
Abstract
Fetal bovine serum (FBS) is the most commonly used supplement for ex vivo expansion of human mesenchymal stem cells (hMSCs) for bone tissue engineering applications. However, from a clinical standpoint, it is important to substitute animal-derived products according to current good manufacturing practice (cGMP) guidelines. Humanized alternatives to FBS include three categories of products: human serum (HS), human platelet derivatives (HPDs)—including platelet lysate (PL) or platelet releasate (PR), produced by freeze/thawing or chemical activation of platelet concentrates, respectively, and chemically defined media (serum-free) (CDM). In this systematic literature review, the in vitro and in vivo osteogenic potential of hMSCs expanded in humanized (HS-, HPD-, or CDM-supplemented) media versus hMSCs expanded in FBS-supplemented media, was compared. In addition, PL and PR were compared in terms of their growth factor (GF)/cytokine-content and cell-culture efficacy. When using either 10–20% autologous or pooled HS, 3–10% pooled HPDs or CDM supplemented with GFs, in comparison with 10–20% FBS, a majority of studies reported similar or superior in vitro proliferation and osteogenic differentiation, and in vivo bone formation in ectopic or orthotopic rodent models. Moreover, a trend for higher GF content was observed in PL versus PR, although evidence for cell culture efficacy is limited. In summary, humanized supplements seem at least equally effective as FBS for hMSC expansion and osteogenic differentiation. Although pooled HPDs appear to be the most favorable supplement for large-scale hMSC expansion, further efforts are needed to standardize the preparation and composition of these products in compliance with cGMP standards.
Background
R
The bone tissue engineering (BTE) approach involves harvesting of osteogenic cells, commonly mesenchymal stem cells (MSCs), usually from an autologous source (bone marrow, adipose tissue, dental tissues, etc.), their in vitro expansion, and combination with an appropriate carrier scaffold for in vivo implantation. 6 More recently, the use of allogeneic or “off-the-shelf” MSC-based products has been proposed, 7 although several concerns still exist around their clinical safety. 8 Nevertheless, the “triad” of (autologous or allogeneic) osteogenic cells, osteoinductive signals (growth factors [GFs] released by cells), and osteoconductive scaffolds may replicate the properties of autologous bone, without the need for invasive harvesting. 9 The prospects of BTE are very promising as demonstrated by several preclinical (see review 10 ) and some clinical studies (see reviews6,11).
Strategies for using human MSCs (hMSCs) in regenerative therapy may involve either (1) the direct clinical use of tissue fractions containing these cells (along with other supporting cells), for example, whole or concentrated bone marrow, or adipose stromal vascular fractions (SVFs), usually performed through a “chair-side” procedure without the need for current good manufacturing practice (cGMP)-grade laboratories, or, more commonly, (2) the ex vivo expansion of hMSCs from harvested tissues, for example, bone marrow or adipose tissues, in cGMP-grade conditions before clinical application, to achieve relevant number of cells. Traditionally, ex vivo expansion of hMSCs has been performed using basal culture media plus supplements to provide GFs, proteins, and enzymes to support cell growth. 12 Fetal bovine serum (FBS) (or fetal calf serum) is most commonly used to supplement hMSC cultures, because the fetal milieu is enriched with GFs and poor in antibodies. However, for clinical use it is important to substitute animal-derived products, because hMSCs can internalize xenogeneic proteins, and thus carry the risk of infection (through viral or prion agents) and immunoreaction; it has been reported that a single injection of 100 million hMSCs expanded in 20% FBS-supplemented media is associated with ∼7–30 mg of calf serum proteins. 13 In addition, there are concerns regarding FBS sample-to-sample inconsistency, and animal welfare in terms of the “3 R's” principle (replacement, reduction, refinement).14,15
Although use of clinical-grade FBS may be permitted by regulatory health agencies in phase-1 clinical studies—>80% of proposals submitted to the FDA for MSC-based products report expansion in FBS, 16 later phase trials involving larger patient groups demand the use of culture conditions free of animal-derived products, according to cGMP guidelines.14,17,18 Such “xeno-free” or humanized alternatives to FBS broadly include three categories of products: (1) autologous or allogeneic human serum (HS), (2) pooled human platelet derivatives (HPDs), and (3) chemically defined media (serum-free) (CDM). 19 HPDs are of particular interest, given the wide range of physiological GFs and cytokines released by platelets, which can significantly enhance cell growth and function.20–23 HPDs include (1) platelet-rich plasma (PRP): a concentration of platelets in a small volume of plasma and a starting point for the other platelet products, which, depending on the method of releasing GFs, are either (2) platelet lysate (PL): produced by physical disruption of platelets in PRP through one or more freezing/thawing cycle(s) or (3) platelet releasate (PR): produced by chemical activation of platelets in PRP through coagulation, most commonly with thrombin and/or calcium compounds. 23
When using new culture conditions for hMSC expansion, it is important to characterize the cells in different stages, to rule out significant changes in their properties, for example, phenotype, proliferation, and differentiation potential, both in vitro and in vivo.24,25 Preclinical in vivo testing of new regenerative therapies in relevant animal models is an important aspect of translational research, and in most cases a requirement of regulatory health agencies, before initiating human clinical trials.26,27 In BTE research, implantation of cells or cell-based constructs in ectopic, that is, nonosseous sites (e.g., subcutaneous or intramuscular), usually constitutes a starting point for proof-of-principle or feasibility studies, whereas implantation in orthotopic sites (e.g., “critical size” defects (CSDs) in calvarial or alveolar bone) aims to simulate clinical conditions, especially in large animal models (dogs, sheep, etc.), and predict potential for therapeutic efficacy.28,29
The primary aim of this study was to systematically review the available literature to answer the focused question: do hMSCs expanded in humanized (HS-, HPD-, or CDM-supplemented) media possess superior osteogenic potential in vitro and enhance bone formation in vivo in ectopic or orthotopic animal models, compared with hMSCs expanded in FBS-supplemented media? A secondary aim of the review was to compare different HPDs in terms of their GF/cytokine content and efficacy for use in cell culture.
