
Abstract
Osteocytes, the most abundant cell type in mammalian bone, are generally considered as the terminally differentiated cells of osteoblasts that are progressively self-buried or passively embedded in bone matrix. Emerging evidence reveals the essential functions of osteocytes in bone homeostasis and mechanotransduction. However, our knowledge on osteocytes, especially their formation, remains scarce. In this regard, the current review mainly focuses on several key factors that drive the osteocytic differentiation of osteoblasts, that is, osteocytogenesis. Available literature has demonstrated the involvement of physicochemical factors such as matrix composition, oxygen tension, and mechanical stress in the osteoblast-to-osteocyte transition. During cell migration and matrix remodeling, the matrix metalloproteinase-dependent collagen cleavage would play an “active” role in maturation and maintenance of the osteocytes. Besides, some in vitro methodologies are also established to induce the transformation of osteoblastic cell lines and primary mesenchymal cells to preosteocytes through cell transfection or addition of exogenous molecules (e.g., fibroblast growth factor-2, retinoic acid), which could potentiate the effort to form functional bone substitutes through elevated osteocytogenesis. Thus, advances of new technologies would enable comprehensive and in-depth understanding of osteocytes and their development, which in turn help promote the research on osteocyte biology and osteopathology.
Introduction
R
As the outer layer of bone, cortical bone is composed of repeating units called osteons (Fig. 1A), in which the lacunar–canalicular network (LCN) (Fig. 1B–E) formed by the embedded osteocytes is responsible for maintaining the homeostasis of bone while sensing mechanical stimulation for coordinating various cascading activities such as osteogenesis and remodeling (Fig. 1F). Because of the unique structure of osteons and their well-connected osteocytic network, cortical bone has a slow turnover, implying a prolonged time for regeneration, especially for the large bone fractures. In this regard, creation of cortical-like bone, that is, the establishment of osteocytic network along with the formation of compacted structure, represents an effective strategy to repair large bone defects for prompt restoration of the physiological functions.
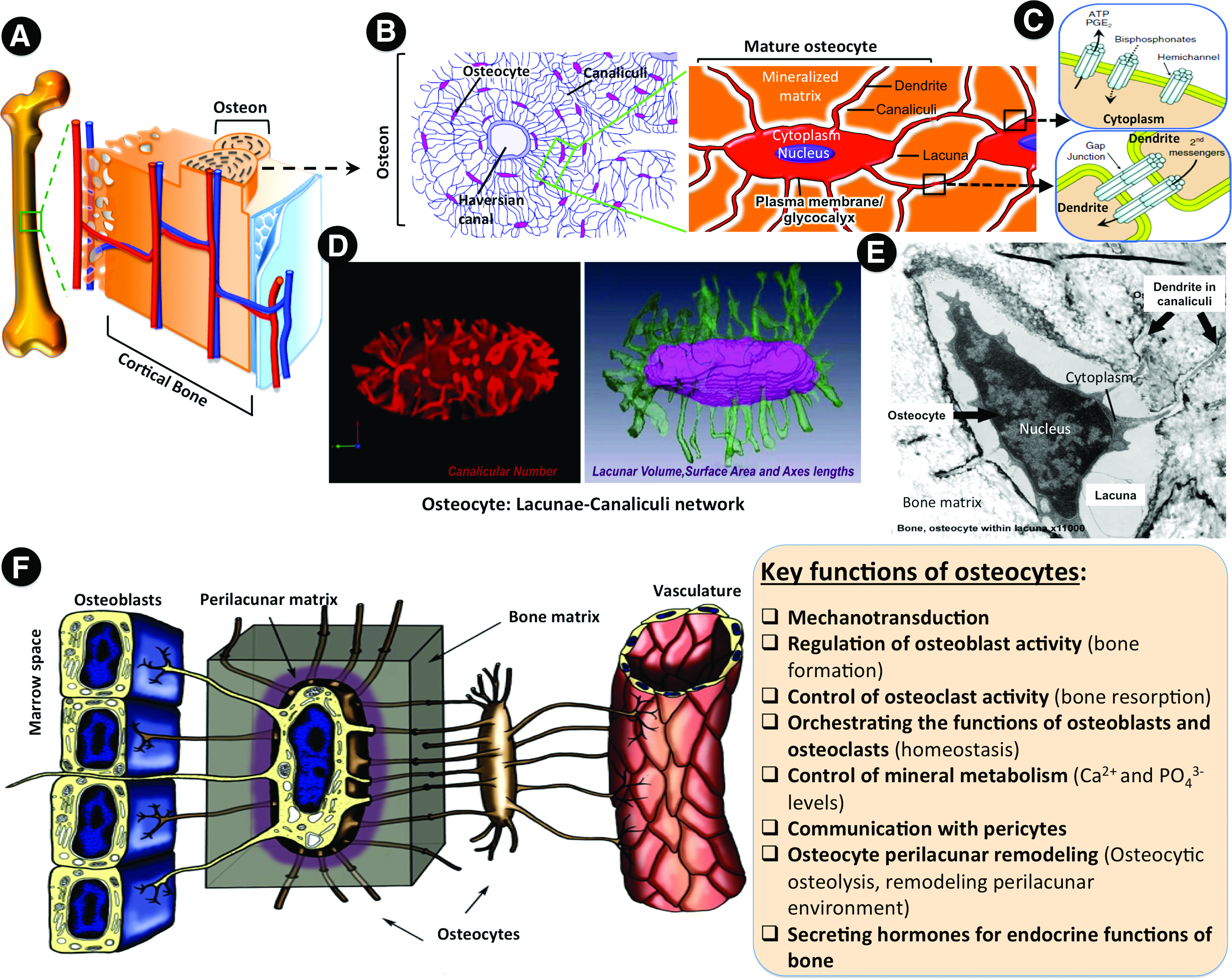
Osteocytes and the LCN.
