
Abstract
Multinucleated giant cells (MNGCs) are a special class of giant cell formed by the fusion of monocytes/macrophages abundantly found in human tissues. While historically their role around certain classes of biomaterials have been directly linked to a foreign body reaction leading to material rejection, recent accumulating evidence has put into question their role around certain classes of bone biomaterials. It was once thought that specifically in bone tissues, all giant cells were considered osteoclasts characterized by their ability to resorb and replace bone grafts with newly formed native bone. More recently, however, a special subclass of bone biomaterials has been found bordered by large MNGCs virtually incapable of resorbing bone substitutes even years after their implantation yet surrounded by stable bone. Interestingly, research from the field of cardiovascular disease has further shown how a shift in macrophage polarization from M1 “tissue-inflammatory” macrophages toward M2 “wound-healing” macrophages in atherosclerotic plaque may lead to MNGC formation and ectopic calcification of arteries. Despite the growing observation that MNGC formation occurs around certain bone biomaterials, their role in these tissues remains extremely poorly understood and characterized. In summary, four central aspects of this review are discussed with a focus on (1) the role of MNGCs in bone/tissue biology, and their ability to induce vascularization/new bone formation, their role around, (2) bone substitutes for bone augmentation, (3) dental implants, as well as (4) during peri-implant infection. The authors express the necessity to no longer refer to MNGCs as “good” or “bad” cells, but instead point toward the necessity to more specifically characterize them scientifically and appropriately as M1-MNGC and M2-MNGC accordingly. Future research investigating the factors influencing their polarization as a “center of control” is also likely to act as a key factor in the progression/resolution of various diseases.
Introduction
M
Monocytes and macrophages are some of the most abundant cell types found in the human body and act as key implicators in bone biomaterial integration since they represent the first cell types in contact with these materials. 5 It is noteworthy that macrophages are one of the most broad ranging cell types capable of polarizing entirely from contributors of tissue inflammation (M1 macrophage) toward contributors of wound healing (M2 macrophage). 6 Due to their broad role in regulating tissue homeostasis, they have extensively been studied in innate and adaptive immunity, as modulators of wound healing, hematopoiesis, as well as malignancy. 6
Despite their widespread role in regulating tissue homeostasis and integration, a recent systematic review found that in the field of bone biomaterial integration, less than 10% of all studies focused on the osteoimmunological reaction of cells toward biomaterials with over 90% of research focused primarily on osteoblast/fibroblast behavior to implanted bone biomaterials. 7 This discrepancy is difficult to understand given the fact that monocytes and macrophages are the first cells to come in contact with implanted biomaterials and the major modulators of tissue biomaterial integration. 8
Interestingly, certain classes of bone biomaterials such as high-temperature sintered xenografts and certain zirconia dental implants are known inducers of MNGC formation, yet these biomaterials fully integrate readily in vivo.4,9 In certain clinical situations, such bone substitute materials are favorably utilized due to their “low-substitution” rate meaning that bone volume can be maintained for long periods. Clinicians often choose to apply such materials in areas such as the labial surface in the maxillary esthetic zone following dental implant placement whereby such biomaterials can “maintain space” due to low bone turnover and limited material resorption. While we have been continuously intrigued by the role of MNGCs in bone biomaterial integration, recent studies investigating atherosclerotic plaque from the past 3–5 years have provided evidence that immune cells (macrophages and MNGCs) are the responsible cell type controlling ectopic calcification in arterial walls.10–14 It therefore is of relevance that while MNGCs around atherosclerotic plaque are considered pathological due to their role in arterial calcification (a life-threatening situation), a similar MNGC around a bone biomaterial may actually be considered a natural turnover mechanism if performing a similar role, thereby creating high interest in the field of bone biomaterial research.
Therefore, the purpose of this review article is to further summarize knowledge acquired to date on MNGC with the following specific aims: (1) to present and summarize MNGC biology in bone tissues, including their fusion from their precursor cells, the osteal macrophage (OsteoMacs). (2) We then describe and compare macrophage polarization and MNGC formation around ectopic calcification of arteries. (3) Finally, this review article discusses the role of MNGCs around (A) bone grafts, (B) dental implants, and more specifically (C) during peri-implant infection/regeneration as a potential shift between M1-MNGC and M2-MNGC polarization. In summary, the authors express the requisite to no longer refer to MNGCs as “good” or “bad” cells, but instead point toward the necessity to better characterize these giant cells scientifically and appropriately as M1-MNGC and M2-MNGC accordingly.
MNGC biology in bone tissues, including their fusion from precursor cells
Over the years, basic science research has revealed the dynamic interactions between the skeletal and immune systems thereby rendering the creation of an entire field termed “osteoimmunology” to further study bone-immune interactions.15–17 With respect to bone tissues, precursor cells of MNGCs are thought to be derived from osteal macrophages (“OsteoMacs”), a distinct population of macrophages residing within bone actively participating in bone modeling and remodeling throughout the life span of mature osteoblasts. 17 Initial macrophage experiments identified their ability to polarize toward two specific phenotypes, classical M1 pro-inflammatory macrophages and M2 wound healing macrophages. Classical M1 macrophages are induced in response to lipopolysaccharides expressing pro-inflammatory cytokines such as tumor necrosis factor-alpha,18,19 interleukin (IL)-6,20,21 and IL-1β,18,22 all contributing to tissue inflammation and osteoclastogenesis. M2 macrophages are induced by IL-4 and IL-13 and typically produce transforming growth factor-β and arginase, both factors implicated in the tissue repair process.23–26 Although macrophages were initially and more commonly known as being implicated in the inflammatory process, a series of mechanistic experiments have also revealed their essential roles in bone development, formation, and repair by releasing M2-related cytokines and growth factors.27–31 Nevertheless, the differentiation and polarization of monocytes toward M1 or M2 macrophages, as well as their fusion to osteoclasts or MNGCs in response to external stimuli and/or bone biomaterials, remains to this day extremely poorly understood.
Previously we reviewed how the monocyte/macrophage lineage polarize and differentiate toward MNGCs and osteoclasts (Fig. 1). Due to common precursors, OsteoMacs have been the favored precursor cell of MNGCs. 5 Interestingly, differences in surface markers and gene expression between osteoclasts and MNGCs have been reviewed recently. 32 To date, the transcriptional regulation responsible for the formation of osteoclasts versus MNGCs from precursor cells in bone tissues is poorly characterized with much greater investigation needed. Emerging findings from our laboratories, as well as others have shown that MNGCs are also capable of polarizing much like macrophages toward M1 and M2 phenotypes, 27 thereby opening the possibility that they act as key regulators during biomaterial integration and peri-implant infection/resolution.