Materials and Methods
Study design
A review protocol was developed based on the preferred reporting items for systematic reviews and meta-analyses (PRISMA) guidelines, 30 and predetermined inclusion/exclusion criteria.
Inclusion criteria
(1) Studies reporting both in vitro and in vivo outcomes of humanized hMSC cultures.
(2) (a) Inclusion of randomized or nonrandomized controlled animal experimental studies with two or more experimental groups and (b) use of ectopic (subcutaneous or intramuscular) and/or orthotopic (bone defect) models in small or large animals.
(3) Transplantation of human-derived cells (MSCs or osteoblasts) expanded in FBS-free cultures in at least one experimental group.
(4) A control group receiving FBS-supplemented culture-expanded cells.
(5) Reporting of qualitative and/or quantitative histological data, or quantitative three-dimensional (3D) radiographic data (through computed tomography [CT] or micro-CT), regarding new bone formation (NBF).
Exclusion criteria
Absence of an FBS-supplemented culture group, that is, studies reporting in vivo comparisons of scaffolds and cells expanded in humanized media versus scaffolds alone (without cells).
Search strategy, screening, and study selection
Electronic databases of MEDLINE (through PubMed) and EMBASE were searched for relevant English language literature up to and including October 2016. “Gray” literature was searched through the Google and Google Scholar search engines. Bibliographies of the selected studies and relevant review articles were checked for cross references. A specific search strategy was developed for MEDLINE (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/teb) and adapted for other databases. Titles and abstracts of the search-identified studies were screened by two authors (S.S. and A.S.) and full texts of all eligible studies were obtained. Uncertainty in the determination of eligibility was resolved by discussion with the other authors. Two authors (S.S. and A.S.) reviewed the selected full texts independently and final inclusion was based on the aforementioned inclusion criteria. A summary of the screening process is presented in Figure 1.
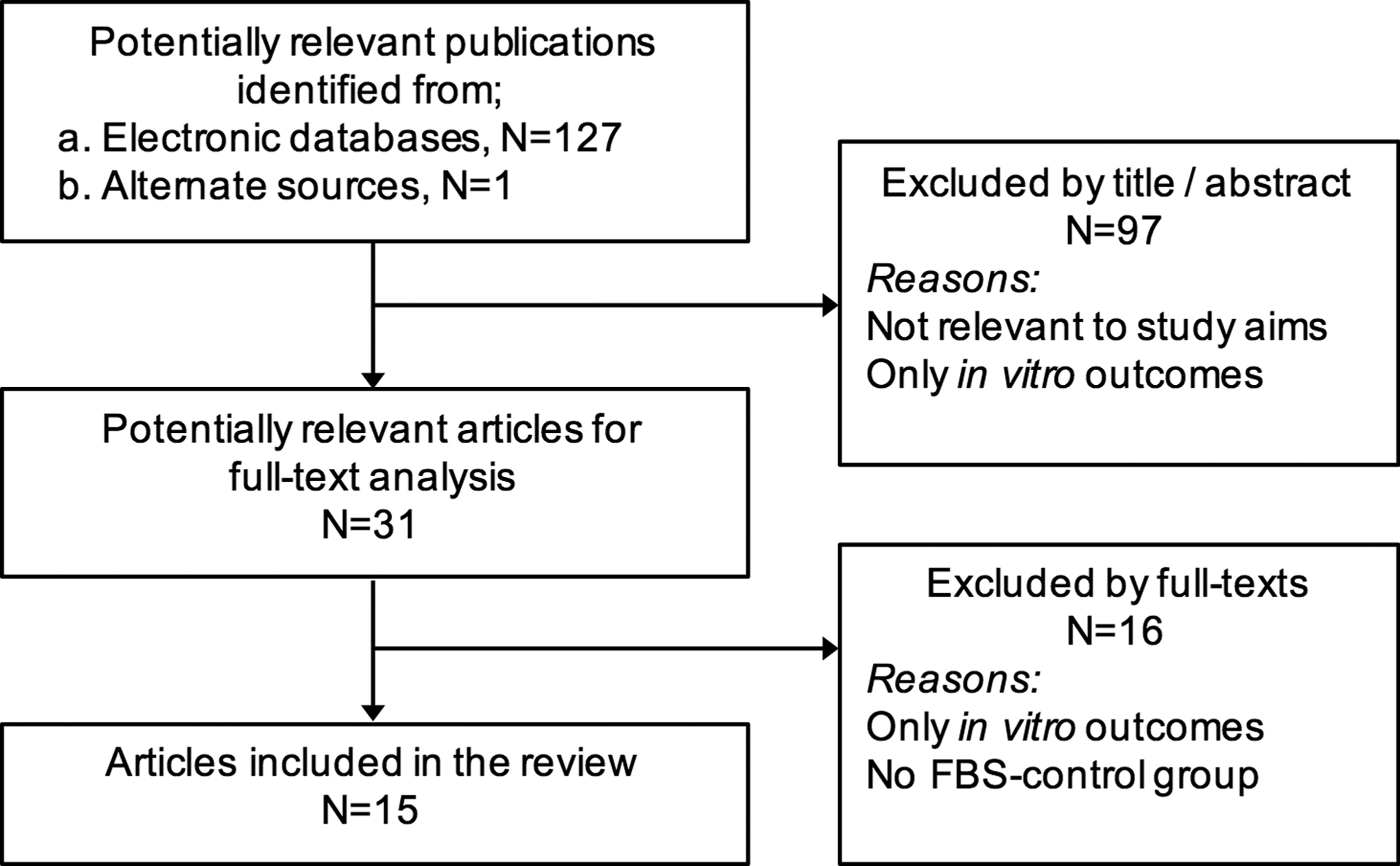
Flowchart for study screening and selection; n, number of articles.
