
Abstract
Volumetric muscle loss (VML) is a debilitating condition wherein muscle loss overwhelms the body's normal physiological repair mechanism. VML is particularly common among military service members who have sustained war injuries. Because of the high social and medical cost associated with VML and suboptimal current surgical treatments, there is great interest in developing better VML therapies. Skeletal muscle tissue engineering (SMTE) is a promising alternative to traditional VML surgical treatments that use autogenic tissue grafts, and rather uses isolated stem cells with myogenic potential to generate de novo skeletal muscle tissues to treat VML. Satellite cells are the native precursors to skeletal muscle tissue, and are thus the most commonly studied starting source for SMTE. However, satellite cells are difficult to isolate and purify, and it is presently unknown whether they would be a practical source in clinical SMTE applications. Alternative myogenic stem cells, including adipose-derived stem cells, bone marrow-derived mesenchymal stem cells, perivascular stem cells, umbilical cord mesenchymal stem cells, induced pluripotent stem cells, and embryonic stem cells, each have myogenic potential and have been identified as possible starting sources for SMTE, although they have yet to be studied in detail for this purpose. These alternative stem cell varieties offer unique advantages and disadvantages that are worth exploring further to advance the SMTE field toward highly functional, safe, and practical VML treatments. The following review summarizes the current state of satellite cell-based SMTE, details the properties and practical advantages of alternative myogenic stem cells, and offers guidance to tissue engineers on how alternative myogenic stem cells can be incorporated into SMTE research.
Introduction
V
VML causes chronic pain, impaired function, and reduced mobility. A study by Corona et al. 5 found that VML was directly responsible for disability in 8% of all disabled veterans surveyed, and within a group of veterans with type III open tibia fractures, VML was an important contributor to 65% of the permanent disability they experienced. Corona et al. also found that the average lifetime disability cost for surveyed veterans with VML in the general disability population was $341,300 per person. 5 Compounding this problem, current surgical procedures to treat injury-induced VML are suboptimal at best, and full structural and functional recovery are rare. 6
The most common procedure that is currently used to treat VML is muscle flap surgery, 6 which involves transferring autologous tissue with intact blood and nerve supply from a donor site into the patient's injury site. Typically, this tissue is rotated from an adjacent donor site directly into the injury site; however, in the case of free functional muscle transfer, autologous donor muscle is transplanted from a site not immediately adjacent to the patient's injury, and the arteries, veins, and nerves are cut at the donor site and sewed into the injury site. 6 Muscle flap procedures are severely limited in terms of the amount of tissue that can be taken and rarely result in full functional recovery. 6 Donor site morbidity, which refers to any complication in the patient's donor site as it heals, is also a significant concern with these procedures. 6
Composite tissue allografts (CTAs) have been used successfully in recent years for face, hand, larynx, and abdominal wall transplants,7–12 and they may emerge as a more popular surgical treatment for VML in the coming years. CTA procedures transplant combinations of skeletal muscle, skin, bone, and tendon from an organ donor that is typically not genetically identical to the patient. More tissue can be implanted using CTA, but this procedure also carries significant negative side effects. Patients require lifelong immunosuppression, and tissue rejection, malignancies, and infections are serious concerns.
Because of the complications and inadequacies associated with current VML surgeries, there is intense interest in devising better treatment options for VML. In more recent years, skeletal muscle tissue engineering (SMTE) has emerged as a promising alternative to traditional VML surgical procedures. Generally, SMTE protocols start with a biopsy to isolate a stem cell population with myogenic potential. After the stem cells are allowed to proliferate, they may be differentiated in vitro. Depending on the specific protocol used, the cells can be seeded onto a scaffold for structural support13–15 or allowed to develop into a scaffoldless skeletal muscle tissue before implantation. 16 The engineered tissue is implanted back into the VML injury site to restore structure and function. Vascularization of engineered skeletal muscle tissues is critical for proper functioning in vivo, and is thus a burgeoning field of study, with several published methods to date.17–21 As summarized in a review of engineered skeletal muscle techniques by Bian and Bursac, 22 innervation methods are also being developed for SMTE, as innervation is essential for the development of functional, nonatrophic skeletal muscle.