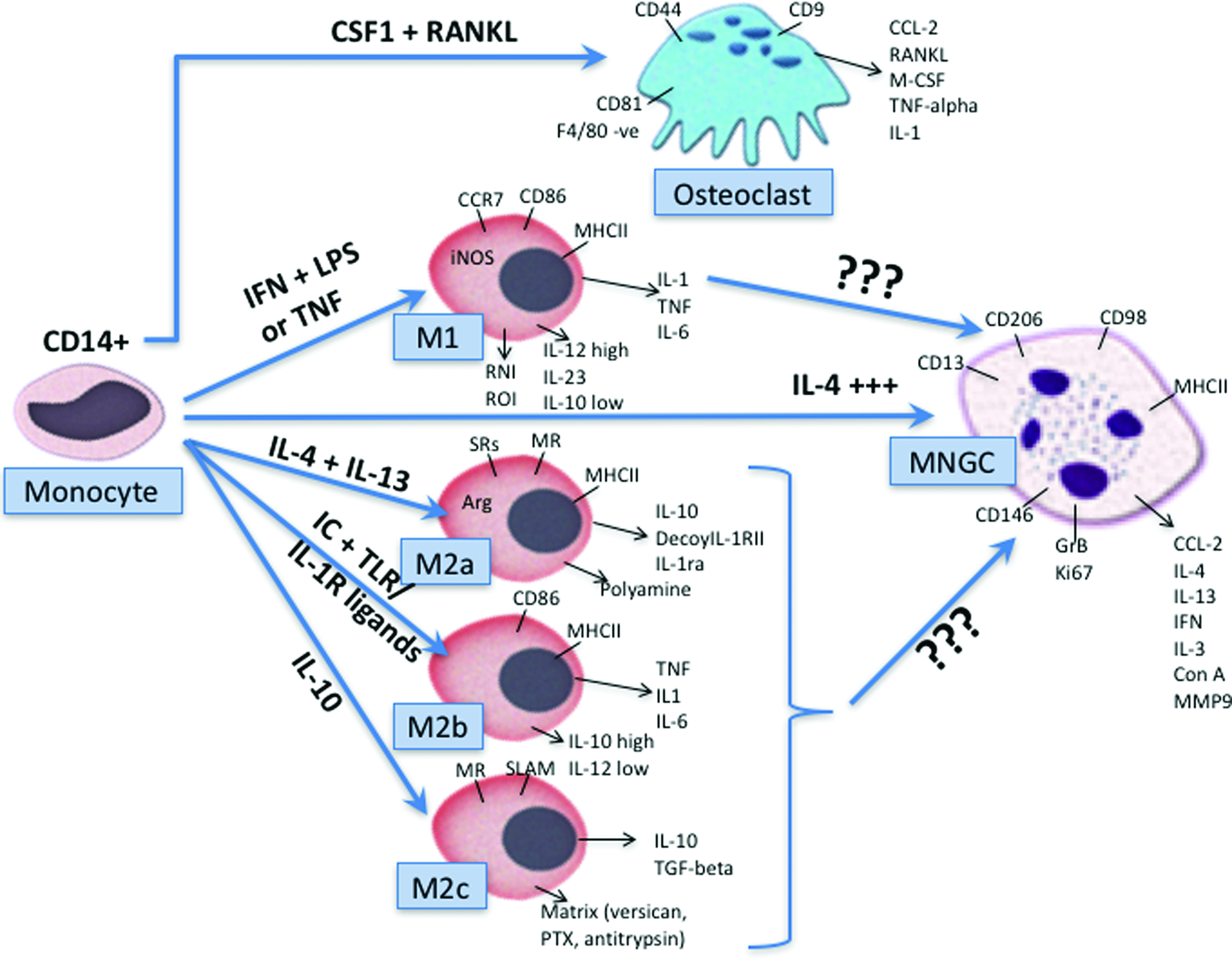
Monocyte differentiation, including expression markers into osteoclasts, M1, M2a, M2b, M2c macrophages, and MNGCs. Reprinted with permission from Miron and Bosshardt. 5 MNGC, multinucleated giant cell. Color images available online at www.liebertpub.com/teb
Giant-cell formation and function
While it is known that MNGC formation is the result of cells derived from the monocyte/macrophage lineage, over the years a variety of different names have been given based on their perceived roles in tissues, including foreign body cell, foreign body giant cell (FBGC), multinucleated cell, MNGC, giant-body foreign cell, or “foam” cell. Despite this, it is important to note that these cells are phenotypically derived from the same precursor cells and often confused in terminology. Early in vitro experiments dealt primarily with demonstrating their role in response to pathogens, an obvious inflammatory state. In those studies, large MNGCs with 15 nuclei or more were routinely found in response to high-virulence mycobacterium thereby inducing a “foreign body” terminology for MNGC described as FBGCs. 33 Simply put, these “unhappy” macrophages were shown to routinely fuse in response to foreign pathogens and thus given the working name FBGCs.
More recently, much advancement has been made with respect to MNGCs. Many biological aspects of their behavior have been discovered from biomaterials implanted in soft tissues, including their recognition motifs, adhesion molecules, fusion pathways, as well as specific intercellular and intracellular signaling pathways (Fig. 2).34,35 Yet with respect to bone biomaterials, a great deal of information is still lacking regarding their cellular control, and since their binding motifs are often presented in response to hydroxyapatite-derived surfaces onto bone biomaterial surfaces, great differences are expected in comparison to soft tissue biomaterials, yet this field of research remains entirely unstudied.
