and construction of larger (> 1 cm3) in vitro tissues for biological studies and transplantation. While there has been success in creating three-dimensional (3D) capillary networks, relatively large arteries (diameter >3–5 mm), and more recently small arteries (diameter 500 μm–1 mm), there has been no success in the creation of a living dynamic blood vessel network comprising of arterioles (diameter 40–300 μm), capillaries, and venules. Such a network would provide the foundation to supply nutrients and oxygen to all surrounding cells for larger tissues and organs that require a hierarchical vascular supply. In this study, we describe the different technologies and methods that have been employed in an effort to create individual vessels and networks of vessels to support engineered tissues for in vivo and in vitro applications. A special focus is placed on the generation of blood vessels with average dimensions that span from microns (capillaries) to a millimeter (large arterioles). We also identify major challenges while exploring new opportunities to create model systems of the entire vascular tree, including arterioles and venules.
Introduction
T
Current methods to circumvent diffusion limitations involve either the vascularization of tissue constructs with small capillaries, or the generation of conduits that serve to augment the transport of nutrients and waste products.14,15 An alternate approach to achieve greater success in the generation of thick tissues could be to mimic more accurately the composition and construction of the vascular network found in vivo. During embryogenesis, the formation of a network of blood vessels occurs through both angiogenesis and vasculogenesis.16–19 Vasculogenesis is the de novo formation of a primitive network of blood vessels by in situ differentiation of endothelial cells from the mesoderm.20,21 Angiogenesis is the growth of new capillary vessels from an existing network of blood vessels. 17 These processes are initiated primarily by the Vascular Endothelial Growth Factor (VEGF) signaling pathway.17,20,22–24 The in vivo formation of blood vessels in tissue organs through vasculogenesis and angiogenesis ultimately leads to a hierarchical network of vessels that includes arteries, arterioles, capillaries, venules, and veins.
Over the past few years, several newer methods, such as bioprinting and soft lithography, have been used to mimic the hierarchical structure of vessels in vitro for biological experiments and in vivo applications.1,15,25 While recreating some of the necessary structure, these methods do not result in a living dynamic network of vessels capable of remodeling in response to changing metabolic demands. Nonetheless, the generation of functional and thick engineered tissues requires a comprehensive understanding (modeling and experimental validation) of oxygen transport and anastomosis between the tissue graft and host vasculature (in vivo transplant case) or artificial inlets and outlets (microfluidic platforms). The satisfaction of these two conditions could potentially solve the transport problem related to the generation of thick tissue constructs. In this review, we focus on the creation of vessels and vessel networks to enhance transport of nutrients and waste products, as opposed to overcoming the low solubility of oxygen in water. We first describe the process of vessel formation in vivo, and then review the methodologies used for the design of blood vessels to support engineered tissues. We also outline current challenges and opportunities to tissue engineering the entire vascular tree to respond to the increasing clinical need for thick, complex, and cell-dense tissues and organs.
Vascular development during embryogenesis
The cardiovascular system is the initial organ formed during embryogenesis.26,27 For continued growth and development of the embryo, the cardiovascular system has to be fully functional. At the onset of a beating heart, the vascular system will primarily serve as a conduit for the transport and distribution of nutrients and oxygen to the developing embryo and for the removal of waste.
Vasculogenesis
In the developmental phase of the embryo, vasculogenesis occurs first through the formation of blood islands in the yolk sac followed by the differentiation of endothelial cell precursors (angioblasts).16,20,21,28 Blood islands originate through gastrulation, from the migration of undifferentiated mesodermal cells in the yolk sac between the ectodermal and endodermal cell layers (Fig. 1a–c). These mesodermal cells differentiate into hemangioblasts, which form blood island aggregates within the yolk sac. 16 The continued maturation and differentiation of the blood island aggregates and their subsequent fusion leads to the formation of primary capillary plexuses and a primitive network20,21 (Fig. 1d, e). Essentially, the inner cells of the aggregates become hematopoietic precursors, whereas the outer cells develop into endothelial cells. These newly differentiated endothelial cells will anastomose and form a primary capillary plexus thus laying down the initial network for blood circulation. At the molecular level, VEGF, basic fibroblast growth factor (bFGF), VEGF receptor 2 (VEGFR2), and FGF receptor (FGFR) play important roles in the differentiation of pluripotent embryonic stem cells into hemangioblastic cells.