As an alternative to the general SMTE process, some laboratories have experimented with decellularization techniques that use biologic scaffolds composed of extracellular matrix (ECM) proteins. When implanted into a VML site, ECM scaffolds encourage some degree of stem cell recruitment and de novo fiber regeneration without a prior biopsy and in vitro cell culture. 23 These methods rely on ECM's ability to create an environment for wound healing that minimizes infection, promotes moisture balance, and regulates cell behavior to promote tissue repair. 24 However, Garg et al. 25 found that using an ECM scaffold alone without a stem cell source was unable to recruit enough host stem cells in vivo to promote sufficient de novo fiber regeneration for functional recovery, and most SMTE laboratories currently use stem cells in their protocols.
Developing a successful SMTE protocol relies on choosing the proper stem cell starting source, and making this determination requires a detailed understanding of the properties of myogenic stem cells. Ideally, a stem cell population for SMTE should be easy to isolate, easy to purify, easy to expand in vitro without sacrificing myogenic potential, and able to properly fuse with existing skeletal muscle tissue in vivo to contribute to structural and functional VML recovery.
Because satellite cells are the native stem cell precursor to skeletal muscle, they are the most common starting source for SMTE. However, there are difficulties associated with using satellite cells. Satellite cell isolations are laborious, time-consuming, and variable between research laboratories. There is no consensus as to whether purification is required, but in protocols that use some degree of purification, the purification process can be challenging, and currently used methods are inconsistent, wasteful, or impractical. Looking toward personalized medicine applications, there are concerns as to how an adequate satellite cell population could be obtained. Acquiring enough satellite cells from a small skeletal muscle biopsy to produce a sufficiently sized tissue is challenging, yet a large skeletal muscle biopsy would not be clinically feasible because of the potential irreversible injury at the biopsy site. Senescence of satellite cells as they are expanded in vivo causes them to lose proliferative capacity, which reduces their ability to contribute to myogenesis over time.26,27 More generally, growing skeletal muscle in vitro is also challenging because there is no devoted collection of growth factors whose only purpose is to induce skeletal muscle growth, whereas, for instance, bone and cartilage tissue engineering methods have had great success using bone morphogenic proteins (BMPs). 28
To overcome the problems currently associated with satellite cell-based SMTE, tissue engineers should investigate the best stem cell source for SMTE. Alternative varieties of myogenic stem cells, including adipose-derived stem cells (ADSCs), bone marrow-derived mesenchymal stem cells (BM-MSCs), perivascular stem cells (PVSCs), and induced pluripotent stem cells (iPSCs), may offer attractive practical advantages over satellite cells, and should thus be explored in future SMTE studies in addition to satellite cells, to advance SMTE toward highly functional, safe, and clinically feasible VML treatments. Before discussing these alternative stem cells, it is first important to understand satellite cell properties and the current state of satellite cell-based SMTE.
Satellite Cells
Satellite cells are located in all skeletal muscle tissues beneath the basal lamina and outside the surrounding myofiber plasma membrane,29,30 and are the primary contributors to skeletal muscle regeneration. In fact, Lepper et al. 31 showed that satellite cells are absolutely required for normal muscle regeneration in vivo. Normal satellite cell myogenesis (Fig. 1) in response to injury begins when satellite cells are activated by local signals and upregulate the transcription factor myogenic differentiation 1 (MyoD) and myogenic regulatory factor 5 (Myf5), thereby committing to a myoblastic lineage and becoming myoblasts32,33 in a process termed induction. Myoblasts proliferate and differentiate into myocytes, which fuse into multinucleated myotubes after upregulating the transcription factor myogenin in a process called terminal differentiation. Alternatively, myoblasts can directly fuse with damaged myofibers in vivo. After terminal differentiation, myotubes can mature into myofibers.

Pathway of normal satellite cell myogenesis. Satellite cells express Pax7 and can commit to myoblasts in a process called induction or can self-renew. Myoblasts express Pax7, transcription factor MyoD, Myf5, and the myogenic protein desmin, and can differentiate into myocytes, which express MyoD, desmin, and the transcription factor myogenin. In a process termed terminal differentiation, myocytes fuse into multinucleated, differentiated myotubes that express the myogenic proteins alpha actinin, desmin, and myosin heavy chain. Afterward, myotubes can further mature into myofibers. Figure adapted from Zammit et al. 33 MyoD, myogenic differentiation 1; Myf5, myogenic regulatory factor 5; Pax7, paired box protein-7.