In view of the difficulty in recapturing the spatial organization of cells and matrix of cortical bone with current bone tissue engineering approach, in-depth understanding of the associated cells, such as osteocytes and their development within the residing microenvironment, may help design a novel avenue to establish the desired physiological functions.
Osteocytes and Osteocytogenesis
Osteocytes, the longest-living and most abundant cellular component of mammalian bone (90–95%), reside in a three-dimensional-LCN (3D-LCN) surrounded by ossified matrix (Fig. 1B, E). Osteocytes are considered as the nonproliferative terminally differentiated cells, which are derived from osteoblasts of mesenchymal origin. 9 During bone formation, about 5–20% of mature osteoblasts are gradually embedded within the surrounding extracellular matrix (osteoid) that mainly consists of type I collagen (COL-1), and then transformed to preosteocytes or osteoid osteocytes. 10 Later, when the osteoid becomes mineralized, the preosteocytes mature into osteocytes. 11 This osteoblast-to-osteocyte transition is often referred to as osteocytogenesis. Due to their dormancy, osteocytes have long been considered inactive with limited physiological functions. However, increasing evidence has demonstrated that networked osteocytes can sense local compressive strain through the 3D-LCN, and consequently initiate strain-dependent new bone formation and homeostatic bone remodeling by osteoblasts (Fig. 1F).12–14 More details about their functions can be found in recent reviews.9,11
The osteocyte formation is accompanied by a striking cell morphology change, that is, from a polygonal shape to a dendritic/ramified one, which is the characteristic feature of osteocyte. 15 As a consequence, the cell body of mature osteocytes resides in the small lacunar spaces within mineralized osteoid, but they are well interconnected with neighbors (e.g., osteoblasts, ostoeclasts, and other osteocytes) through an extensive network of the long, slender cytoplasmic processes in the channels (canaliculi) connected by gap junctions. 16 Typically, the osteocyte-to-osteocyte distance is maintained within 20–30 μm for effective diffusion of molecules. 