Data extraction
Data were extracted from the full texts of selected articles on author(s), study design, in vitro evaluations (hMSC proliferation, characterization/phenotype, differentiation), in vivo evaluations (animal species, model type [ectopic/orthotopic], number of animals/procedures, observation time(s), and method(s) of outcome evaluation), main findings, and conclusions. Descriptive summaries of studies were entered into tables. Quantitative data regarding NBF were extracted for possible meta-analysis. Authors were contacted in cases of incomplete or unclear data.
Risk of bias
Risk of bias (RoB) assessment was performed using a modification of the SYstematic Review Centre for Laboratory animal Experimentation (SYRCLE) RoB tool for animal studies and judged as “high,” “low,” or “unclear”31,32 (Supplementary Table S2). Any disagreement between the reviewers during study selection, data extraction, and quality assessment was resolved by discussion and consensus.
Results
Search results and study characteristics
Of the 128 search-identified studies, 15 studies33–47 reporting both in vitro and in vivo comparisons of FBS-free and FBS-supplemented hMSC cultures were included in the review (Tables 1 and 2). A list of excluded studies along with reasons for exclusion—most commonly absence of an FBS control group—is reported in Supplementary Table S3. All included studies reported the use of rodent models, that is, rats and mice. Thirteen studies reported on subcutaneous, that is, ectopic, implantation models, three studies reported on orthotopic, that is, bone defect, models, and one study 44 reported on both ectopic and orthotopic models. Observation times ranged from 4 to 9 weeks and 5 to 12 weeks for ectopic and orthotopic models, respectively.
Risk of bias
Random allocation of animals or defects to different treatment groups, to minimize “selection bias,” was not reported in any of the studies. Similarly, no studies reported blinding of outcome assessors to treatment groups, to minimize “detection bias.” All studies reported outcome evaluation, including statistical analyses in case of quantitative outcomes, but few reported any information on adverse reactions or complications. Overall, RoB in most studies was judged to be “high” (Supplementary Table S2).
Characteristics of humanized culture strategies
Cells
All studies reported the use of human-derived cells, most commonly bone marrow-derived MSCs (BMSCs). Other types of cells used were periosteum-derived cells, 39 umbilical cord-derived MSCs (UCMSCs), 44 dental pulp-derived MSCs (DPSCs), 38 and adipose tissue-derived MSCs (ASCs) 47 or SVFs.42,45 The number of implanted cells ranged from 1 × 105 to 2.5 × 106 per implant. One study 34 reported cryopreservation (at passage 2) and revival of BMSCs after 3 months for in vivo implantation.
Media supplements
Humanized supplements could be broadly categorized as HS, HPDs (PRP, PL, or PR), and CDM. HS (5–20%) was derived from peripheral blood and used in autologous (same donor(s) as MSCs) or allogeneic forms (pooled from multiple donors). One study 40 reported the use of 3% pooled PRP with additional GFs (epidermal growth factor [EGF], platelet-derived growth factor-BB [PDGF-BB]). Four studies41–44 used 5% pooled PL, whereas one study 45 used 10% pooled thrombin-activated PR. All studies using PL or PR reported the addition of heparin to the medium (usually 2 U/mL) to prevent clot formation. Two studies used serum-free CDM: (1) a commercial MSC expansion medium (STEMPRO® MSC SFM basal medium+supplement; Invitrogen) 46 and (2) a specifically defined medium for ASC, containing growth factors (fibroblast growth factor-2 [FGF-2]/basic fibroblast growth factor [bFGF], transforming growth factor [TGF]-β, PDGF-BB), and bovine serum albumin 47 —this medium was not humanized in the true sense, as it was not completely free of animal products. In seven studies,34–36,38,41,45,46 cells were osteogenically induced before in vivo implantation.
In vitro outcomes
A summary of the in vitro outcomes in the included studies is presented in Table 1. A majority of studies reported no differences between the immunophenotype of hMSCs expanded in humanized or FBS-based cultures, based on surface antigen expression using fluorescence-activated cell sorting. Seven of the 13 studies reported significantly greater MSC proliferation in humanized cultures, that is, HS,38,39 PRP, 40 PL, 43 PR, 45 or CDM,46,47 versus FBS-supplemented cultures. Two studies33,37 reported significantly greater proliferation in FBS-supplemented versus HS-supplemented cultures. Osteogenic differentiation of hMSCs, assessed through alkaline phosphatase activity (ALP-a), gene expression of osteogenic markers, and/or mineralization assays, was reported to be comparable or greater in humanized versus FBS-based cultures. Detailed in vitro results from the included studies are presented in Supplementary Data.
Statistically significant differences.
HS-supplemented cultures had significantly higher expression of CD29+, CD49c+, CD49d+, CD106+, and/or CD71+ cells.
SVF cells were directly seeded and cultured inside a porous HA scaffold in a bioreactor system with either 10% FBS or tPR for 5 days.
PL cultures had significantly higher proportion of CD31+/CD34+ cells, whereas FBS cultures had higher proportion of CD146+ cells.
SVF cells cultured for 1 week inside fibrin gels formed CD31+ tubular structures.
CDM cultures had significantly higher proportion of CD105+ cells, whereas FBS cultures had higher proportion of CD146+ cells.
Absolute ALP-a of BMSC expanded in FBS was higher than in CDM; however, the ratio of ALP-a (ALP-a of induced cells/noninduced cells) was higher in CDM than in FBS.