The essential satellite cell marker is paired box protein-7 (Pax7), which is a transcription factor that is required for normal satellite cell function, and is expressed by both quiescent and proliferating satellite cells. 34 Beyond Pax7, there is significant heterogeneity within the satellite cell population, along with differences in satellite cell marker expression depending on the anatomical location of their niche. 35
The anatomical location of the satellite cell niche also affects satellite cell differentiation potential. For instance, as demonstrated by Collins et al., 36 satellite cells from mouse extensor digitorum longus and soleus muscles contribute significantly more to regeneration in vivo than tibialis anterior (TA) muscle satellite cells. These differences are caused by local stem cell niche and injury microenvironment, 37 and are also affected by differences in cellular composition of the niche that arise from two distinct satellite cell populations that Shultz 38 identified: a reserve population and a responsive population. The responsive satellite cell population comprises roughly 80% of total satellite cells and is the population that readily divides to contribute to de novo myofiber formation. 38 The remaining 20% of satellite cells are part of a reserve population that divides slowly and only in response to an intensive need for muscle regeneration. 38
Satellite Cell Isolation
All satellite cell-based SMTE protocols must create an in vitro approximation of the in vivo stem cell niche and injury environment to induce proper induction, proliferation, terminal differentiation, and tissue formation, and each protocol must begin with proper satellite cell isolation. Satellite cell isolations begin with a skeletal muscle biopsy to obtain skeletal muscle tissue. Skeletal muscle biopsies are painful, and only a small amount of tissue can be taken before a permanent functional deficit is introduced. After biopsy, most satellite cell isolation methods fall into one of two general categories: single fiber explant culture or enzymatic digestion.