17 Moreover, during the osteocytic differentiation, cells greatly downregulate or switch off a multitude of key molecular markers specific to their preceding osteoblasts, such as Col-1, alkaline phosphatase (ALP), and osteocalcin (OCN). Meanwhile, they start to highly express some well-known osteocyte-specific markers, including connexin43 (CX43), podoplanin (also called as E11/gp38), dentin matrix protein 1 (DMP1), phosphate-regulating neutral endopeptidase on chromosome X (PHEX), matrix extracellular phosphoglycoprotein (MEPE), fibroblast growth factor 23 (FGF23), and sclerostin (SOST) (Table 1, Fig. 2). CX43 is associated with gap junction for maintenance of the intercellular communication and response 18 ; podoplanin (E11/gp38), as a marker of preosteocytes, involves in the development of dendritic processes 19 ; DMP1, PHEX, and MEPE are considered as the major regulators of mineralization20–22 ; FGF23 is responsible for phosphate absorption 23 ; SOST is thought to be uniquely expressed in mature osteocytes as a negative regulator of osteoblast differentiation. 24 Previous studies demonstrated that genetic ablation of these markers in the knockout animal models would result in defective osteocyte differentiation and abnormalities in bone morphogenesis.18,20,25
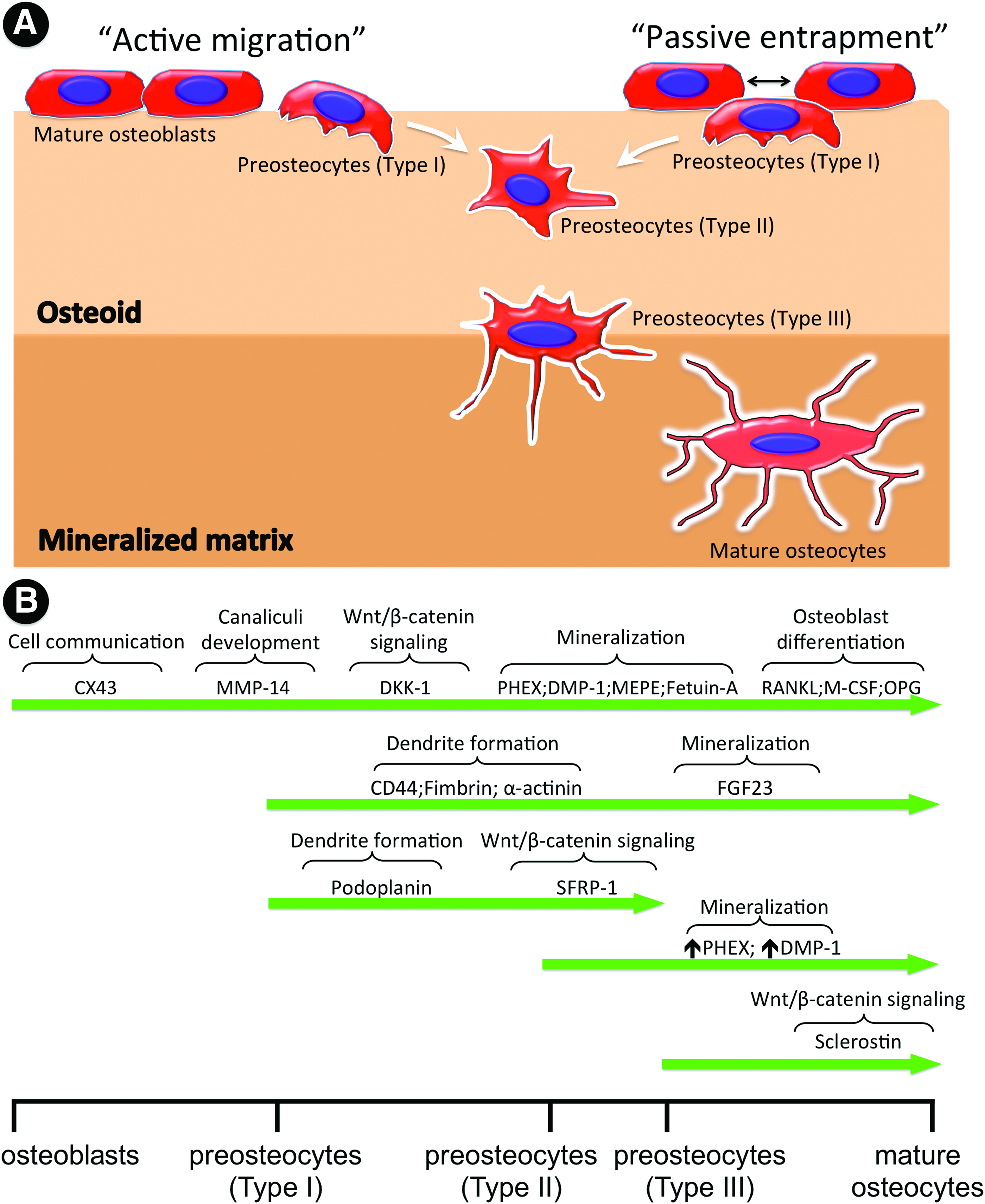
Osteocytogenesis and key molecular regulators.
COL-1, type I collagen; ALP, alkaline phosphatase; RUNX2, runt-related transcription factor 2; BSP, bone sialoprotein; OPN, osteopontin; OCN, osteocalcin; CX43, connexin43; DMP1, dentin matrix protein 1; PHEX, phosphate-regulating neutral endopeptidase on chromosome X; MEPE, matrix extracellular phosphoglycoprotein; FGF23, fibroblast growth factor 23; SOST, sclerostin; MMP, matrix metalloproteinase.
Despite the recognized role of osteocytes and 3D-LCN in bone function, the mechanisms behind osteocytogenesis remain poorly understood, mainly because of the difficulty of accessing osteocytes deeply entrapped in the mineralized bone matrix and their low mitogenic activity. 26 With the advances of high-resolution microscopic technologies and high-throughput molecular screening, as well as the development of cell- and tissue-specific transgenic animals, it becomes possible to identify the molecular expression patterns of osteocytes.15,21,27 Thus, this review would focus on the elucidation of several key regulatory factors that induce the osteoblast-to-osteocyte differentiation.