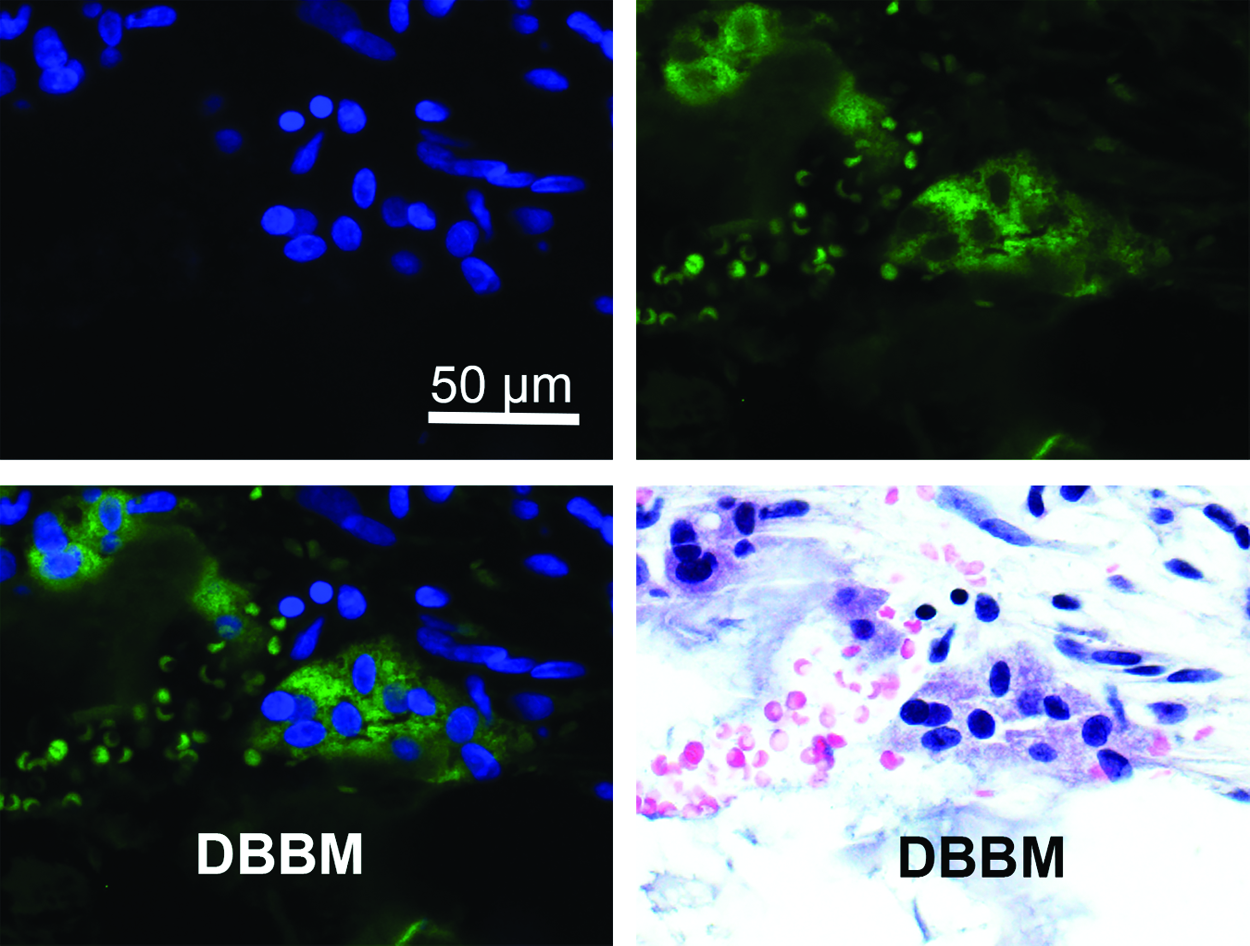
MNGC on the surface of a bone grafting material deproteinized bovine bone mineral (DBBM) stained for DAPI (top left), HLA-DR (top right), combined (bottom left), and H&E (bottom right). HLA-DR is a marker only expressed in MNGCs and nonexpressed in osteoclasts confirming the presence of MNGC-specific cells around bone biomaterials. H&E, hematoxylin and eosin; HLA-DR, human leukocyte antigen-D related. Color images available online at www.liebertpub.com/teb
It is also interesting to note that MNGCs in response to biomaterials form in response to foreign particles greater in size than permitted for macrophage phagocytosis to occur. 36 In response, “frustrated” macrophages fuse into larger MNGCs in an attempt to thereafter phagocytize larger foreign particles. 36 In the context of soft-tissue biomaterial integration, it has been proposed that “FBGCs are generated by macrophage fusion and serve the same purpose as osteoclasts and degradation/resorption/removal of the underlying substrate”. 37 While this may be the case in soft tissues and the authors do not oppose this view, it is interesting to point out that MNGCs are also found around bone biomaterials, characteristically different than osteoclasts by their inability to resorb and replace bone grafts and dental implants. Therefore, and as previously stated, “If these cells serve the same purpose as osteoclasts in bone, one has to ask the question why are these cells found in bone altogether?” 5
Macrophage Polarization and MNGCs in Atherosclerosis
A major pathway for the development of calcifying arteries in atherosclerosis is the critical role and involvement of immune cells/macrophages during ectopic bone formation. While it is known that initially, classically activated M1 macrophages are present within these tissues expressing high levels of interferon-gamma,38,39 it was recently discovered that they later polarize toward M2 wound healing macrophages thereby causing ectopic bone calcification within arterial walls. 40 Within the inner most intimal layer of the artery, monocytes have been shown to accumulate and differentiate into macrophages where they ingest modified lipoproteins through scavenger receptors and secrete inflammatory mediators giving rise to lipid-rich macrophages (foam cells) responsible for the lipid core buildup. For yet an unknown mechanism, a transient shift of polarization of these cells has been observed from M1 to M2 macrophages subsequently causing ectopic bone formation and calcification of arteries. 41
Interestingly, it has been demonstrated that the formation of M2 macrophages was primarily activated by endoplasmic reticulum stress, an ongoing area of research in the cardiovascular field. 40 Current research in this field has proposed actually suppressing M2 macrophage phenotypes and attempt to modify/repolarize macrophages back to pro-inflammatory M1 macrophages to prevent ectopic mineralization/calcification of arterial walls, a scenario highly dangerous and life-threatening. 40 Presently, it remains poorly understood why these macrophages shift from M1 to M2 phenotypes in these tissues. Nevertheless, it remains interesting to note how in one clinical scenario, M2-macrophage accumulation and MNGC “foam cells” in calcified arteries producing ectopic bone are considered pathological and potentially life threatening, whereas that same cell type located around certain classes of bone biomaterials would ultimately be considered greatly therapeutic, thus necessitating the need for future research in these fields.
MNGCs around bone biomaterials
It was once thought that all MNGCs surrounding bone biomaterials were osteoclasts due to their similarities in histological features between both cell types. More recent research derived from soft tissue biomaterial integration has however revealed marked differences between MNGC formation and osteoclasts.42,43 One of the main difficulties encountered to date in the bone biology field has been to characterize giant cells from bone biopsies. MNGCs assessed around dental implant surfaces are routinely embedded in resins (as opposed to paraffin due to the presence of metal or ceramic), thereby preventing the ability to use routine antibody staining and immunohistochemistry or fluorescent techniques commonly utilized in paraffin sections.
Interestingly, initial observations noted that MNGCs located around bone grafts appeared to be distinctly different from osteoclasts. 44 In this study, it was found that certain synthetic bone particles appeared to recruit MNGCs similar to osteoclasts by their TRAP activity, ruffled borders with calcitonin receptor expression; however, an obvious inability to resorb bone particles was observed. 44 What was the role of these cells in bone tissues?