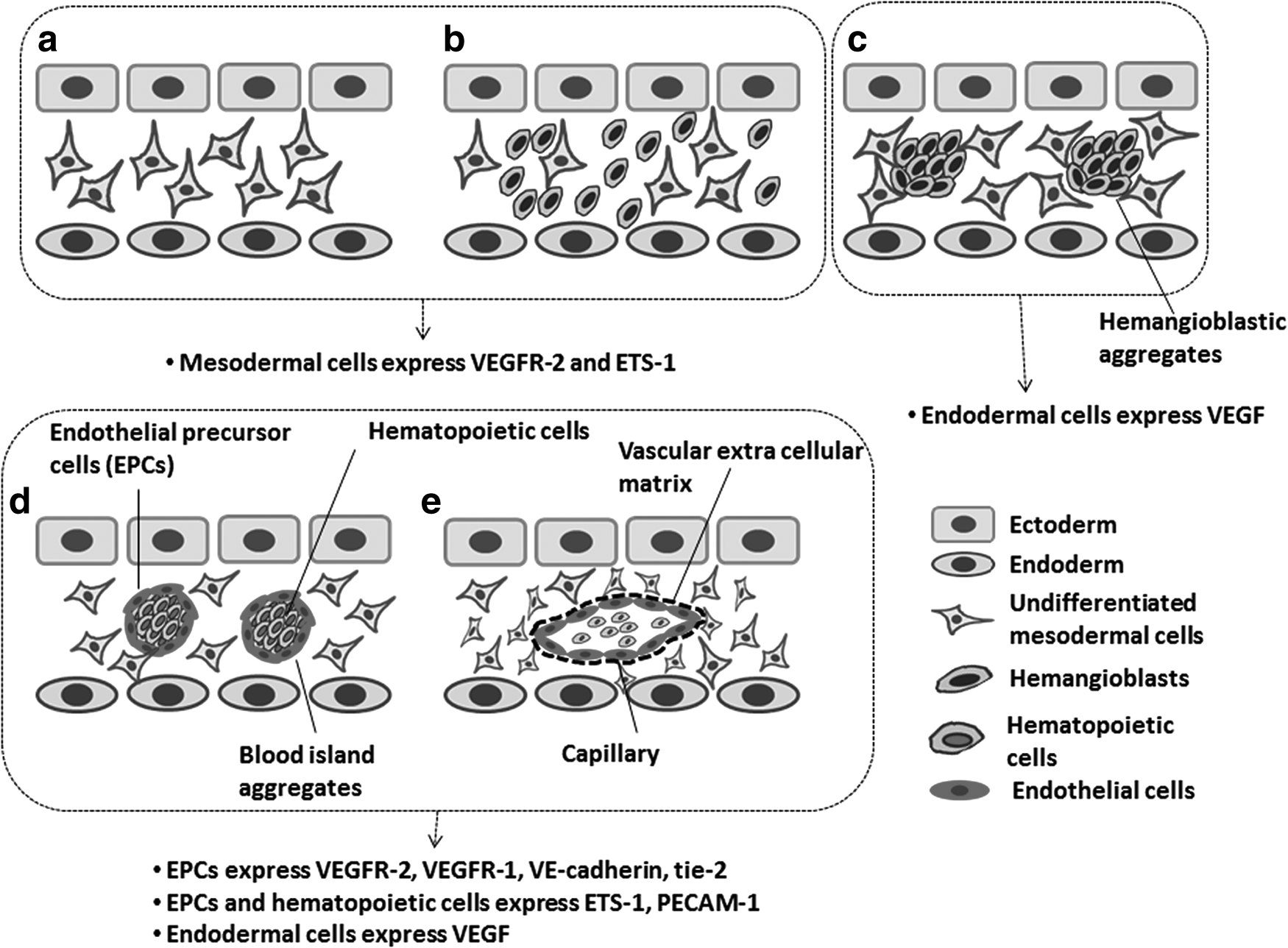
Steps that illustrate the formation of a capillary plexus during vasculogenesis.