Physicochemical Factors Trigger Osteocytogenesis
In the natural circumstances, the osteoblast-to-osteocyte transition follows closely interregulated steps that correlate intimately to the changes of the cell surroundings (Table 2). Superficial osteoblasts are progressively self-buried or embedded by neighboring cells into osteoids, in which osteoblasts experience 3D environmental stimulation and commit toward the phenotype of preosteocytes (Fig. 2A). Further mineralization of the osteoids leads to the maturation of preosteocytes into osteocytes. 11 Gradual and accumulative mineralization of osteiods would result in the temporal and spatial alterations in matrix composition, oxygen tension, and mechanical stress, which in turn play a pivotal role in osteocytogenesis as described below.
2D, two-dimensional; 3D, three-dimensional.
Matrix mineralization and topography
As osteocytogenesis occurs concomitantly with the osteoid mineralization, numerous studies have been performed to understand the contribution of matrix mineralization to the transformation of osteoblast-to-osteocyte.28–32 Irie et al. found that disturbed mineralization of primary matrix resulted in immaturity of osteocytes with a restricted dendrite formation and the lack of the secretion of sclerostin, an osteocyte-specific marker in a rat model. 31 Prideaux et al. reported that the inhibition of matrix mineralization in preosteocyte-like cells (MLO-A5) by the addition of natural mineralization inhibitor (PPi) or omission of β-glycerophosphate (β-GP) would dramatically downregulate gene expression of the osteocyte-specific markers, such as E11, Dmp1, and Sost, but upregulate the expression of osteoblast makers, such as Col1α and Ocn. 32 It is suggested that impairment of mineralization would cause retention of late osteoblast stage and abrogate osteocytic differentiation. These studies have confirmed the driving function of matrix mineralization in osteocyte formation and maturation; however, its underlying mechanism is poorly known. It is reasonable to assume that the pronounced differences in fluid flow and oxygen diffusion between the mineralized and nonmineralized matrix, as well as the dramatic change of mechanical properties during matrix mineralization might be sensed by osteoblasts or preosteocytes, and further alter their gene expression pattern to initiate the osteoblast-to-osteocyte transformation. Moreover, another possibility is the presence of unknown mineralization sensors in preosteocytes, which might perceive specific factors related to matrix mineralization (such as DMP1) to induce osteocyte maturation. 31 Studies have shown the differential modulation of calcium signaling within osteoblasts and osteocytes especially with the presence of fluidic flow and hydrostatic pressure,33–36 indicating its possible participation in osteocytogenesis.
On the other hand, matrix organization, especially the arrangement of collagen fibers, could influence the behaviors of osteoblasts and the development of osteocytes. Our previous study found that matrix fibers with aligned orientation favored the osteogenic differentiation of mouse preosteoblast-like cells (MC 3T3-E1) for both two-dimensional (2D) and 3D culture, by the evidence of elevated ALP activity and mineral deposition. 37 Similar results were also recently reported by others. 38 In natural bone, the differences of matrix topography between woven and lamellar bone result in distinctive physiologic-type shape and orientation variations for osteocytes. In woven bone, randomly arranged collagen fibers lead to osteocytes with almost isodiametric shape; whereas in lamellar bone, thicker collagen bundles parallel to the lamellar plane induce the polarization of osteocytes with flattened oblate spheroidal shape. 39 Despite the distinctive morphology alteration, how matrix topography contributes to osteoblast–osteocyte differentiation remains poorly understood and requires further elaboration.
Oxygen tension
In native bone, mature osteocytes deeply embedded in the mineralized osteoid experience a much lower oxygen tension than those superficial osteoblasts, exhibiting the oxygen diffusion gradient as a result of the accumulated layers of mineralized bone matrix. 40 Initial efforts showed that compared with MC3T3-E1 preosteoblastic cells, MLO-Y4 osteocyte-like cells exhibited a significantly higher level of hypoxia-associated oxygen-regulated protein (ORP150), implying the potential role of oxygen tension in osteoblast-to-osteocyte transition.21,41 Hirao et al. demonstrated that low oxygen (hypoxia, 5% [v/v] O2) rather than normoxic (20% [v/v] O2) conditions could facilitate the transformation of MC3T3-E1 preosteoblasts to osteocytes in vitro by upregulating osteocyte-specific marker genes, such as Dmp1, Mepe, Fgf23, and Cx43. 42 In the ex vivo mouse calvarial culture, hypoxia increased the number of osteocyte-like cells that expressed DMP1 and MEPE proteins, and meanwhile encouraged their residing in the bone matrix. Intriguingly, under the normoxia culture ORP150 expression was only found in those mineralized nodules, in contrast to much abundant expression under a hypoxia, suggesting that the mineralization process might cause the development of hypoxic zones. Similarly, Zahm et al. affirmed the regulatory involvement of oxygen tension in preosteocyte maturation, in which the preosteocytic cell line of MLO-A5 displayed a reduced osteogenic activity, an essential event before osteocytic differentiation, under a hypoxic condition (2% [v/v] O2) in comparison to normoxia. 40 This differential regulation was closely associated to the activation of transcription factor hypoxia-inducible factor 1 (HIF-1). However, Montesi et al. suggested that an extreme hypoxia (e.g., 1% [v/v] O2) induced the apoptosis of osteocytes, which was partially counteracted by the upregulated ORP150 that is regarded to play a cytoprotective role in an oxygen-deprived condition. 43 Taken together, previous findings indicate that low oxygen tension inhibits the growth and differentiation of osteoblasts,44–46 whereas hypoxia promotes osteocytogenesis by specifically activating the hypoxia-related proteins (e.g., ORP150 and HIF-1) to induce osteocyte-specific gene expression.40–43 Moreover, recent studies also suggest that hypoxia or chemical mimicry of hypoxia may exert its effect on bone formation through canonical Wnt signaling pathway by regulating the cytosolic stabilization and nuclear accumulation of its signal transducer—β-catenin to subsequently induce β-catenin-driven gene transcription, as well as through BMP signaling pathway by mediating the expression of BMP antagonists (e.g., gremlin and noggin) and the phosphorylation of downstream Smad 1/5/8.47,48