Some years later, Kelly and Schneider implanted mineralized and demineralized bone, as well as composite (nonmineralized) materials into the dorsal body wall of young adult rats, and found that only a small population of MNGCs was observed on the composite implant, whereas MNGCs on demineralized implants demonstrated high TRAP expression similar to osteoclasts. 45 Around the same time, Marks and Chambers observed giant cells to mineralized bone particles in subcutaneous tissues that did not share similar functional features and enzymatic activity as osteoclasts. 46 Interestingly, in 1995 Dersot et al. renamed these giant cells “macrophage polykaryons” after confirming certain differences between the MNGCs recruited to a synthetic material fabricated of hydroxyapatite revealing that “they did not exhibit the morphologic, enzymatic, and functional characteristics of osteoclasts.” 47
Interestingly, a recent review article by our group characterized cell surface marker differences between osteoclasts and MNGCs. Out of 19 potential markers which have been previously utilized to identify osteoclasts from MNGCs, only 5 markers were found to represent ideal candidates whereby expressed in only one of the two cell types. These included calcitonin receptor and RANK in osteoclasts and CD86 (B7-2), CD206, and human leukocyte antigen-D related (HLA-DR) in MNGCs. 32 Of the remaining, six were found more highly expressed in one cell type versus the other, including TRAP, cathepsin K, and MMP9 in osteoclasts and CD68, CD98, and B7-H1 (PD-L1) in MNGCs. The remaining eight markers, including CD9, CD13, CD14, CD40, CD44, CD51, CD147, and EMR1 (F4/80), showed no preference for either cell type. 32 While these markers have now been identified in giant cells around various bone biomaterials, much further research is needed to fully characterize them.
Therefore, research dating back over 20 years has hinted at the fact that giant cells around bone grafts are different from osteoclasts morphologically, enzymatically, or functionally, yet the ability to accurately characterize them accordingly by cell surface markers and receptors did not exist at that time. Furthermore, their role was much more frequently associated with a foreign body reaction as it relates to bone biomaterial integration. In this study, we present evidence that MNGCs may also serve as a bone-enhancing cell type similar to those found around atherosclerotic calcification.
Evidence that MNGCs induce vascularization and new bone formation
The role of macrophages and MNGCs around bone grafting materials has gained tremendous momentum with the advancements made in the field of osteoimmunology. 48 Despite the growing trend to study macrophage behavior and polarization around bone biomaterials, this review focuses only on MNGC function as it relates to their ability to induce vascularization and ultimately new bone formation (Table 1).
BMSC, bone marrow stromal cell; DBBM, deproteinized bovine bone mineral; MNGC, multinucleated giant cell; TCP, tricalcium phosphate.
Abshagen et al. were one of the first to show the implications of MNGCs in vascularization. 49 Owing to the importance of adequate vascularization during bone augmentation procedures, they studied the effects of NanoBone, a fully synthetic nanocrystalline bone graft, in a murine dorsal skinfold model. It was found that NanoBone markedly increased ingrowth of vascularized fibrous tissue associated with an increase in MNGC formation. 49
In 2011, Ghanaati et al. found that implanting fibroin scaffolds with previously harvested osteoblasts served to instruct host endothelial cells to migrate, proliferate, and initiate the process of scaffold vascularization. 50 It was concluded that the robust effect on scaffold vascularization at least appeared at the time to occur in concert with the pro-angiogenic stimuli arising from host immune cells, most notably macrophages and MNGCs. 50 It was later shown histologically by this same group in a split-mouth design in eight oral cancer patients that both NanoBone, as well as deproteinized bovine bone mineral (DBBM), led to MNGC formation with similar new bone volume after a 6-month healing period. 51 This study confirmed the presence of MNGCs in stable bone even in oral cancer patients. 51
In 2014, Tour et al. showed that bone marrow stromal cells (BMSCs) enhanced the osteogenic properties of hydroxyapatite scaffolds by modulating the foreign body reaction. 52 They loaded (BMSC)-incorporated biomimetic constructs composed of hydroxyapatite into rat calvarial critical-sized defects (8 mm). 52 Therefore, BMSC-loaded biomimetic constructs significantly enhanced bone repair by modulating the foreign body reaction and MNGC formation, 52 confirming the previous report by Ghanaati et al. 51 Others have also shown that cell incorporation into bone grafts leads to an increase in MNGC formation and subsequent angiogenesis.53,54
An interesting study revealed that when zinc was added to tricalcium phosphate (TCP) bone grafts, RAW264.7 cells cultured with different dosages of zinc supplements demonstrated that zinc could influence both the activity and the formation of MNGCs. 55 After a 12-week implantation period in the paraspinal muscle of canines, de novo bone formation and bone incidence increased with increasing zinc content (and MNGC formation), indicating that zinc incorporated in TCP can modulate bone metabolism and render TCP osteoinductive through MNGC formation. 55 This group later showed that using liposomal clodronate to inhibit the formation of macrophage/MNGC/osteoclast progenitors, no ectopic bone formation could take place around osteoinductive bone grafts at least partially implicating immune cells in the formation of ectopic bone. 56 Later, Davison et al. demonstrated that the scale of TCP biomaterial surface architecture could further affect MNGC/osteoclast cellular resorption. 57
Jensen et al. showed in both a human and animal study that the presence of MNGCs was frequently found around DBBM bone grafts.4,58 In a human study, it was found that long-term stability of bone augmentation was observed histologically in 12 human biopsies between 14 and 80 months after augmentation surrounded by many MNGCs displaying little to no bone resorption of such bone particles. 4 In a minipig study it was shown that two types of low-resorbing bone grafting materials formed MNGCs on the surface of bone grafts. 58 While MNGCs formed sealing zones and ruffled borders, both features of mature osteoclasts, MNGCs also demonstrated distinctly different histological features depending on the bone substitute material used with no evidence of particle resorption. 58
In a human study the material-specific tissue response to the synthetic, hydroxyapatite-based bone substitute material NanoBone was compared with that of DBBM. 59 In sinus cavities of 14 patients augmented with NanoBone and DBBM (split-mouth design), MNGCs were found on both bone graft particles with a significantly higher number of MNGCs and blood vessels in the NanoBone group compared to DBBM group. 59
The influence of granule size of two biphasic bone substitutes (BCP, 400–700 μm and 500–1000 μm) was evaluated on the induction of MNGCs and implant bed vascularization in a subcutaneous implantation model in rats. 60 Higher numbers of MNGCs were detected in the group with small granules starting on day 30, with higher vascularization. Furthermore, the results revealed that a synthetic bone substitute material could induce tissue reactions similar to those of some xenogeneic materials, thus pointing to a need to elucidate their “ideal” physical characteristics. 60 Another study showed that addition of monocytes to bone grafts led to their being involved in the tissue response to a biomaterial, however without marked changes in the overall tissue reaction. 61 Finally, high-temperature sintering of xenogeneic bone substitutes was found to increase MNGC formation associated with enhanced vascularization. 62 These results revealed that high heat treatment of xenografts led to an elevation in the inflammatory tissue response to the biomaterial and a combined enhancement in MNGC formation. 62 In that study, it was concluded that “further clarification of the differentiation of the multinucleated giant cells toward so-called osteoclast-like cells or foreign-body giant cells is needed to relate these cells to the physicochemical composition of the material.” 62
Interestingly, in 2015 Katsuyama et al. demonstrated specifically that MNGCs do not resorb bone but rather express M2 macrophage markers. 63 They report the critical findings that strongly suggest that implant failure due to bone loss likely resulted as a direct activity from osteoclasts and not MNGCs since they were unable to resorb bone. 63 This report highlights the fact that MNGCs can polarize toward M2 phenotypes with much further research necessary to characterize their behavior around bone biomaterials.