Explant culture is a general category of cell isolation, wherein pieces of tissue are plated and cells are allowed to egress out of the tissue and onto the cell culture dish. Bischoff 39 developed the first single fiber explant culture isolation protocol for satellite cell isolation in rats, and it was later improved upon by Rosenblatt et al. 26 In their current state, single fiber explant culture methods (Fig. 2, top) 40 are highly pure, but low yield due to the nature of isolation. 40 In contrast, enzymatic digestion methods (Fig. 2, bottom) 40 are less pure, but yield larger satellite cell populations, and are thus the most common isolation method currently used in SMTE. 40 Enzymatic digestion protocols involve dissecting muscle biopsies, removing any connective tissue by hand, mincing the muscle, and then performing enzymatic digestion, using enzymes like trypsin, pronase, dispase, and collagenase to reduce the tissue into individual cells. 40 Enzymatic digestion is laborious, and efficiency varies widely between laboratories.
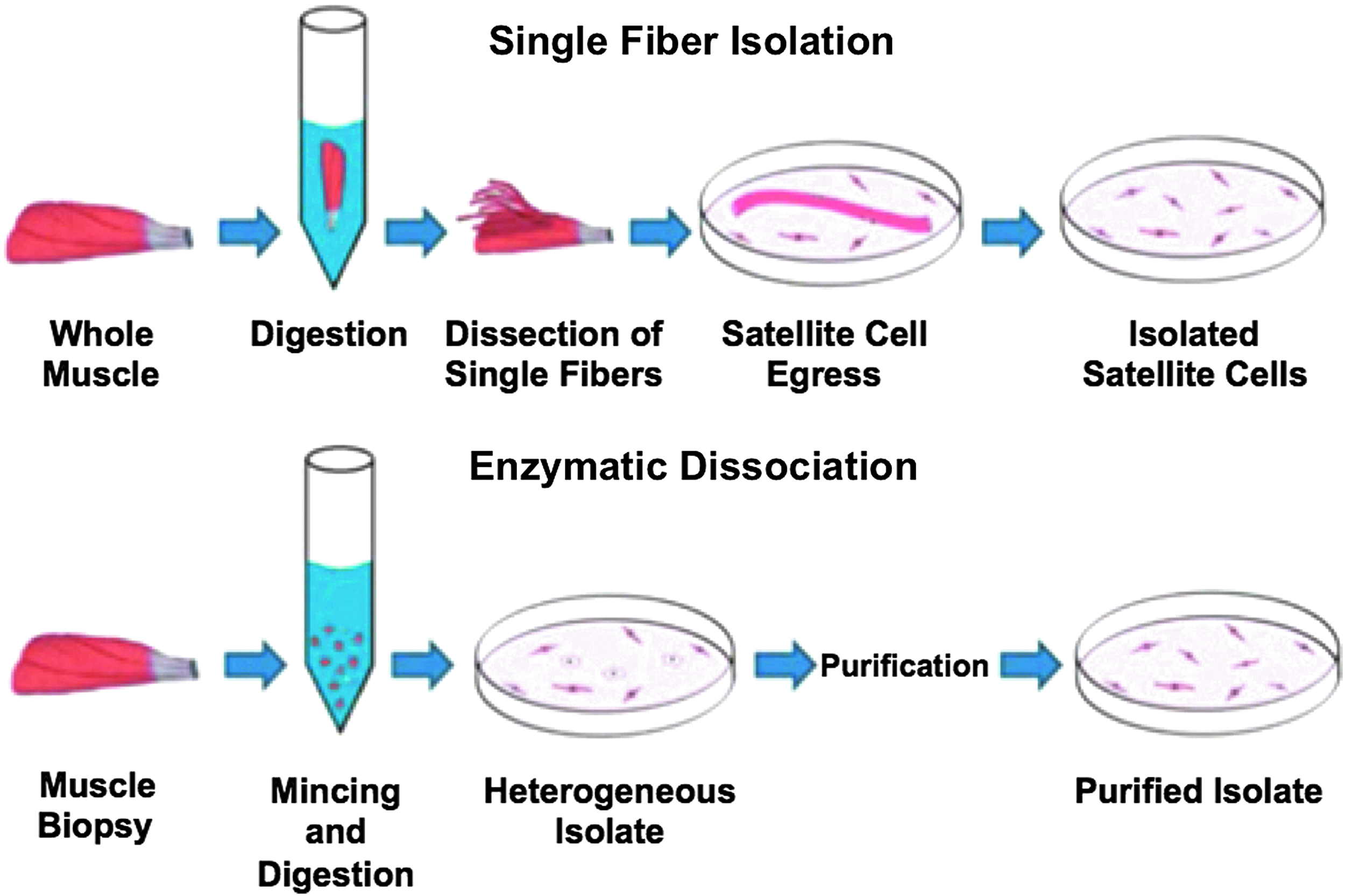
Methods for satellite cell isolation. Single fiber isolation (top) typically involves digesting a whole muscle and dissecting out individual fibers. Fibers have attached satellite cells that are exposed during the digestion process, and the fibers are plated, allowing the satellite cells to egress onto tissue culture plates. Depending on the protocol, fibers may later be removed from the plates to obtain a more pure satellite cell population. Enzymatic dissociation (bottom) involves mincing a muscle biopsy, performing an enzymatic digestion to obtain a heterogeneous population of isolated cells that is typically less pure than the product from single fiber isolation, and then typically purifying the population to obtain an isolated satellite cell product. Figure adapted from Syverud et al. 40
Satellite Cell Purification
After enzymatic digestion, the resulting isolate is typically purified to remove debris and other abundant cell types, including epithelial cells and fibroblasts. Fluorescent-activated cell sorting (FACS) (Fig. 3A) 33 is a purification technique that uses fluorophore-conjugated cell markers to dye cells and applies electrical charges to samples to sort cells by surface marker.41,42 Determining appropriate positive and negative satellite cell markers can be challenging due to satellite cell heterogeneity. Although satellite cells have successfully been purified using FACS,43–46 applying electrical charges to live cells can alter gene expression 47 and reduce proliferative capacity, 48 which could limit their SMTE applications.