Mechanical stress
Osteocytes, as the major mechanosensor could detect the changes of mechanical stimuli and subsequently regulate the activities of osteoblasts and osteoclasts, resulting in bone formation or remodeling. 49 On the other hand, mechanical stimuli also serve as an important trigger for osteocytogenesis. Frank et al. claimed that mechanical stimulation could be used not only for a better understanding of the effects of frequent mechanical stress on the fate of hMSCs, but also for synchronized differentiation of adult stem cells to a specific lineage. 50 Zhang et al. found that the shear stress of the fluid flow upregulated the expression of early osteocyte marker E11 and promoted the dendrite elongation of osteocyte-like cells of MLO-Y4 in vitro, 19 and compressive loading increased podoplanin (E11) expression in osteocytes near the bone surface in vivo, indicating the critical role of mechanostimulated E11 expression in osteocytogenesis. 51 To understand its molecular basis, Hang et al. studied the differential involvement of A-Raf and C-Raf (the upstream mediators of extracellular signal-regulated kinases [ERK]/mitogen-activated protein kinases [MAPK] pathway, also known as the Ras-Raf-MEK-ERK pathway) in the mechanoregulated osteogenesis, and revealed that mechanosensitive cation channel (P2X7)-activated A-Raf was specifically responsible for the mechanical stretch-induced osteogenesis, whereas C-Raf contributed to antiapoptotic protection and promotion under the low-magnitude mechanical stretch. 52 With the increased deposition of ossified matrix, more osteoblasts would experience the transformation into osteocytes. Moreover, mechanical loading also imposes a profound influence on the osteocyte viability. 51 For example, Aguirre et al. showed that the prevalent increase of osteocytic apoptosis was associated with the lack of mechanical stimulation in the tail suspension test of a murine model 53 ; however, the mechanical stress from fluid flow and stretching at the physiological levels could effectively prevent the osteocyte apoptosis in vitro.54,55 These findings have suggested that the mechanical stress-induced osteocytogenesis and antiapoptosis are driven by intracellular signaling pathways (e.g., ERK/MAPK pathway), as mechanical forces can be transmitted by transmembrane integrins to cytoskeletal proteins and catalytic molecules (e.g., Src kinases), and finally induce nuclear translocation of ERKs to regulate the transcription of osteocyte-specific genes (e.g., E11) both in vitro and in vivo.51,52,54 Moreover, it is generally believed that as an important mechanosensor, osteocyte has the capacity of translating mechanical stimuli into biochemical signals to affect bone formation and remodeling through Wnt/β-catenin signaling pathways.56–58 Regarding the mechanosensitivity of osteocytes to various mechanical stimuli and the corresponding recruitment of osteoblasts and osteoclasts for bone formation and remodeling, a comprehensive review was provided by Klein-Nulend et al. 59
Collagenolytic Activities Promote Osteocytogenesis
Apart from the well-described theory of “passive” entrapment of osteoblasts for osteocytogenesis, recent work has also highlighted a more “active” role of osteoblasts and preosteocytes in the osteocytogenesis (Fig. 2A).60,61 Robin et al. observed the migration of osteoblasts into the osteoid-like dense collagen matrices over a large distance, accompanied with a decrease of collagen synthesis, but upregulated expression of matrix metalloproteinases (MMPs) (MMP-2, 13, and 14), which are a family of metal-dependent endopeptidases and responsible for the cleavage of collagens. 61 Such an observation implied that those osteoblasts, which migrated into collagen matrices through matrix degradation, would eventually transform into the osteocytes, as evidenced by the complete cell burial in the collagen matrix, the formation of lacunae, and the presence of cellular pseudopods. As such, it is suggested that osteocyte formation in natural bone can also be considered as an active invasive process that requires the MMP-dependent cleavage of collagen. In general, MT1-MMP is considered to play an important role in bone remodeling and matrix formation.62–64 In the MT1-MMP-deficient murine models, Holmbeck et al. observed a significant reduction of the osteocyte projection, which depended on the formation of canaliculi and were promoted by collagen cleavage. 65 This means that disruption of the MMP-dependent collagenolytic activity could exert a detrimental effect on osteoblast-to-osteocyte transition. Indeed, Zhao et al. detected the increased osteocyte apoptosis in Col1a1r/r mutant mice, in which type I collagen was resistant to collagenase. 66 Moreover, with the involvement of MMPs, PHOSHO1, and CX43, osteocytes are capable of directly remodeling their perilacunar surroundings (i.e., osteocytic osteolysis), which appears to be associated with the development of osteocyte phenotype.9,67 These studies have demonstrated that MMP-dependent collagen cleavage/breakdown is essential for the formation and maintenance of osteocytes.