Role of MNGCs around dental implants
It remains of interest to note that to this day, very few studies beyond bone grafting materials have focused on MNGC formation surrounding dental implants. This may be due to the fact that along metal implant surfaces, observable differences between osteoclasts and MNGCs are not as obvious as around bone grafts where MNGCs can be distinguished based on their inability to resorb grafts. Since neither MNGCs nor osteoclasts can resorb metals, samples around dental implants are typically embedded in paraffin where surface marker characterization can be performed. As such, very little research has investigated MNGC versus osteoclasts with respect to dental implant biomaterials.
It remains interesting to note that to this day, some dental implants and other bone biomaterials are lost every year due to unknown reasons. It is hypothesized that such failures could be due to humoral immunity in the case of biomaterials, a field of research that continues to grow rapidly.5,64 This area of investigation has been one of the main topics of focus by a prominent research group in Sweden working in implant dentistry for many decades studying the foreign body reaction of host tissues to dental implants.65,66 These concepts will be discussed later in the section “Role of MNGCs in peri-implant infections.”
Based on questions emanating from this work, new parameters used to quantify the number and formation of MNGCs around dental implant surfaces termed MNGC-to-implant contact and quantified data in relation to bone-to-implant contact and peri-implant bone density have been established. 67 It was found that significant differences in MNGCs were present on all tested implant surfaces, yet this was not associated with an inflammatory cell infiltrate, fibrous encapsulation, or implant osseointegration in the miniature pig defect model. 67 Based on these results, it is difficult to assess the role of MNGCs/osteoclasts around dental implants as to date there has been no attempt to characterize these cells using cell surface markers. While other studies have shown that implant surface topography/composition has an influence on MNGC formation,68,69 more studies remain necessary.
Role of MNGCs in peri-implant infections
Most of the research to date investigating MNGCs around dental implants has dealt with the clinical condition of peri-implantitis.65,66,70 A group of researchers from Sweden has revealed the marked impact of MNGCs around dental implants during peri-implant bone destruction. In a publication reviewing the foreign body reaction to biomaterials, a hypothetical view on the mechanisms leading to buildup and breakdown of osseointegrated dental implant interfaces was proposed. 66 While the authors provide a view of biomaterial integration as it relates to a “foreign body reaction” of biomaterials into host tissues, it is difficult to gather a clear description/understanding of the role of MNGCs or FBGCs. In their proposed model (Fig. 3E, reprinted with permission), they theorize the mechanisms whereby invading bacterial pathogens induce the fusion of macrophages to FBGCs which then degrade alveolar bone tissues surrounding implants. 66 While such a pathway might be possible, it has more recently been shown that fusing macrophages into MNGCs (or FBGCs) are incapable of resorbing bone (Fig. 4), 42 thereby refuting the hypothesis that MNGCs or FBGCs are responsible for resorption of bone following pathogenic infection. It has further been shown that MNGCs have minimal capability to degrade bone compared to osteoclasts with a 40-fold decrease in efficiency. 42 It is therefore not plausible that these cells are directly responsible for alveolar bone degradation and are more likely to interact with other cell types during this process. This topic has been very superficially studied to date.
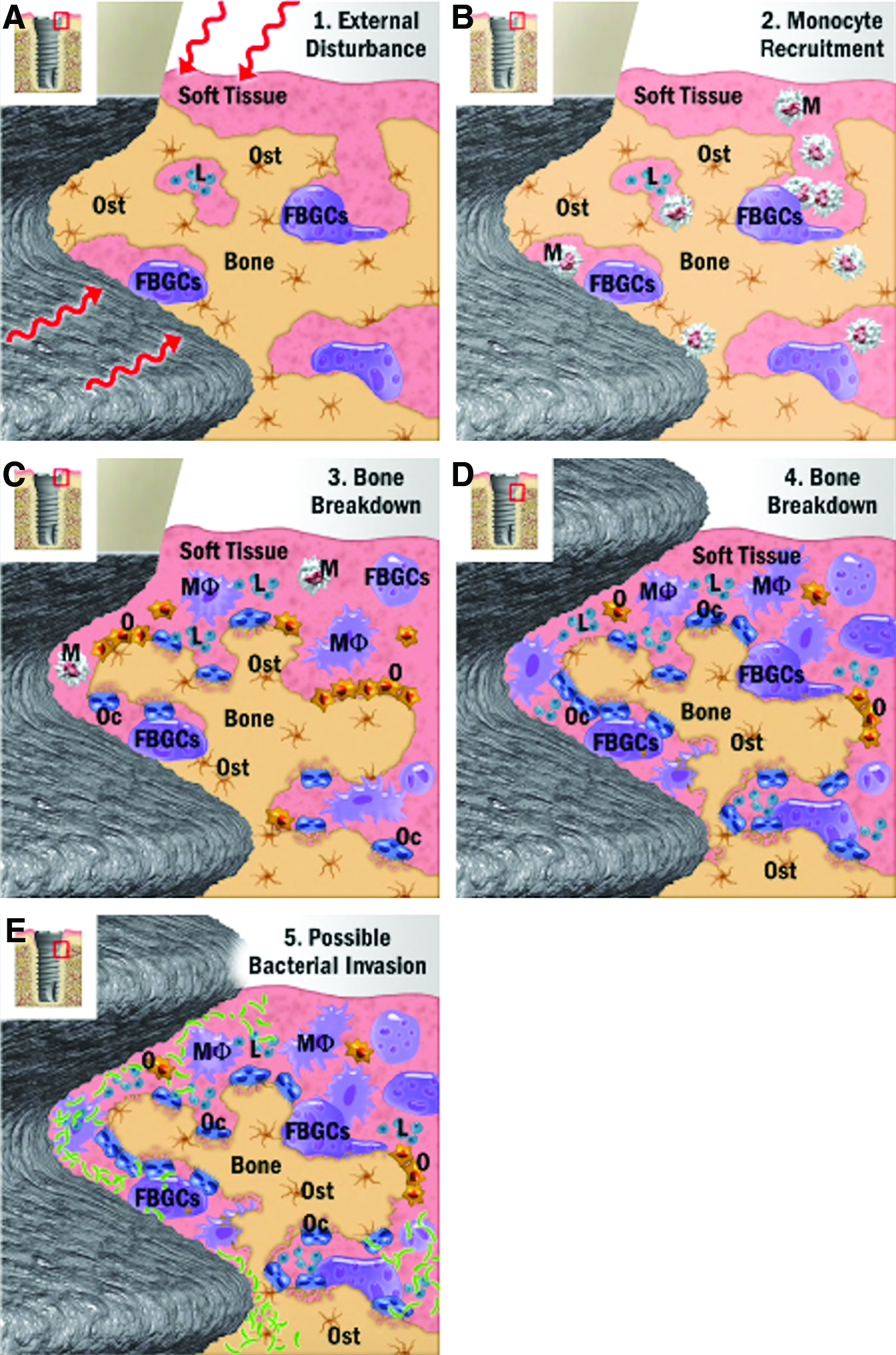
Proposed peri-implant defect progression from Trindade et al.