Angiogenesis
Angiogenesis was first described in 1787 by a British surgeon, John Hunter, and today describes the sprouting, budding, branching, bridging, intussusception, and remodeling of new vessels from preexisting ones29,30 (Fig. 2a–d). The permeability of the new vessels is an essential characteristic in the angiogenic process. The presence of VEGF, also known as vascular permeability factor in the microenvironment stimulates the permeability of the neovascular network thus allowing the leakage of blood proteins, including fibrin, into the microenvironment. 20 This process favors the generation of a temporary scaffold required for migrating endothelial cells and the formation of tubes (tubulogenesis). Angiogenesis involves the disruption of interendothelial cell junctions and the destabilization of mature vessels.19,29,31–33 This normally entails the removal of perivascular stromal cells from the capillaries (Fig. 2e, f). The disruption of mature vessels includes the degradation of matrix proteins, which also liberates growth factors such as VEGF and bFGF from the surrounding extracellular matrix. 18 At this point of the process when physical barriers are degraded, the microenvironment is conducive to the migration and proliferation of endothelial cells. Endothelial tip cells (a specialized endothelial cell) are leader cells during vessel sprouting; they are located at the forefront of vessel buds, and are able to probe the adjacent microenvironment and direct migration through chemotaxis following spatial morphogen (e.g., VEGF) gradients34–36 and mechanical stimuli (i.e., interstitial flow).37–39
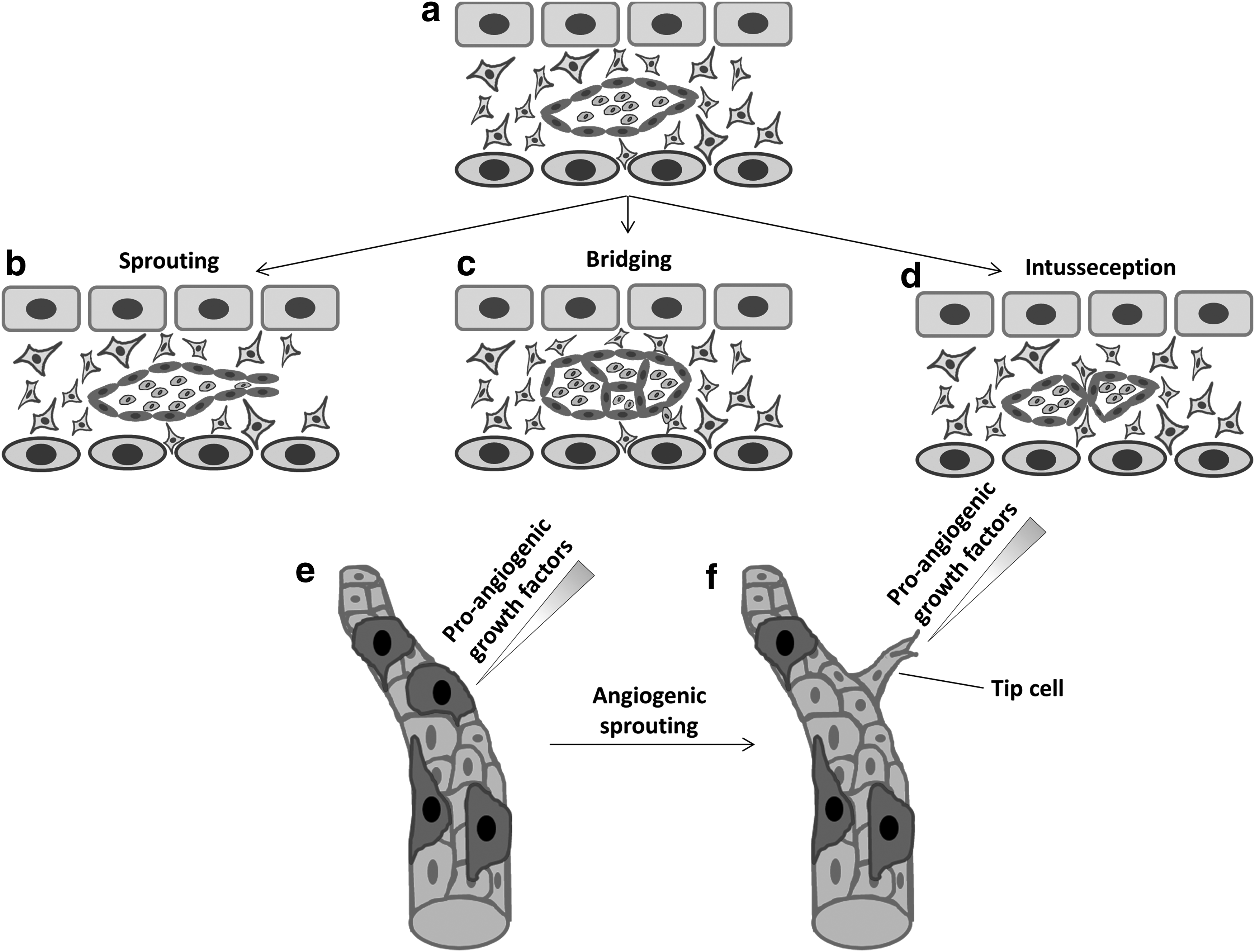
Blood vessel formation through angiogenesis.