Exogenous Molecules Stimulate Osteocytogenesis In Vitro
Various studies have so far identified multiple intrinsic markers that are specifically expressed in osteocytes; however, exogenous molecules with the potential to induce osteocytogenesis remain elusive. Recently, some efforts have been made to use the in vitro 2D cell culture models to determine some exogenous molecules, such as oncostatin M (OSM), FFG-2, and retinoic acid, with the inductive capacity to transform osteoblasts into osteocyte phenotype. The regulatory mechanism is further elaborated below.
Oncostatin M
OSM is a cytokine that is produced within the bony microenvironment by cells of both mesenchymal and hematopoietic origin and belongs to the interleukin (IL)-6 family with proapoptotic effects on multiple cancer cells.68,69 Besides their noted involvement in cancer and other disorders,70,71 OSM also plays the role in bone metabolism by direct and indirect participation through two independent receptor/ligand complexes: OSM receptor:gp130 and leukemia inhibitory factor receptor:gp130.72,73 The former one directly stimulates the osteoblast mineralization and differentiation as well as the osteoblastic commitment of mesenchymal stem cells and osteoclastic formation, whereas the latter is mainly involved in the suppression of sclerostin production, a bone formation inhibitor. More details on the endogenous OSM-regulated cellular activities can be referred to the insightful review. 73 On the other hand, it would be of particular interest in understanding the regulatory role of OSM in osteocytogenesis. Brounais et al. constructed the transgenic primary osteoblastic and osteosarcoma cells through transduction with lentiviral particles encoding OSM (lvOSM) for stable expression of OSM in vitro. 74 The long-term exposure to OSM could induce an osteocyte-like differentiation as evidenced by the decrease of osteoblastic markers, the morphology change to stellate shape with cytoplasmic processes, and the increased expression of osteocyte-specific markers (Sost, E11, and Phex). The cells undergoing OSM-induced differentiation did exhibit preosteocyte phenotype; however, they had a short lifespan and were more sensitive to apoptosis induced by the kinase inhibitor Staurosporine (STS).
Fibroblast growth factor-2
FGF-2 is recognized as a pluripotent growth factor that is involved in angiogenesis and wound healing and exerts a great impact on bone formation and fracture repair.75–77 The involvement of FGF-2 in bone formation, bone resorption, and downstream signaling pathways as well as the role of exogenous FGF-2 in fracture healing can be found in the recent review. 78 In addition to its regulatory function in osteoblast differentiation, 79 Gupta et al. found that under the in vitro cell culture conditions, the treatment of MC3T3-E1 preosteoblasts and primary bone marrow stromal cells with exogenous FGF-2 could cause a striking cell morphology change from an osteoblast-like spindle shape to a dendritic osteocyte-like morphology only after 8-h culture, and also dramatically promote the gene expression of osteocyte-specific markers (Dmp1, E11, and Cx43). 80 However, the FGF2 treatment, on the other hand, also inhibited MLO-Y4 osteocyte-like cells to express Sost, a marker uniquely expressed in mature osteocytes to function as a negative regulator of bone formation. 24 Collectively, the results suggest that FGF-2 could induce the transition of osteoblastic cells to preosteocytes through regulating gene expression patterns related to osteocytes; however, it might be detrimental to maturation and maintenance of osteocytes.