66
Notice the illustration that FBGCs are resorbing bone.
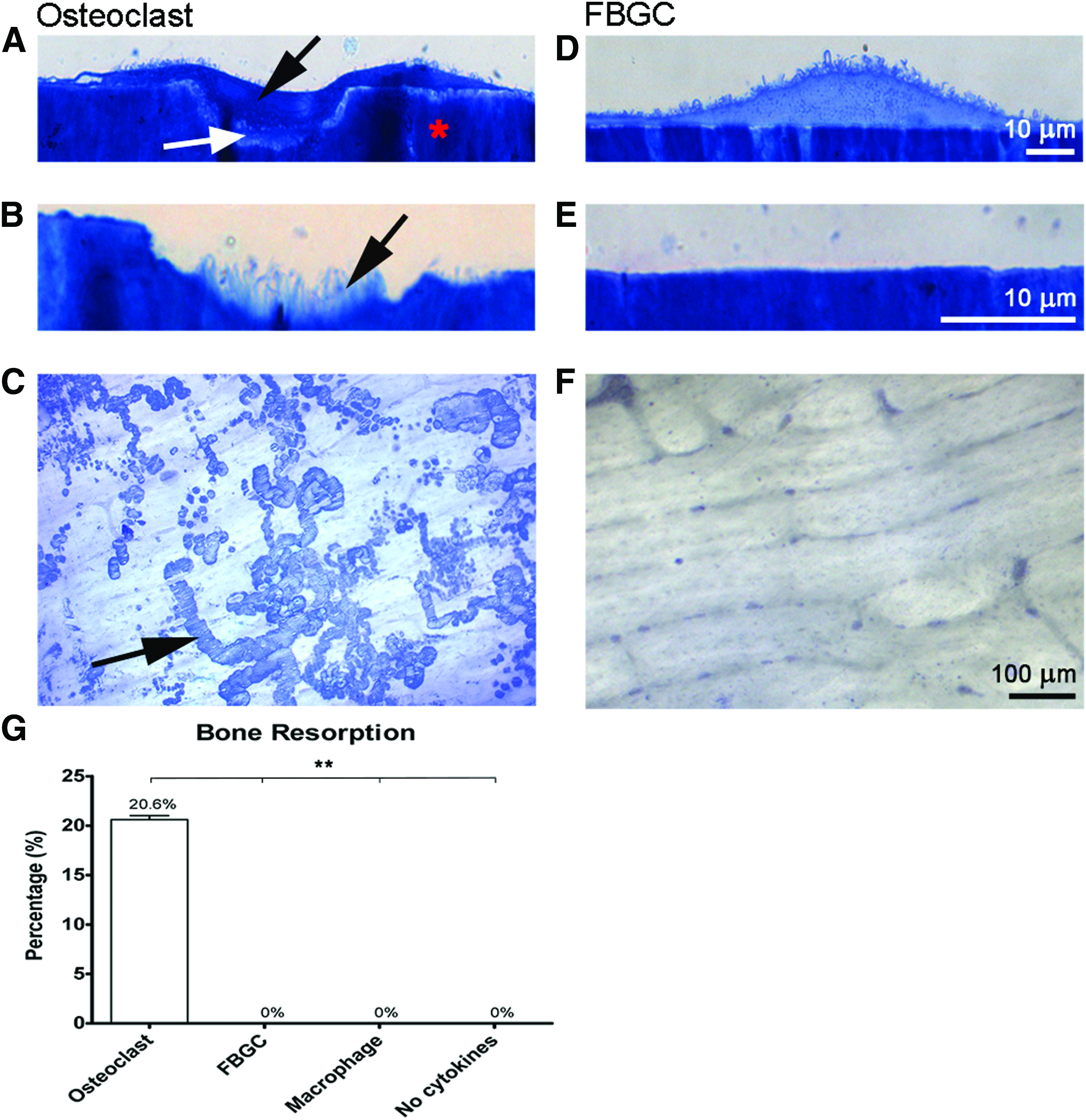
Bone resorption by osteoclasts and FBGCs by ten Harkel et al.
42
After 25 days, cells were stained with Richardson's staining solution
It goes without saying that the complex nature of MNGC biology would greatly benefit from a more accurate characterization, as well as description of their role in bone tissue hemostasis and disease. It is most likely that these cells are reversibly capable of shifting polarization in a similar manner to macrophages in response to changes in cytokines and various pathogens within their microenvironment. 71 Interestingly, recent findings have demonstrated that MNGCs are able to polarize toward wound healing M2-MNGCs thereby contributing to tissue wound healing and regeneration. 27 As it relates to dental implant biology, this field of research has thus far been left entirely unstudied.
Discussion and Future Research Outlooks
In summary, this review integrates current knowledge to date as it relates to MNGC behavior associated with bone biomaterials. While the role of MNGCs has largely been associated with a foreign body reaction over the years, a more contemporary view is that MNGCs do not prevent osseointegration of dental implants 67 and are capable of expressing M2-macrophage markers. 27 Therefore, based on MNGC fusion from M1 and M2 macrophages, we propose that MNGCs be assessed more specifically in future research endeavors as M1-MNGCs and M2-MNGCs (Fig. 5). In this way, their polarization and subsequent fusion from macrophages can be better characterized in health and disease.