Hierarchical Network of Vessels In Vivo
Molecular regulation of vessel growth, maturation, and stabilization
To match the demand of a growing organ, the nascent vasculature formed during vasculogenesis and angiogenesis must mature into a network capable of sustaining the metabolic demand of the organism. This process relies mainly on reshaping the already existing networks into a hierarchical network of larger vessels as well as capillaries. 31 However, for the process to be complete and ensure stable vessels, the recruitment and differentiation of perivascular stromal cells, such as pericytes and smooth muscle cells, and the generation of an extracellular matrix are major requirements.40–42
The first molecular pathway employed by endothelial cells to recruit mural cells for vessel stabilization is the platelet-derived growth factor (PDGF) signaling pathway.43,44 Endothelial cells recruit mural cells (pericytes) expressing PDGFRβ by secreting PDGF-BB. This process promotes the migration and proliferation of mural cells during vessel maturation and ensures long-term function and stability. The presence of VEGF may upregulate PDGF-BB in the recruitment of perivascular cells. 33 The contact between endothelial cells and pericytes is regulated through the S1P1/EDG1 signaling pathway in endothelial and mural cells. 42 Even though this pathway serves to localize N-cadherin molecules on the abluminal surface of endothelial cells, therefore strengthening the contact between the endothelial cells and pericytes,31,42 it has been shown to also promote mural cell recruitment potentially downstream of PDGF-BB signaling. The vasculature's maturation process also involves the Angiopoietin-1(Ang1)/Tie2 signaling pathway. 32 Ang1 is known to activate the Tie2 receptors on endothelial cells; this process strengthens the adhesion between endothelial cells and pericytes while reinforcing the interendothelial junctions. 45 Recently, Ang1 has also been shown to clearly affect the baseline diameter of vessels as well as vessel density during the development of the vasculature. 46
Hemodynamic regulation of vessel growth, maturation, and arteriogenesis
The remodeling process of the vasculature is highly dependent on hemodynamic factors, such as blood flow and pressure.21,44,47 The onset of blood flow creates physical forces (shear, normal, and circumferential stresses) that are sensed by the endothelial cells, as well as perivascular cells (Fig. 3) Endothelial cells possess mechanoreceptors that can detect as well as respond to shear stress, including, but not limited to, integrins, receptor tyrosine kinases, ion channels, G protein-coupled receptors, cell–cell junction molecules (PECAM-1), adherens molecules, and the membrane lipid bilayer.48,49 The mechanotransduction process begins with the deformation of the cytoskeleton of endothelial cells subjected to fluid-derived mechanical stresses (e.g., shear stress, pressure), which is rapidly transmitted to the interior of the cell body.48,50,51 The third step in this process consists of the conversion of the mechanical stress into chemical information followed by chemical signaling.
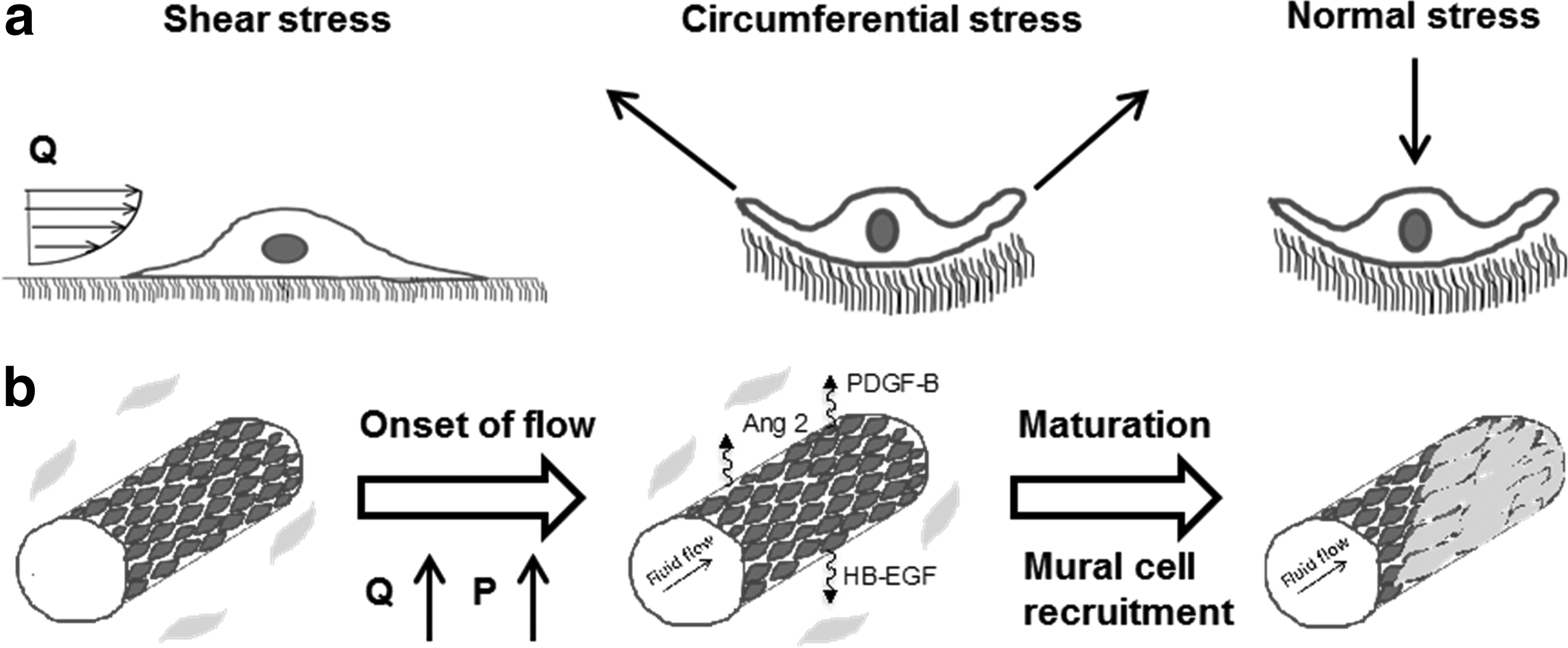
Illustration of capillary maturation into arterioles.