Retinoic acid
Retinoic acid, as an active metabolic product of vitamin A, can regulate gene transcription by binding to the nuclear receptors (RARs). RARs then bind to DNA as heterodimers with the retinoid X receptors, ultimately leading to the modulation of retinoic acid (RA)-responsive target genes.81,82 The involvement of various RA receptors in bone formation can be found in a very recent review. 83 In vitro analyses of the effects of RA reveal very contradicting observations on the maturation and mineralization of osteoblasts.84–87 Actually, both stimulatory and inhibitory effects have been observed, depending on the cell lines used and the culture conditions. However, it seems evidently clear that RA could accelerate the maturation of murine preosteoblasts 86 and promote its transition to preosteocytes. 88 It was found that external supplement of RA to the culture of mouse (MC3T3-E1 cells) and human osteoblasts (human fetal osteoblasts, hFOBs) in 3D collagen gel could modulate the cell number and function. In particular, RA significantly (p < 0.001) reduced cell numbers (hFOBs, MC3T3-E1). In MC3T3 cells, RA decreased IL6 secretion, and meanwhile significantly increased RANKL expression and stimulated FGF-23 expression and secretion in MC3T3-E1 cells, confirming the differentiation into osteocytes. 89 Similarly, Mattinzoli et al. showed that the addition of RA (5 μM) could induce both MC3T3-E1 osteoblasts and primary calvarial cells to differentiate toward the osteocyte lineage in a 10-day culture, as confirmed by the ramified (dendritic) cell morphology, the reduced matrix mineralization, and the upregulation of osteocyte-specific marks (Dmp1, Phex, Fgf23, and Sost). 88 This study suggested that RA might be a potent molecule to induce osteocytogenesis with in vitro cell culture. Clinical evidence has shown that high levels of serum retinol are associated with thinning of bone cortex and increase the risk of bone fractures in humans,90,91 indirectly suggesting the stimulatory effect of RA on the osteocytogenesis of osteoblasts. Furthermore, a very recent effort made by Jeradi and Hammerschmidt using zebrafish as the model has demonstrated that RA-induced premature osteoblast-to-preosteocyte transition has multiple effects on calvarial development, that is, during the calvaria growth, the RA-regulated osteoblast deprival would decrease the osteoid production and thereby lead to smaller and thinner calvaria, whereas the gained preosteocytes could increase osteoclast stimulation and subsequently cause calvarial fragmentation. 92 Similar effect was also observed with periosteal bone. 93
Conclusion and Future Perspectives
With the established evidence, we now have a better understanding of several key factors that regulate the osteoblast-to-osteocyte transition. In combination with known physiological triggers (e.g., matrix mineralization, low oxygen tension, and mechanical stress), the established cell transfection approach (e.g., OSM-transgenic cells) and the identified exogenous supplemental molecules (e.g., FGF-2, retinoic acid) 88 could provide more interventions and options to modulate osteocytogenesis. These in vitro induction methods hold particular promise in yielding a large quantity of osteocyte-like cells along with the successful isolation of primary osteocytes. However, the cells induced by in vitro exogenous molecules often exhibit preosteocyte-like phenotype. As noted, it remains a challenge to induce the development of mature osteocytes with physiological functions under the conventional 2D in vitro culture conditions, despite some success in patterning osteocytes through biomolecule immobilization (Fig. 3A). 94 In recognition, increasing attention has shifted toward 3D cell culture systems, by using loose or dense collagen hydrogels,60,61 calcium phosphate particles, 95 commercialized 3D polystyrene scaffolds, 96 and nanofiber-enabled layer-by-layer cell assembly, 97 to recapitulate the architectural complexity of osteoid, in which osteocytes are completely embedded within the 3D matrix environment (Fig. 3B–D). Compared with 2D culture, only 3D culture allows for reaching the maturation stage as osteoid-osteocytes. Furthermore, considering the essential roles of mechanical stimuli in osteocytogenesis, various mechanical loadings, such as cyclic biaxial stretching,52,54 stimulated microgravity (rotation), 96 cyclic hydraulic pressure, 98 and steady or pulsating fluid flow,55,99,100 have also been introduced into the in vitro culture system. It is worthy of noting that isolated osteocytes cultured under the 2D conditions evidently showed reduced sensitivity to mechanical stimuli, which may largely result from the dramatic cell morphology changes from 3D ellipsoid shape to 2D flattened one.59,101 As such, it is reasonable to speculate that the incorporation of 3D culture with mechanical stimuli in in vitro 3D tissue-engineered bone constructs may better mimic the in vivo physiological circumstances and therefore facilitate the maturation of osteocytes. While osteoblasts assure the production of bone matrix (osteoid), preosteocytes help the mineralization.30,51 All these findings indicate that timely commitment of each stage is essential to achieve the desirable physiological functions of bone. Early occurrence of osteocytogenesis would leave insufficient time for the production of bone matrix; otherwise, delayed mineralization would cause poor mechanical strength.102,103 Future efforts are required to explore the comprehensive effect of each factor, decipher the underlying mechanism, and identify more exogenous molecules with osteocytic inducibility. Thus, it will enlighten us to rationally combine physicochemical factors and exogenous molecules to construct in vitro osteocyte culture models as well as cortical bone-like constructs. In view of the potential advantages such as low setup cost, easy manipulation, high repeatability, and reproducibility, the in vitro 3D culture models hold great promise in scientific research on osteoblast-to-osteocyte transformation, osteocyte biology, and osteopathology, as well as in translational utility for drug screening and the creation of functional bone grafts for reconstructive surgery.