Although CCL2 was higher in CDM-cultured cells than in FBS-cultured cells, CCL5 and BMP-2 were equally stimulated by TNF-α in both conditions.
3D, three dimensional; (non)induced, osteogenically (non)induced; ALP-a, alkaline phosphatase activity; ASCs, adipose tissue-derived MSCs; ARS, alizarin red staining; auto, autologous; blood, peripheral blood; bFGF, basic fibroblast growth factor; BMP, bone morphogenetic protein; BMSC, bone marrow-derived MSC; BSP, bone sialoprotein; CFU-F, colony forming unit-fibroblast efficiency; CDM, chemically defined media (serum-free); Col-I, collagen-I; cryo, cryopreserved; DPSCs, dental pulp-derived MSCs; EGF, epidermal growth factor; FBS, fetal bovine serum; FC, flow cytometry; FGF, fibroblast growth factor; FT, freeze/thaw cycles; GF, growth factor; HA, hydroxyapatite; HS, human serum; hMSC, human MSC; hT, human thrombin; IHC, immunohistochemistry; M-CSF, macrophage colony-stimulating factor; MSC, mesenchymal stem cell; n, number of cells/counts; n, number of animals; N, number of donors; ND, no significant differences; NR, not reported; OC, osteocalcin; OP, osteopontin; pass-, passage (number); PCR, polymerase chain reaction (gene expression); PD, population doubling; PDGF, platelet-derived growth factor; PL, platelet lysate; PRP, platelet-rich plasma; rh-GF, recombinant human growth factor; SVFs, stromal vascular fractions; TGF, transforming growth factor; TNF, tumor necrosis factor; tPR, thrombin-activated platelet releasate; UCMSCs, umbilical cord-derived MSCs; VEGF, vascular endothelial growth factor; VKS, von Kossa staining; yield, initial cell yield.
In vivo outcomes
Qualitative outcomes
Seven studies35–37,42–44,47 reported only qualitative histological outcomes after ectopic (six studies) or orthotopic (one study) implantation of cells grown in FBS-free or FBS-supplemented media (Table 2). In most cases, more favorable histological outcomes were reported in implants of FBS-free (HS or PL) cultured cells, for example, enhanced cellular response, better organization of collagen fibers, more osteoid and extracellular matrix formation, and superior mineralization. In one study, 44 more mature bone formation was observed in implants of BMSCs cultured in FBS media than in those of UCMSCs in PL media, whereas in another study, 42 limited bone formation was observed in implants of both FBS- and PL-cultured cells after 3D dynamic culture.
SVF cells were directly seeded and cultured inside a porous HA scaffold in a bioreactor system with either 10% FBS or tPR for 5 days. For details of cells and media supplements, see Table 2.
μ-CT, micro-computed tomography; BCP, biphasic calcium phosphate; BSA, bovine serum albumin; CDHA, calcium-deficient HA; CP, calcium phosphate; CSDs, critical size defects; HA, hydroxyapatite; histo, histological bone formation; ID, immunodeficient; n, number of animals; PLA, poly(
Quantitative outcomes
Nine studies reported quantitative (or semiquantitative) histomorphometric outcomes, most commonly, estimation of NBF within ectopic implants33,34,39–41,45,46 or calvarial defects38,44 (Table 2). One study 47 reported radiographic bone density measurements, through micro-CT, in a rat femur “fracture model.” Mineral phase (hydroxyapatite [HA], beta-tricalcium phosphate [β-TCP], biphasic [HA-TCP]) or collagen scaffolds were used as cell carriers for implantation.
In ectopic models, a majority of studies reported either similar or significantly greater NBF by cells cultured in FBS-free media than by cells cultured in FBS-supplemented media after 4–8 weeks. In one study, 33 maximum NBF was observed in implants of BMSCs cultured in FBS-supplemented media changed to serum-free CDM (supplemented with insulin–transferrin–sodium selenite), compared with BMSCs cultured continuously in FBS-supplemented media, BMSCs cultured continuously in HS-supplemented media, or BMSCs cultured in FBS-supplemented media changed to HS-supplemented media, 3 days before implantation. In two studies, adipose-derived SVF cells were directly cultured on 3D scaffolds under dynamic conditions in either 5% pooled PL 42 or 10% pooled PR 45 before implantation, and similar 42 or superior 45 NBF in comparison with cells cultured dynamically in 10% FBS (+FGF-2) was observed. However, no blood vessels of human origin could be detected in implants of PL-supplemented cells (as compared with FBS-supplemented cells), 42 but similar vessel densities were reported in implants of PR-supplemented cells and FBS-supplemented cells. 45 In another study, 44 significantly greater vessel formation was observed in implants of UCMSCs cultured in PL-supplemented media than in those of BMSCs in FBS-supplemented media (although a reverse trend for orthotopic bone formation was observed). Further analysis revealed that vessel formation resulted from the paracrine angiogenic effects of UCMSCs rather than their direct endothelial differentiation. 44
In calvarial defect models, one study 44 reported similar NBF by UCMSCs cultured in PL-supplemented media and BMSCs in FBS-supplemented media, and seeded on HA-copolymer scaffolds. However, in situ hybridization revealed that in the latter, NBF resulted from direct osteogenic differentiation of implanted BMSCs, whereas in the UCMSC implants, NBF resulted primarily through recruitment of host cells. 44 In another study, 38 significantly greater NBF and vessel formation were observed in implants of osteogenically differentiated DPSCs cultured in HS-supplemented versus FBS-supplemented media. Finally, in one study of a mouse femur fracture model, 47 similar biomechanical quality (“bending strength”) but greater radiographic bone density was observed in implants of ASCs cultured in CDM supplemented with GFs (bFGF, PDGF-BB, and TGF-β) versus ASCs in FBS-supplemented media; histology revealed a better healing response, with more osteoblastic activity, in the FBS-free ASC implants.