The ability for endothelial cells to sense and respond to mechanical forces impacts the maturation process of blood vessels.44,52,53 For instance, low shear stress levels (<10 dynes/cm2) can help promote the migration, differentiation, and proliferation of vascular smooth muscle cells 54 through endothelial cell expression of molecules that promote growth factors such as PDGF-BB and VEGF. PDGF-BB then contributes to the proliferation and recruitment of perivascular cells toward the endothelial cells through chemotaxis.43,53 Previous studies have also shown that shear stress is directly related to arterial development postnatally, whereas smooth muscle cells in the adult vasculature directly respond to the increase in hemodynamic pressure in the vessel. 55 This is especially the case during arteriogenesis of collateral vessels56,57 of patients with coronary artery disease (CAD). Collateral vessels in humans are present at birth and have the ability to attenuate the extent of myocardial ischemia and thus minimize myocardial cell death. Blood flow in collateral vessels of patients with stable CAD has been associated with reductions of myocardial infarctions and with increased survival rates. The growth of collateral vessels occurs mainly through arteriogenesis, where the growth of arterioles is stimulated by hemodynamic forces that emanate from blood flow. Twenty-four hours after occlusion of an artery, the wall of the collateral vessels is exposed to all three hemodynamic forces.57,58 These forces are thought to activate the endothelial cells to release soluble and diffusible molecules that eventually lead to the recruitment and proliferation of smooth muscle cells that result in vessel maturation and arteriogenesis. 55
Blood vessel structure is highly correlated to the blood flow pattern, and thus hemodynamic forces throughout the vasculature, which can explain both the structural and functional differences among vessels in most organisms. For instance, the larger shear stresses and pressures in arteries with pulsatile flow 12 are known to correlate with more elastin in the extracellular matrix (ECM) (to increase the compliance) and multiple smooth muscle cell layers to dampen the circumferential stretch experienced by these vessels. This is quite different in arterioles, where less elastin is present and a relative abundance of smooth muscle cells providing the ability to control flow without the need to absorb a pressure pulse.59,60
ECM in blood vessels
The ECM in blood vessel wall is often overlooked when referring to engineering thick tissues. The ECM plays a crucial role in regulating growth factor signaling at the cellular level. The basement membrane lies right underneath the endothelial cells, and its source has generally been thought to be the endothelial cells themselves, but growing evidence also supports a role by mural cells such as pericytes.43,61,62 The basement membrane is primarily comprised of laminins and other connective tissue proteins such as collagen type IV and perlecan. As the vasculature matures (from capillary to higher hierarchical structures, such as the artery, during embryonic development), the ECM remodels to ensure proper function. For instance, the increase in blood pressure and cardiac output coincides with an increase in production in elastin and collagens by stromal cells in the tunica media of vessels. Elastin and collagen production persists until the cardiac output becomes stable. Thus, ECM deposition is a major determinant in the creation of the vessel network hierarchy.40,41,63
In Vitro Vasculature Development
Arteries (diameter 0.4–8 mm)
The vessels present in our body vary in size from a diameter of several centimeters (aorta, vena cava) to as small as five microns (capillary). 12 For the past several decades, researchers have attempted to recreate vessels with similar size characteristics and functionality. Major advances have been made in the construction of larger vessels primarily for transplantation purposes. The steady increase in vascular diseases in developed countries around the world largely contributes to the use of biological vascular grafts (autografts, allografts, and xenografts) of venous and arterial vessels in surgical bypass procedures.