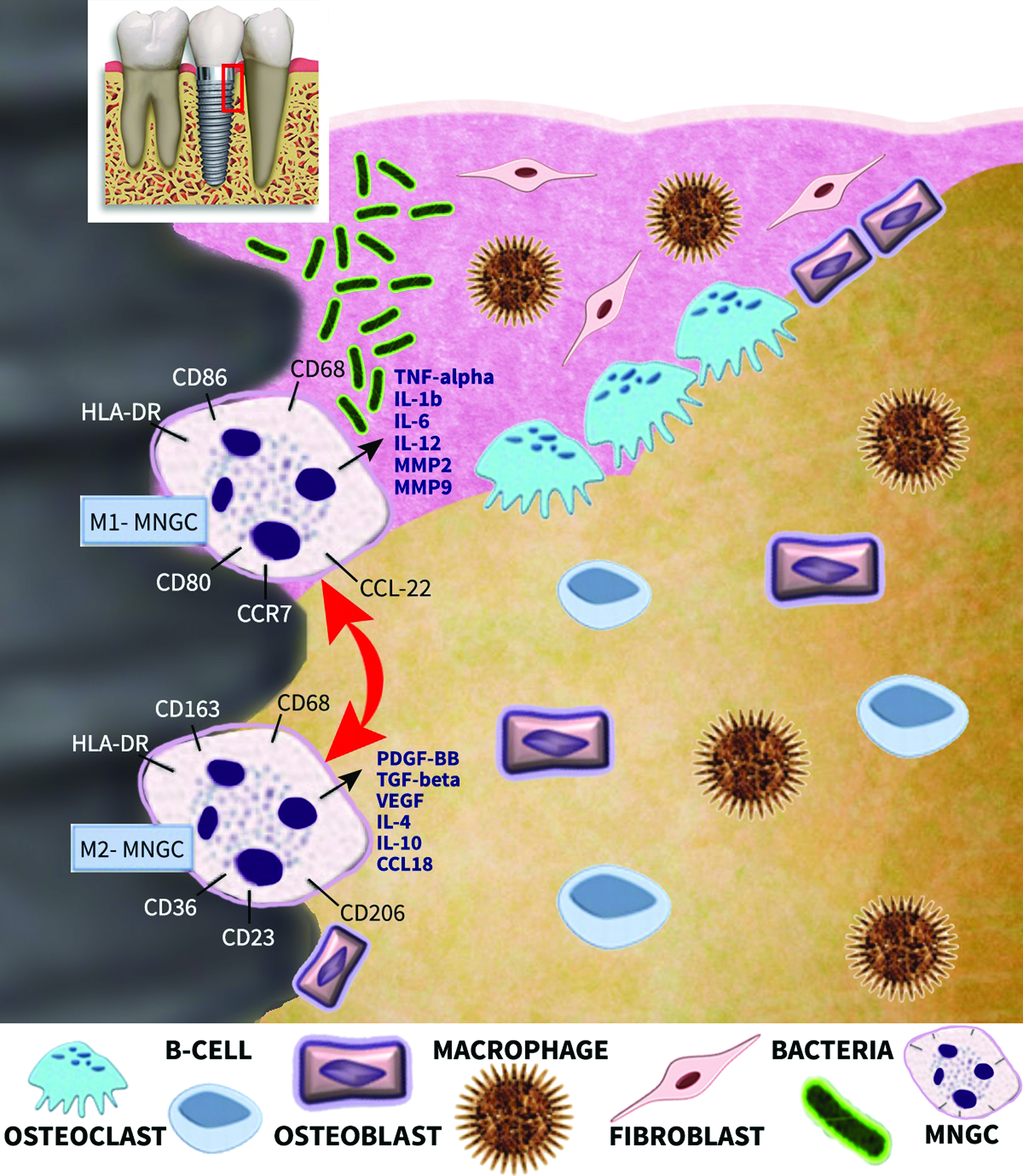
Proposed model of MNGC behavior during peri-implant infection. During bacterial invasion macrophages and MNGCs may polarize toward tissue-inflammatory M1-macrophages and M1-MNGCs. Interestingly, other macrophages found more coronal in stable bone may potentially be of M2-macrophage/M2-MNGC phenotype. An intriguing unanswered question is how these macrophages and MNGCs polarize during peri-implant breakdown and regeneration. Is it possible that MNGCs share similar properties as macrophages where they may potentially be capable of shifting polarization readily in response to their microenvironment? Color images available online at www.liebertpub.com/teb
The physiological behavior of MNGCs, especially as it relates to dental implants, is a growing area of research. While MNGCs are known to exist on various dental implant surfaces (at least during the osseointegration phase), one of the key differences between these tissues and others is that they are transmucosal and thus bear the risk of being invaded by bacterial pathogens found in the oral cavity (Fig. 6). If bacterial accumulation occurs on an implant surface, it is likely that both macrophages and/or MNGCs will polarize toward M1-macrophage and M1-MNGCs, respectively, creating an environment dictating tissue inflammation and further tissue breakdown. While but a hypothesis, it is now known that these cells are completely unable to resorb bone and therefore their interaction with other cells such as osteoclasts (direct interaction vs. indirect interaction through e.g., T-cells) is highly probable. How cell–cell communication occurs (through direct cell contact or paracrine activity) during this process also remains completely unstudied.