Discussion
A majority of the evidence for BTE is based on in vitro evaluation; however, it is important that these in vitro findings are eventually confirmed in vivo to facilitate clinical translation. 48 Recent reviews of preclinical in vivo studies have revealed favorable outcomes of BTE approaches for periodontal 32 and alveolar bone defect regeneration. 10 However, a translational limitation of traditional BTE strategies is the use of mostly animal-derived serum supplements, such as FBS, for ex vivo cell expansion. FBS substitutes are increasingly being used to develop cGMP-compliant hMSC expansion protocols according to international regulations (European Directives for EU countries, 2007/1394/EC)—to ensure reproducibility, efficacy, and safety of the therapeutic products.25,49 Human blood components (plasma, serum, platelet derivatives, etc.) represent safe and pragmatic FBS substitutes for clinical-grade hMSC expansion. 14 Moreover, commercial CDM are increasingly being proposed for hMSC expansion to minimize the risk of pathogen transfer. The type of material used for medium supplementation can largely influence cellular responses, the concentration and nature of GFs released, and, ultimately, the clinical outcomes.50–54
Summary of in vivo outcomes
The majority of reviewed studies reported ectopic models of bone formation. Ectopic models allow the evaluation of bone regeneration in nonosseous (e.g., subcutaneous or intramuscular) sites, usually in small animals. 29 The lack of endogenous osteogenic cells, cytokines, and mechanical factors (all of which can potentially stimulate bone formation) within the local intradermal environment ensures that, theoretically, any resulting bone formation after cell transplantation is of exogenous origin. 29 Overall, the majority of reviewed studies reported either similar or superior ectopic bone formation in implants of hMSCs expanded in HS- (5/6 studies), HPD- (5/6 studies), or CDM-supplemented media (1/1 study), than in those of FBS-supplemented media.
Orthotopic bone models, which involve surgically created defects in osseous sites, for example, CSDs in rodent calvaria, allow for testing of exogenous agents (e.g., cells) in the presence of endogenous osteogenic factors. 28 Rodent calvaria are considered as a challenging environment for bone regeneration because of poor blood supply and limited bone marrow and, therefore, it may be inferred that regeneration in this context is largely caused by the direct and/or indirect effects of exogenous transplanted cells. 28 All reviewed studies reported similar or superior orthotopic bone regeneration in implants of hMSCs expanded in HS-, HPD-, or CDM-supplemented media (one study each) than that in FBS-supplemented media, suggesting that hMSCs expanded in humanized media demonstrate adequate osteogenic and/or osteoinductive potential in vivo.
Characteristics of Humanized Media Supplements
Human serum
A majority of studies in this review investigated HS in comparison with FBS for hMSC expansion. In two studies,35,36 HS was further supplemented with insulin and recombinant human GFs—EGF, PDGF-BB, FGF-2, and macrophage colony-stimulating factor (M-CSF). Optimal results were observed only when HS-based cultures were supplemented, for example, a significantly lower proportion of hMSCs was observed in nonsupplemented HS-based cultures versus FBS-based cultures, but addition of supplements enhanced the hMSC yield beyond that of FBS-based cultures. 35 Regarding the concentration of HS, a majority of studies reported the use of 10% HS, whereas one study used 20% HS. 33 A previous study 55 found 10% HS to be equally or more effective than 10% FBS for large-scale expansion of human BMSCs, but 1% and 3% HS were found to be inferior. The authors did not include concentrations >10% because of significantly larger quantities of blood needed. 55 Therefore, although 10% HS appears to be effective for hMSC expansion, it is unclear whether superior hMSC growth can be achieved with higher concentrations.
Use of both autologous (i.e., hMSCs and blood derived from the same donors) and allogeneic HS (i.e., prepared from pooled whole blood of multiple donors) was reported. Autologous HS appears to be a suitable FBS substitute for clinical applications, whereas allogeneic HS is reported to be inferior to autologous HS with regard to hMSC survival and proliferation. 56 However, the amount of serum needed for large-scale MSC expansion is a potential limiting factor. Considering a 2- to 3-week expansion period (2L of medium with 10% HS), 200 mL of serum would require at least one 500 mL blood donation. 57 The lack of availability of HS from blood establishments also limits its large-scale use as an allogeneic product. 20 Furthermore, previous studies have reported lower proliferation of hMSCs in media supplemented with HS versus platelet derivatives, which have emerged as the preferred supplement for clinical-grade hMSC expansion.14,58
Chemically defined media
To circumvent the risks of pathogen transmission and difficulty of standardized preparation of HS-derived supplements, synthetic serum-free CDM have been developed. These CDM are usually supplemented with recombinant human GF combinations (PDGF, FGF, TGF-β, EGF, etc.), although the exact content and formulations of commercial media are rarely disclosed. 20 In this review, two studies reported the use of CDM for expansion of BMSCs 46 or ASCs. 47 In both studies, superior in vitro proliferation and/or in vivo osteogenesis of hMSCs were reported in CDM versus 10% FBS. However, the feasibility of using CDM for large-scale hMSC expansion has been questioned, given that (1) the inherent variability and specificity of primary cell cultures may complicate the use of CDM, (2) a specific mix of GFs will have to be developed to adjust to different hMSC sources (bone marrow, adipose tissue, cord blood, dental pulp, etc.), and (3) several recombinant GFs and their combinations required to develop certain CDM would need to be approved for therapeutic use by regulatory authorities. 20 In one study, 59 defined combinations of several recombinant GFs (TGF-β1, activin-A, bFGF, EGF, PDGF-BB, insulin-like growth factor [IGF]-1, and vascular endothelial growth factor [VEGF]) and chemokines (CCL21, CCL25, CXCL12, and RANTES) in a serum-free medium failed to achieve hMSC proliferation compared with 5% or 10% PL. Conversely, a recent study 60 reported significantly shorter expansion times to achieve clinically relevant number of hMSCs, when using a commercial CDM versus 10% PL. However, the PL-based system was found to be significantly more cost-effective than CDM (total cost of CDM was 200% that of PL) for large-scale MSC expansion. 60
Platelet derivatives
Platelet concentrates
Platelet derivatives are attractive supplements for cell culture, because platelets contain high concentrations of physiological GFs. These are usually prepared from platelet concentrates, obtained as apheresis products or whole blood-derived buffy coat units. 21 Platelet derivatives may be used as autologous or “pooled” products. Autologous products eliminate the risk of disease transmission, although obtaining a sufficient quantity, and quality in terms of GF contents, of autologous concentrates for clinical-grade hMSC expansion may be highly dependent on the donor and method of platelet isolation. 61 Alternatively, pooling platelets from multiple donors can provide larger volumes of concentrates, and also reduce donor-based variations in terms of platelet counts, GF contents, and effects on hMSCs.62–67 Studies have reported pooling of platelet concentrates from 4 up to 40 donors (Supplementary Table S4). Pooled platelet concentrates are routinely prepared by blood establishments for transfusion, 21 whereas safety practices such as donor screening and pathogen inactivation reduce the risk of transmitting infectious diseases through pooled products. 20 Together, these efforts could allow the large-scale, cost-effective, and standardized manufacturing of “off-the-shelf” humanized cell culture supplements.