Synthetic materials have been utilized as viable alternatives to meet the demand for vascular grafts. Vascular prostheses made with materials such as polyethylene terephthalate known as Dacron, expanded polytetrafluoroethylene (ePTFE), and polyurethane have demonstrated excellent in vivo performance. This success is limited to vascular grafts with a diameter larger than ∼8 mm.64,65 Smaller diameter (<6 mm) vascular grafts in carotid and common femoral arteries have not been successful 64 due to surface thrombogenicity and intimal hyperplasia. Surface thrombogenicity impairs vessel patency and is due to the lack of endothelial cell coverage of the inner surface of the vessel. These hurdles have stimulated the use of chemicals and anticoagulants, such as 66 hirudin, dipyridamole, tissue factor pathway inhibitor, and nonthrombogenic phospholipid polymers to reduce the thrombogenicity of the vascular grafts. 67 Intimal hyperplasia is the cellular proliferation of stromal cells and generally arises at the anastomotic junction between the local vasculature and the prosthetic vessel. The use of venous cuffs between an artery and the synthetic graft has been shown to alleviate some of these problems. 68
Another type of successful artificial vascular conduit is the tissue-engineered blood vessel (TEBV), which has been commercialized by companies such as Cytograft. In the TEBV, a cell sheet is produced by culturing smooth muscle cells in cell culture plates with higher concentrations of ascorbic acid to promote the synthesis of collagen. 69 The formed cell sheets constitute the tunica media of the vessels and are subsequently wrapped around a porous cylindrical mandrel. The use of a porous mandrel is essential as it allows nutrient access to all cells during tissue maturation ex vivo. The adventitial layer of the vessel is constructed by wrapping a fibroblast cell sheet layer around the tunica media made of smooth muscle cells. Following maturation of the vessel for several weeks, the support mandrel is removed and the luminal surface of the vessel is seeded with endothelial cells. TEBV exhibits important characteristics of arteries such as the abundant ECM in the medial layer, even though the fiber orientation is different compared with medial fibers in native arteries.70,71
More recently, Fernandez et al. 72 were able to create in vitro human tissue-engineered human blood vessels with diameters between 400 and 800 μm. These vessels exhibited vasoactivity and were used to evaluate the toxicity of pharmaceutical drugs before preclinical animal studies. The vessels created in this study not only showed mechanical strength and stability as validated by a burst pressure above 1777 mmHg, but also contractile and ECM protein expression (collagen IV and laminin) over time when subjected to laminar flow perfusion.
Microvessels (diameter 10–50 microns)
The formation of a network of capillaries or microvessels within an ECM in vitro can be achieved through two major strategies. The first strategy entails mixing endothelial cells with stromal cells in a hydrogel, such as fibrin or collagen, while ensuring that all cells within the microtissue have access to oxygen and other nutrients. The hydrogel is polymerized using either thrombin (fibrin) or by raising the temperature (collagen). This can be achieved in either a standard well plate or within a microfluidic chamber.14,73–75 A stable capillary network self-assembles over the ensuing 4–7 days in a process that mimics vasculogenesis (Fig. 4).76–78 Once a stable vascular network has formed, an appropriate perturbation can induce angiogenic sprouting. 79
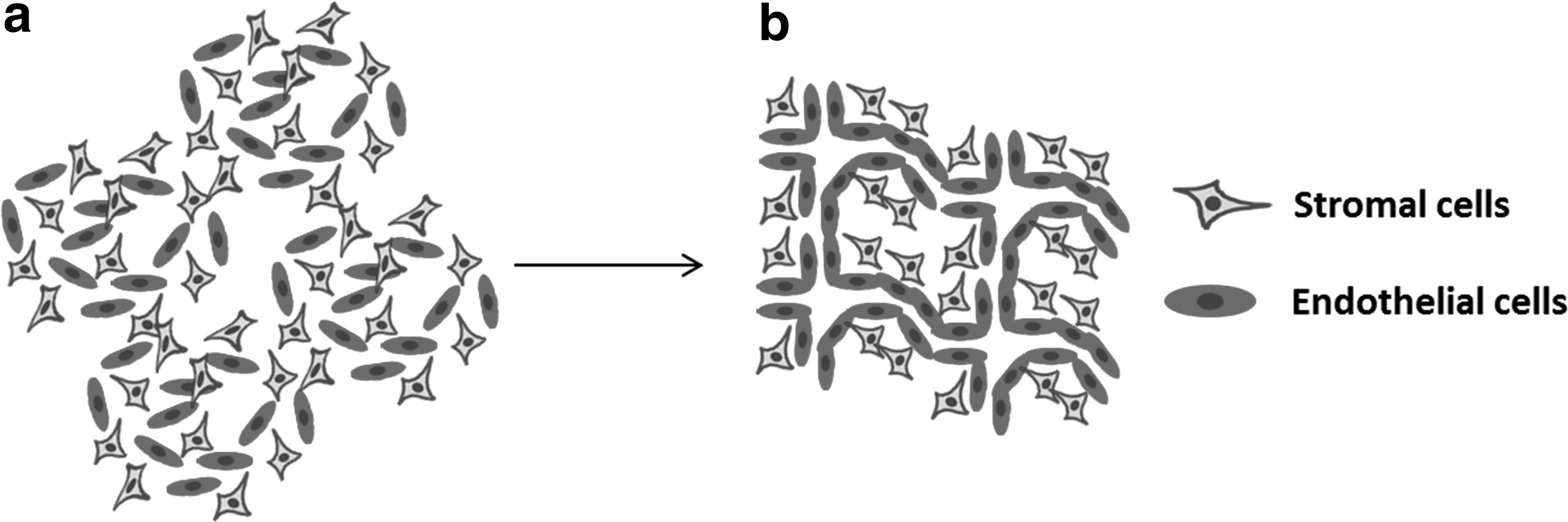
Spontaneous formation of blood vessels in vitro through vasculogenesis.