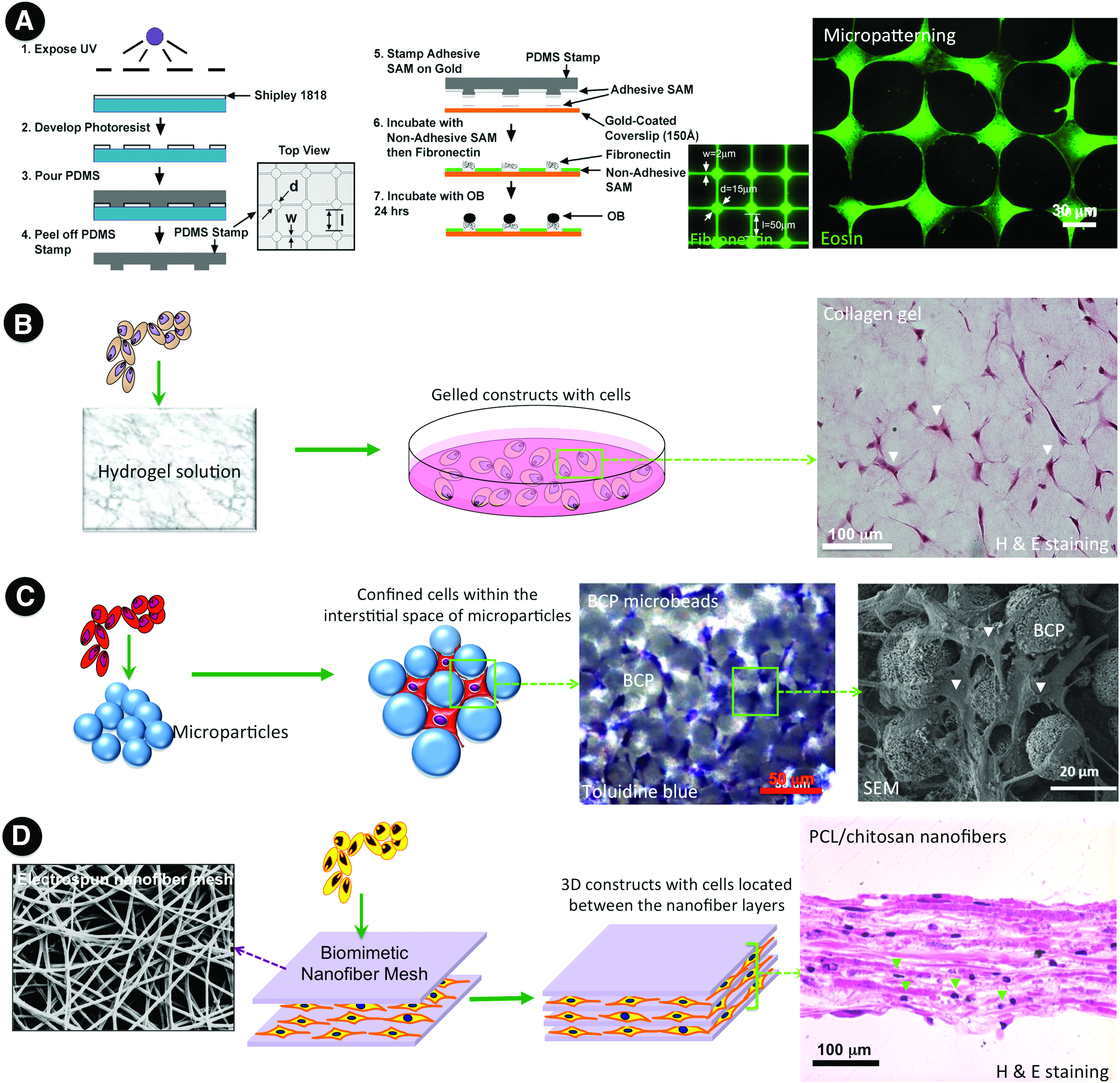
Various possible engineering approaches toward the establishment of osteocytic networks.
Footnotes
Acknowledgments
This investigation was financially supported by the National Natural Science Foundation of China (31400819, 31670985), National Science Foundation (NSF)-DMR (1508511), NIAMS (1R01AR067859), and NSF-ICorps (IIP 1637765).
Disclosure Statement
No competing financial interests exist.