Pooling can also help to enhance platelet concentrations in these products. A platelet concentration of 1 × 106/μL is considered to be of therapeutic value. 68 Allogeneic platelet concentrates with a minimum platelet content of 2 × 1011/unit in ∼300 mL are routinely prepared for platelet transfusion by blood banks in Europe. 69 A significant advantage is the ability to use “expired” platelet concentrates (older than 5 days) that are no longer suitable for transfusion, but are equally effective as fresh platelets, in supporting hMSC growth and osteogenic differentiation.17,70 However, platelet concentrations cannot solely predict GF levels, 71 because the method of preparation, specifically the method of platelet “activation,” can largely affect the content and efficacy of the final product, that is, the lysate (PL) or releasate (PR). 72
Platelet lysate and platelet releasate
PL is usually prepared from pooled PRP units by one or more freezing and thawing cycle(s) to mechanically disrupt the platelet membranes, whereas PR is prepared by chemical activation of platelets in PRP, most commonly by addition of thrombin and/or calcium chloride.45,73 Although PL contains the entire intracellular contents released from platelets, activation with thrombin or calcium in PR closely mimics physiological platelet activation and GF release that occurs during wound healing. 74 Moreover, activation with exogenous thrombin causes a rapid release of platelet GFs (a majority of the stored GFs are released within the first 1–4 h75,76), whereas activation with calcium causes a more gradual GF release (for 7 days) through the formation of endogenous thrombin and “partial” platelet activation.77,78 Recent studies have identified optimal GF release profiles after PRP activation with solely calcium, 79 or calcium with a low dose of thrombin. 78
Growth factors
Platelets contain large quantities of GFs, such as platelet-derived growth factor isoforms (PDGF-AA, -AB, and -BB), TGF-β, EGF, FGF-2, VEGF, brain-derived neurotrophic factor, hepatocyte growth factor, connective tissue growth factor, and bone morphogenetic protein (BMP)-2, -4, and -6. 21 Of these, PDGF-BB and FGF-2 have been identified to be particularly important for the hMSC growth-promoting effects of platelet derivatives. 80 However, it is challenging to compare platelet derivatives (or other humanized supplements) with FBS in terms of their GF contents, given the differences in their origins, and also because of the scarcity of literature regarding the GF contents of FBS. To our knowledge, only one published study has compared GF contents in PL and FBS and reported significantly higher concentrations of PDGF-AA, -AB, -BB, TGF-β, IGF, and VEGF in PL. 81
Recent studies82,83 have also highlighted the role of platelet-derived extracellular vesicles (EVs), also known as microparticles or exosomes, which may be internalized by hMSCs, thereby functioning as an efficient mechanism for GF delivery. These EVs, present in PL and PR, have been shown to contain a high concentration of GFs, for example, FGF, VEGF, PDGF-BB, and TGF-β1, and are thus mediators of platelet-stimulated hMSC proliferation and osteogenic differentiation.83,84
A number of studies have compared GF contents in PL and PR, and between PL/PR and HS (Supplementary Table S4). The most commonly evaluated GFs were PDGF-AA, -AB, -BB, TGF-β1, FGF, EGF, IGF-1, and VEGF; these GFs are commonly associated with favorable hMSC proliferation and osteogenic differentiation.85,86 However, because of large differences between studies in terms of protocols for preparation of the products, and methods for evaluating GF concentrations, specific values of GF concentrations could not be reliably compared. Nevertheless, these studies demonstrated that both PR and PL contained high concentrations of multiple GFs and equally promoted proliferation and differentiation of various cell types, including hMSCs. Most commonly, high concentrations of PDGF isoforms, TGF-β1 and FGF, were identified in PL and PR, all of which are considered important for hMSC recruitment, proliferation, and osteogenic differentiation.87–90 Other bone-related proteins such as osteocalcin and osteopontin have also been identified in PL. 91 Interestingly, no studies compared the release of BMPs from platelet products. BMPs are members of the TGF-β superfamily, and important regulators of skeletal development and bone formation. 92 BMP-2, -4, -6, -7, and -9, which are reported to be the most potent inducers of osteogenic differentiation,93,94 have been identified in PL.95,96 Moreover, platelets may contain potentiators of BMP-mediated osteogenic differentiation. 97 However, further research is needed to determine whether the presence of these GFs in culture media indeed “primes” hMSC differentiation toward an osteoblastic lineage. 86
It is currently inconclusive whether PL is superior to PR, or vice versa, because of differences in the methods of PL/PR preparation and platelet concentrations, which do not allow direct interstudy comparisons. However, a trend for reporting of higher GF concentrations in PL was observed, and the number of freeze/thaw cycles and/or freezing temperatures seemed to affect GF release in PL—2–5 cycles were reported to be adequate for achieving optimal platelet lysis56,98–100 and lower freezing temperatures were beneficial when using fewer cycles. 101
Inflammatory cytokines