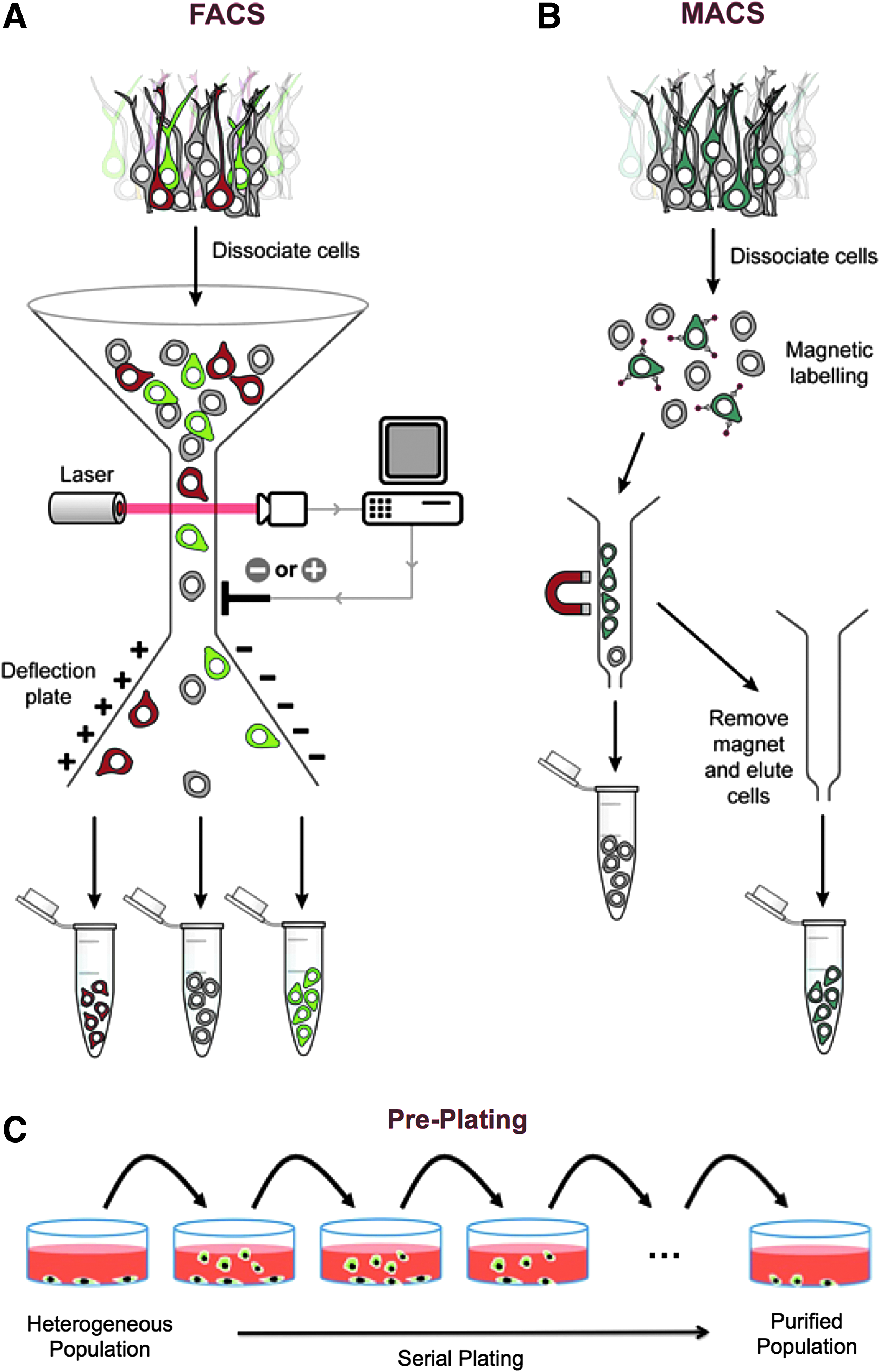
Methods for satellite cell purification. FACS
Magnetic-activated cell sorting (MACS) (Fig. 3B) 33 is an alternative to FACS that works by incubating cells with antibodies for surface markers that are conjugated to magnetic microbeads. A magnetic field is then applied to sort the cells out of the cell suspension. However, appropriate markers must also be determined for MACS, and the tight antibody-antigen association can sometimes cause the magnetic microbeads to be endocytosed by the sorted cells, 40 rendering the beads impossible to remove before implantation. Although MACS and FACS are powerful experimental techniques, given their limitations, it is presently unclear whether they will ever be clinically useful purification techniques for SMTE.