With the advent of microfluidic devices, more control over the physical and chemical environment can be exerted during tissue generation and maturation. For example, interstitial flow can easily be generated in a controlled and stable form, 80 and the creation of defined chemical gradients (e.g., VEGF gradient) is also possible. Generally, the primary limitation of such systems resides in the ability to only generate microtissues that are a few hundred microns in thickness. More recently, several groups have been able to leverage the microenvironmental control of microfluidic devices to create flow within the vascular networks.73,81–84 These perfused vascular networks represent a major advance in the creation of vascular networks, as they include an important feature of in vivo capillary networks; namely, convective transport of nutrients and waste products and endothelial shear stress.
The second major strategy to create microvessels employs 3D templating, which provides more control over multiple parameters (dimensions, shape, network distribution and branching, etc.) in the construction of the microtissue. Three-dimensional templating is the generation of a vascular template using a solid substrate within an ECM, which is eventually removed to create an empty channel. Cylindrical rods and needles have been the mainstay due to a relatively simple and straightforward process.85,86 Such systems have numerous advantages such as live cell imaging of processes, including cell intravasation and extravasation,87,88 endothelial cell structure and function, and transport of solutes.72,89–93 One limitation in using rigid rods or needles to create microvessels is the perfectly straight cylindrical shape of the vessels created by the rod, a shape which rarely occurs in vivo. An additional limitation is the gel matrix needs to be sufficiently stiff to maintain the integrity of the channel during and following the needle removal process. Collagen type I and fibrin gels (or a mixture of both) have routinely been used to construct microchannels using this method. Once the empty channel is generated, endothelial cells are seeded onto the surface and left to form a confluent monolayer over 12–48 h. There are many advantages with such systems, including the relative simplicity and reproducible geometry that allows for the control of fluid shear stress and pressure. A similar and just as powerful methodology to 3D templating is the patterning in 3D of gelatin methacrylate and other hydrogels. 94 This technique enables the creation of complex and cell-responsive microenvironments for the study of microvascular networks.
More recently more complex methods for 3D printing95,96 and templating were presented that include the creation of a 2D or 3D 89 lattice from a sacrificial carbohydrate using 3D printing. The lattice is embedded within a hydrogel (collagen, fibrin, or synthetic), and once the hydrogel is set, the carbohydrate is removed by simple dissolution. The result is an interconnected network of channels that can subsequently be seeded with endothelial cells. Such a system provides a potentially powerful platform that can be used toward the construction of thicker 3D tissues. A major limitation in these bioprinted vessel networks resides in the size of the microchannel diameter. Smaller diameter microchannels are prone to clog once endothelial cells are perfused into the microchannels.
Engineering tissues with a hierarchical vascular tree may necessitate the incorporation of both artificial and spontaneous vessel formation. Moreover, the generation of bioengineered structures by additive manufacturing of biological and biologically relevant materials, has been used to generate microvascular blood vessels and vessel networks.89,97 Bioprinting spans from printer-based to laser-based and nozzle-based methods. Nozzle-based bioprinting usually has a resolution of several hundreds of microns; they are normally used for structures that are in the millimeter size range. In general, such methods require the printing of relatively large diameter vessels (500 μm). The immediate advantage of bioprinted tissue constructs reside in its generation of complex vascular networks.15,25 Table 1 summarizes the benefits and drawbacks of the two classes of 3D in vitro microvascular systems while listing their corresponding critical features.