In addition to GFs, platelet products contain physiological levels of a range of inflammatory cytokines, including interleukin (IL)-1α, IL-6, IL-7, IL-8, tumor necrosis factor (TNF)-α, granulocyte-colony stimulating factor, granulocyte macrophage colony-stimulating factor, macrophage inflammatory protein (MIP)-1α, MIP-1β, and interferon-γ, among others. 20 These cytokines play important roles in chemotaxis, cell proliferation, differentiation, and angiogenesis. Evidence for the effects of inflammatory cytokines on MSC osteogenic differentiation is conflicting, with several cytokines reported to have both pro- and antiosteogenic effects (see review 102 ). However, recent studies have suggested that controlled delivery of certain inflammatory cytokines may enhance hMSC proliferation and osteogenic differentiation.103–105 Indeed, these findings can be correlated with in vivo observations of osseous wound healing, which is characterized by an initial “inflammatory phase” during which several proinflammatory cytokines are upregulated to recruit and guide the proliferation and differentiation of MSCs at the injury site and initiate the regenerative process. 106
Differential expression of bioactive factors and consequent effects on hMSC proliferation have been reported in PR and PL. 107 In a detailed comparison of 174 cytokines (and GFs), Bieback et al. 58 reported similar expression of a majority of cytokines in PL and thrombin-activated platelet releasate (tPR), including IL-1α, IL-1β, and TNF-α, although higher concentrations of PDGF-AA, -BB, -AB, and VEGF were reported in PL. Conversely, another study 108 reported significantly higher levels of IL-1α, IL-1β, IL-6, monocyte chemoattractant protein (MCP)-1, INF-γ, TNF-α, and M-CSF in PL versus PR. Interestingly, in one study, 109 significantly higher levels of IL-1β were detected in tPR 1–6 days after thrombin activation (mean 425 pg/mL) versus PL (i.e., total intracellular IL-1β content, mean 1.4 pg/mL), suggesting de novo synthesis. 110 This finding was supported by a more recent study, 78 which reported a 10-fold increase in IL-1β levels in PRP (vs. baseline) after calcium or thrombin activation. Similarly, another recent study 76 identified a number of proinflammatory cytokines, most prominently chemokine CCL5/RANTES, to be upregulated in tPR for 5 days, which is in agreement with a previous report. 58 In context, results from a number of studies suggest a possible negative effect of thrombin activation on PRP-mediated in vivo bone regeneration.111–114 Indeed, high levels of RANTES have also been identified in PL preparations.58,80,115 Interestingly, in one of these studies, 80 inhibition of RANTES (or other cytokines, e.g., PDGF-AA and VEGF) in 10% PL-supplemented medium had no effect on hMSC proliferation. Nevertheless, when considering platelet products for clinical-grade MSC expansion, use of exogenous products for platelet activation, such as thrombin (especially of nonhuman origin 116 ), may complicate the regulatory approval process. 21
In vitro and in vivo osteogenic efficacy
In this review, studies reported the use of either 5% PL or 10% tPR. A lower concentration of PL (5–10%) than FBS (usually 10–20%) may be adequate for cell culture because of a higher concentration of GFs in PL 81 ; an optimal media composition of alpha-minimal essential medium supplemented with 10% PL has been reported for hMSC expansion. 54 This is consistent with current literature, which suggests that PL in concentrations of 5–10% can enhance the growth of several cell types compared with 10% FBS,20,21,98,117,118 and effectively support the clinical-grade expansion of hMSCs from different sources in vitro.119,120 The efficacy of 5–8% PL-expanded hMSCs to regenerate bone in calvarial CSD of nude mice has previously been reported. 121 This expansion protocol is currently being applied in an ongoing clinical trial of alveolar bone regeneration with autologous BMSCs. 122
Vascularization of bone constructs is an important determinant of their in vivo success, and is considered to be a translational limitation of current BTE strategies.123,124 The proangiogenic effects of platelet products have been well studied, both in vitro and in vivo, 125 and it could be hypothesized that culturing hMSCs in platelet-supplemented medium could enhance their proangiogenic properties through paracrine mechanisms. This was confirmed in one included study, 44 wherein a significantly higher number of blood vessels were detected in ectopic implants of UCMSCs cultured in 5% PL versus BMSCs in 10% FBS; the vessels were determined to be of host origin, confirming that the observed angiogenesis was a result of the paracrine effects of implanted UCMSCs, rather than direct endothelial differentiation. 44 In contrast, two studies, in which adipose-derived SVF cells containing distinct subpopulations of endothelial cells (CD31+/CD34+) were cultured under dynamic conditions in either 5% pooled PL 42 or 10% pooled tPR, 45 in comparison with 10% FBS (+FGF-2), identified in vivo blood vessels of human origin in implants of tPR- and FBS-cultured cells, suggesting direct endothelial differentiation, but not in PL-cultured cells.42,45 Possible explanations for these observations could be differences in PL, PR, and FBS, in terms of (1) GF profiles (as previously discussed), (2) their ability to support the growth and differentiation of endothelial cells, and (c) their effects on hMSC secretory profiles. Although both PL 126 and PR 127 have been shown to independently stimulate angiogenesis in vitro, their effects on hMSCs, particularly on their in vitro and in vivo angiogenic and osteogenic secretory/paracrine profiles, are currently unclear.