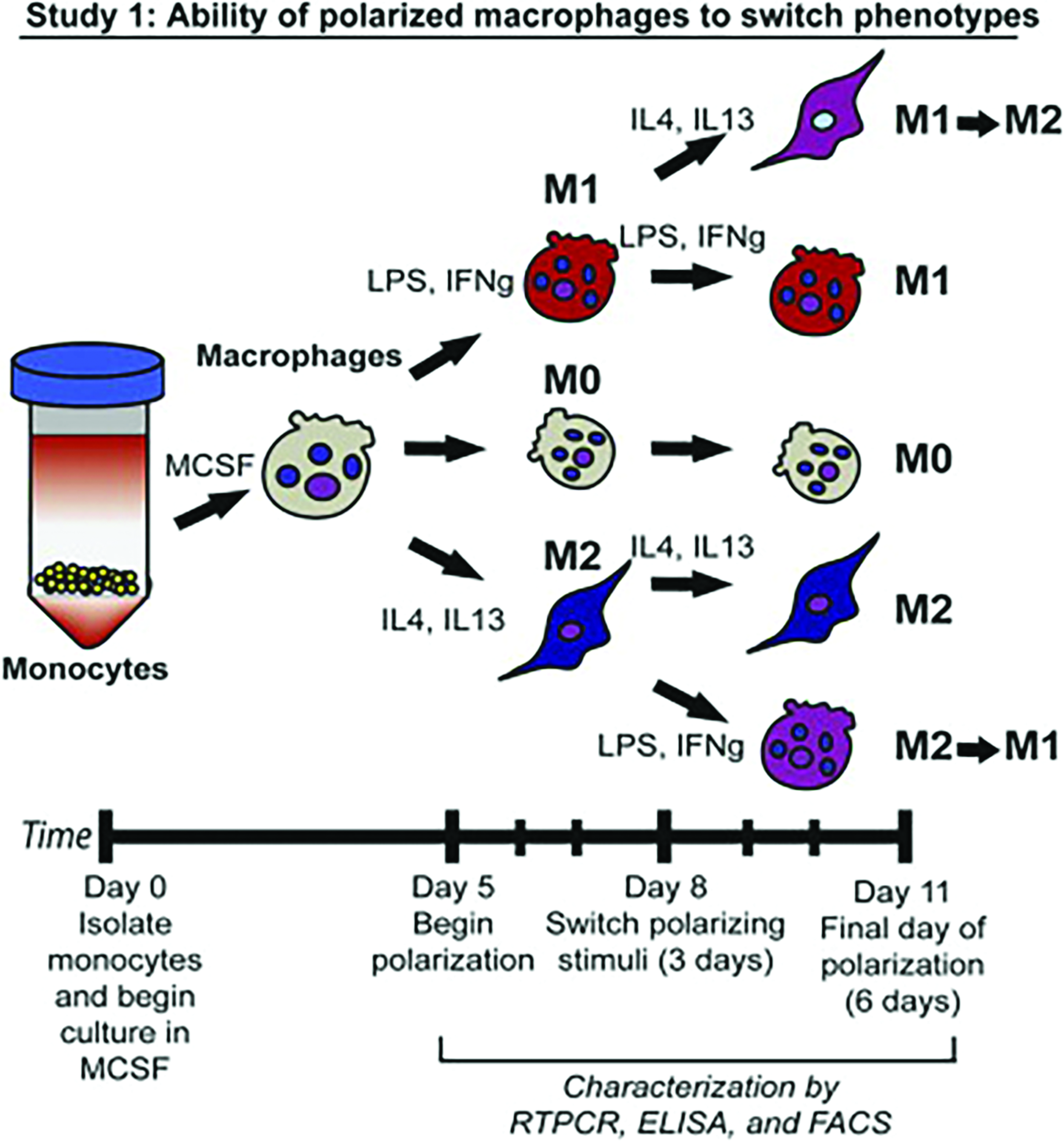
Study published by Spiller et al. 71 demonstrating the experimental design whereby full polarized M1 and M2 macrophages can reverse their polarization within 3 days. Study design. Study 1: Monocyte-derived macrophages were exposed to M1- or M2-polarizing stimuli for 3 days followed by polarizing stimuli of the other phenotype for an additional 3 days (M1→M2 and M2→M1). Unstimulated macrophages (M0) or macrophages cultured under M1- or M2-polarizing stimuli for 6 days (M1 and M2), with a media change at day 3, served as controls. Reprinted with permission. 71 Color images available online at www.liebertpub.com/teb
At the same time, it is known that MNGCs can also exist on the surface of fully integrated biomaterials even years after their implantation. These MNGCs are often seen surrounded by stable bone, and our research team has now shown that these cells express M2-related phenotypes (unpublished data). Therefore, evidence that MNGCs are both capable of acting as M1 tissue inflammatory and/or M2 tissue wound healing MNGCs in various bone environments should therefore be characterized more appropriately as M1-MNGCs and M2-MNGCs according to their role in tissue homeostasis. It is also interesting to point out that very similar to macrophage biology, MNGCs lining a dental implant surface could potentially coexist as M1-MNGCs at the coronal portion of the implant surface during periodontal bacterial invasion, yet also exist as M2-MNGCs at the protected apical portion of the implant not yet contaminated by bacterial invasion. While this hypothesis requires much further investigation, interestingly it has been shown by Spiller et al. that macrophages proficiently polarize from M1 to M2 and vice versa in as little as 3 days (Fig. 6). 71 It therefore remains of interest if MNGCs are capable of also shifting their polarization as readily with similar efficiencies. If so, one future strategy to resolve peri-implant infection might be to subsequently focus on macrophage/MNGC polarization which may further improve/enhance current strategies attempting to resolve peri-implantitis focused solely on strategies stimulating bone formation and suppressing bone resorption through growth factors and bioactive materials. While only a hypothesis, more evidence now points to the marked impact of immune cells in the field of bone biomaterial integration and homeostasis.
Another aspect that remains of interest in the dental field is the influence of macrophage/MNGC interactions to multiple biomaterials in small spaces. For example, tooth loss in the esthetic zone is commonly replaced by dental implants fabricated from titanium often requiring contour augmentation procedures with bovine-derived bone grafting materials and porcine collagen membranes. 72 Therefore, within this small regenerated area (often 1 mm in thickness), macrophages and MNGCs will be in contact with (1) a titanium dental implant, (2) a bovine bone graft, and (3) a porcine collagen membrane. It therefore remains unknown how polarization of macrophages/MNGCs is affected by the response of these cells to various materials simultaneously nor has their effect on cell–cell interactions within this microenvironment, including osteoblasts, osteoclasts, osteocytes, fibroblasts, endothelial cells, and leukocytes, been investigated. What would happen if one class of biomaterial favors M2-MNGC formation, whereas another favors M1-MNGC? How then would tissue integration occur in such a limited-sized environment? Furthermore, there are many patient-related risk factors such as (poly)-medications that may further increase susceptibility in some patients. The current influence of the large array of medications currently utilized on immune cells and their influence on bone biomaterial tissue integration is another field of research completely unstudied.
Interestingly, a category of bone diseases, including giant cell tumors, cherubism, and Noonan's syndrome, include large populations of MNGCs. These cells are associated with significant areas of osteolysis and their activities have been “reversed” with a disparate group of medications, for example, interferon, denosumab, and calcitonin. Very little is known to date regarding the specific characterization of these MNGCs as a potential third population of giant cells or M1-/M2-MNGCs. Therefore, it remains of interest to further investigate MNGCs found in the above-mentioned bone diseases with the appropriate markers comparing osteoclasts versus MNGCs and M1 versus M2 macrophage markers to better characterize their differences. A great deal of new information could be learned which may also further lead to new therapeutic strategies.
In summary, it is clear that much future research is needed to better understand MNGC formation and behavior in bone tissue homeostasis. Within the present article, we introduce the notion that MNGCs should no longer be referred to as “good” MNGCs or “bad” FBGCs and instead propose the necessity to better characterize them scientifically and appropriately as M1-MNGC and M2-MNGC accordingly. Future research investigating the factors influencing their polarization as a “center of control” is also likely to act as a key factor in the progression/resolution of various diseases such as peri-implantitis.
Footnotes
Acknowledgment
The authors thank Professor William Giannobile, University of Michigan, for critically reviewing the article.
Disclosure Statement
No competing financial interests exist.