Challenges and Opportunities
Hierarchical networks with arterioles, capillaries and venules
A hierarchical network of blood vessels may be the key to the successful implementation of thick tissue designs in vitro, but has not been demonstrated. Mechanical forces generated by blood flow during later stages of embryonic development are believed to control the remodeling of the vessel network leading to the formation of the vascular tree. As such, each vessel type is known to play very specific roles in maintaining tissue function over time. For instance, the primary role of the capillaries is to exchange nutrients, fluid, hormones, and electrolytes between the blood carrying those substances and the interstitial space. For that reason, capillary walls are thin and permeable to a number of small molecules and water. In contrast, the major function of the muscular arterioles is to control flow.
In mimicking an arteriole-like vessel, it will be necessary to ensure that endothelial cells are supported by pericytes and other mural cell types. The cellular composition of the vessel alone may not in and of itself suffice to create physiological vessel mimics. The vessel structure and morphology is just as important. As seen in vivo, vessels are dynamic and respond to the needs of local tissues. Although numerous factors (mechanical cues and biological and chemical cues) contribute to the formation of a hierarchical network of blood vessels, mechanical factors seem to contribute the most to the tree-like structure of the vasculature illustrated by the branching and pruning of blood vessels.
Mechanical factors such as blood flow patterns in the vascular tree regulate vessel network structures and morphology. This connecting of larger vessels to smaller ones in mammalian circulatory systems obeys Murray's Law, a principle that relates the diameter of daughter branches to that of the parent branch for a lumen-based system, and is responsible for the fractal-like structures of blood vessels in vivo. This principle is centered on the fact that mammalian cardiovascular systems evolved to minimize the energy required to pump blood from the heart into all parts of the organism.98,99 To our knowledge, in vitro studies that focus on creating vascular networks have not considered validating Murray's law primarily due to challenges that still exist in the perfusion of vascular structures.
Arterioles (diameter 40–300 μm with smooth muscle cells)
The delivery of nutrients through blood is tightly controlled by the blood flow in arterioles within the vascular tree. The potential demands of a tissue in various parts of the body, such as oxygen and other nutrients, are continuously monitored by microvessels and arterioles within that specific tissue. For instance, the oxygen tension and the waste product (e.g., CO2) levels affect the caliber of the arterioles, promoting either vasodilation or vasoconstriction to control blood flow. The muscular wall in arteriolar conduits is directly responsible for the arteriole's ability to decrease or increase blood flow to specific parts of the body. This critical function highlights the importance of arterioles and smaller arteries in their ability to intrinsically control their caliber to meet the perfusion needs of the tissue. The need to tissue engineer thick tissues (larger than 1 cm3) resides in the engineering a vascular tree that resembles the vasculature in vivo. Major advances have been made in engineering larger vessels using both synthetic materials and living cells. A major gap exists in tissue engineering arterioles, and unless it is filled, engineering thick tissues will remain an elusive goal.
Pluripotent stem cells for precision or patient-specific applications
The advent of stem cell technologies will facilitate the creation of patient-specific derived tissues and vascular networks.100,101 Various protocols have been used to differentiate endothelial and smooth muscle cells from induced pluripotent stem (iPS) cells. In the case of endothelial cells, numerous differentiation protocols lead to a heterogeneous population of cells that display arterial, venous, or lymphatic markers. The different subtypes of endothelial cells are heavily influenced by the concentration of key growth factors (VEGF, Ang 1, BMP-4) used in the differentiation protocol. Similarly, the differentiation of iPS cell into smooth muscle cells can be inefficient and in many instances does not yield smooth muscle cells with the kind of contractile phenotype seen in vivo.102–104 Nonetheless, these developments have the potential to minimize host rejection and develop patient-specific drug regimens. Unlike adult cells, iPS cells can be cultured in large quantities and can be differentiated into other needed cell types (e.g., endothelial cells). While iPS-derived somatic cells generally have an immature phenotype,105,106 iPS cell-derived endothelial and stromal cells have the potential to effectively create a hierarchical network of blood vessels in vitro and thus patient-specific or personalized engineering of thick tissues for transplantation.
Footnotes
Acknowledgments
This work was supported, in part, by NIH grants UH3 TR000481 and UC4 DK104202.
Disclosure Statement
No competing financial interests exist.