In the context of (bone) tissue engineering, implantation of exogenous hMSCs is expected to induce regeneration through direct (osteogenic) differentiation and/or through paracrine mechanisms, that is, secretory molecules stimulating endogenous cells (see reviews128,129). Paracrine effects of hMSCs can also have anti-inflammatory and immunomodulatory functions, that is, suppression of the host immune response. Results from previous in vitro studies suggest that secretory profiles of hMSCs may vary depending on culture conditions, in the contexts of both regeneration, for example, between PL- and PR-supplemented medium,58,107 and immunomodulation, for example, between PL- and FBS-supplemented medium.130,131 However, whether humanized cultures enhance the paracrine and/or immunomodulatory functions of hMSCs in vivo remains to be determined.
Long-term expanded hMSCs undergo reduction in their proliferation and differentiation potential as a result of “aging” or replicative senescence,132,133 which may be influenced by culture conditions. 134 However, studies that have investigated the influence of humanized media reported significantly higher proliferation (population doublings), but similar senescence-associated changes, in hMSCs cultured in either HS- 135 or PL-supplemented media, 136 compared with FBS-supplemented media, with no evidence of chromosomal transformations, suggesting that senescence-associated changes in hMSCs may be independent of culture supplements. Interestingly, changing to PL-supplemented media has been reported to attenuate senescence in late-passage FBS-expanded hMSCs and induce “cellular rejuvenation” by enhancing, or at least maintaining, their proliferation and differentiation potential. 137 These findings further support the substitution of FBS in hMSC cultures with humanized supplements, particularly PL.
Few published reports exist regarding the clinical applications of PL-expanded MSCs for bone regeneration. In contrast, PRP has been extensively used for this application, alone, in combination with bone substitute materials, 138 or as a scaffold for hMSCs, 139 although the evidence regarding its clinical efficacy is inconclusive. 140 The use of autologous BMSCs expanded in 10–20% autologous PL has been reported for various orthopedic indications in a large patient sample (n = 339) 141 —BMSCs were mixed with autologous PL or PRP and injected into peripheral joints or intervertebral disks. No evidence of tumor formation at the injection sites or major adverse events related to the procedure were identified after up to 3 years. 141 Finally, one ongoing clinical study 122 is investigating the use of autologous BMSCs expanded in 5–8% pooled PL-supplemented media, in combination with biphasic calcium phosphate scaffolds, for alveolar bone regeneration.
Although a majority of clinical trials of hMSC-based therapy report the use of autologous cells—most often expanded in FBS media—the use of allogeneic hMSCs is emerging as a promising alternative. 18 Allogeneic MSCs, often “pooled” from multiple donors, have been applied in the treatment of graft-versus-host disease (GVHD), 142 osteogenesis imperfecta, 143 and, more recently, in osteoarthritis. 144 To our knowledge, no published clinical trials have reported the use of allogeneic hMSCs for bone regeneration. Nevertheless, recent trials145–147 have demonstrated the effectiveness of pooled allogeneic hMSCs (8–12 donors) expanded in PL-supplemented (5–10%) media for treatment of GVHD, in both adults and children. It has previously been suggested that variability of observed clinical success rates may be attributed to donor-based variations in hMSCs, and “xenocontamination” through FBS cultures, 148 which may lead to hyperimmunogenicity, improper trafficking, and poor engraftment of implanted hMSCs. 149 Variations in hMSC properties, including osteogenic differentiation potential, between donors 150 and tissue sources, 120 even when cultured in PL-supplemented media,120,151 have been reported. An interesting avenue for future research could be to determine whether optimization of humanized supplement production can attenuate donor-based variations in allogeneic hMSC cultures.
Methodological issues and quality of reporting
In this review, no meta-analysis of data regarding in vivo bone regeneration from individual studies could be performed, because of large inconsistencies (heterogeneity) in the nature of the animal models used, therapies tested, and outcomes evaluated. Furthermore, the review identified a lack of studies comparing humanized alternatives and FBS as hMSC culture supplements for in vivo bone regeneration. Therefore, an overall “pooled estimate” of the efficacy of FBS-based and FBS-free hMSC cultures could not be obtained. 152 Since the reliability of results of systematic reviews directly depends on the quality of the primary studies, RoB within the included studies was assessed through compliance with the SYRCLE tool, which addresses particular aspects of bias that play a role in animal experimental studies. 31 Studies were judged to be of a high RoB, and a clear need for more standardized reporting of studies was identified, to allow reliable future reproduction and synthesis.
Conclusions
Based on the results of 15 studies in rodents, mostly using ectopic (subcutaneous) models, implantation of hMSCs cultured in humanized media results in similar, if not superior, in vivo bone formation compared with that of hMSCs cultured in FBS-supplemented media. In a majority of studies, the in vitro benefits of humanized cultures were translated in vivo. Pooled platelet derivatives currently represent the most feasible alternative to FBS for clinical-grade hMSC expansion. Based on limited evidence, a trend for reporting higher concentrations of GFs was observed in PL versus thrombin- and/or calcium-activated PR. However, further well-designed studies are needed to validate the preclinical and clinical in vivo efficacy of platelet derivatives, in addition to their cost-effectiveness, in comparison with that of other FBS substitutes. Moreover, standardization of preparation protocols, in compliance with cGMP standards, is needed to facilitate their large scale use for hMSC expansion.
Footnotes
Acknowledgments
This review was supported by funding from Helse Vest, Norway (project number: 912048, K.M.), and the Bergen Stem Cell Consortium (project number: 502027, K.M.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.