In contrast, the preplate protocol (Fig. 3C) 33 developed by Richler and Yaffe 49 is a simple, fast method to remove fibroblasts and epithelial cells by repeatedly plating and recollecting the cell suspension obtained from enzymatic dissociation over time. Lavasani et al. 50 later developed the modified preplate technique, which coats preplates in collagen I to purify cells by their affinity for collagen I. Preplating is attractive for its simplicity, but it is variable and results in significant loss of satellite cells on preplates. Recently, Syverud et al. 51 published a novel, promising purification method that uses a microfluidic sorting device termed a “Labyrinth” to obtain significantly purer satellite cell populations using label-free, inertial separation; the higher-purity population was then used to grow skeletal muscle constructs able to produce significantly greater tetanic forces than unsorted controls.
Satellite Cell Preservation
To date, satellite cell preservation is not well studied, and the field has yet to comprehensively address the best method for satellite cell cryopreservation or cryopreservation's effects on satellite cell viability, proliferation, marker expression, and differentiation potential. Bian et al. 52 found that satellite cells isolated from Luxi cattle embryo could be successfully cryopreserved after fifteen passages and retained an average viability of 97.90% ± .96% after cryopreservation. Although it is perhaps likely that cryopreservation would be an effective method for satellite cell preservation, more research must be done to assess the most effective satellite cell preservation techniques and any negative effect of the cryopreservation process.
Satellite Cell Proliferation and Differentiation
After isolation and purification, most SMTE protocols proliferate satellite cells by suspending them in a muscle growth medium with high serum and high glucose content. 40 A selection of biomolecules is typically added to enhance proliferation and myogenic potential in culture, including fibroblast growth factor (FGF), hepatocyte growth factor (HGF), platelet-derived growth factor, and insulin-like growth factor 1 (IGF-1).39,53–55 Weist et al. 56 found that when added to muscle medium, transforming growth factor beta (TGF-β1) enhances contractility and ECM organization in scaffoldless, tissue-engineered skeletal muscle tissues. Syverud et al. 57 found that the glucocorticoid dexamethasone (DEX) is a useful addition to muscle media because it enhances the myogenic potential of rat satellite cells in culture, thereby improving the terminal differentiation process by increasing myotube fusion and thickness and enhancing sarcomeric organization.
After being suspended in appropriate media, satellite cells are often seeded onto tissue culture plastic that is coated with an adhesion protein. Laminin, fibronectin, matrigel, and polylysine have all been successfully used in various protocols,53,55,58 and there is no consensus as to which adhesion protein works best. Alternatively, hydrogels, which are gelatinous mixtures of proteins and water, are sometimes used in lieu of protein-coated, stiff glass dishes. Pollot et al. 59 recently compiled a useful review of natural polymeric hydrogels for SMTE. It has been hypothesized that hydrogels can promote growth in a way that is more similar to a stem cell niche by mimicking the stiffness of the ECM niche in vivo. Indeed, Engler et al. 60 found that increasing or decreasing hydrogel stiffness outside of physiologically normal range for skeletal muscle significantly reduces myogenic differentiation in mouse myoblasts, which underscores the importance of engineering an in vitro environment that is as similar to the stem cell niche as possible.
After allowing the cells to proliferate into myoblasts, SMTE protocols induce myogenic differentiation by a muscle differentiation medium (MDM). MDMs typically contain low serum content, because myoblasts interpret serum deprivation in the local environment as a differentiation signal. 61 A wide variety of growth factors, including IGF-1, are also known to promote skeletal muscle differentiation and increase skeletal muscle mass in vivo. 62 Kang et al. 63 found that adding recombinant netrin, a secreted axonal guidance protein that associates with the cell membrane and ECM,64,65 to mouse myoblasts in vitro can induce myotube production and increase myotube size by nuclear factor of activated T cell (NFAT) activation. Arachidonic acid enhances myotube growth in vitro, 66 as do cytokines through the Janus kinase/Signal Transducer and Activator of Transcription (JAK/STAT) pathway. 67
The presence of ECM also encourages myogenic differentiation due to interactions between integrins in the ECM and myoblasts that induce signaling events to promote myogenesis. A study by Stern et al. 68 found that adding ECM extracts from the thigh muscles of adult rats to myoblasts enhances myoblast differentiation in vitro. Mechanical stimulation can also be used to encourage myogenic differentiation. In vivo, mechanical stimulation is required to prevent muscle atrophy, and this has been attributed to the effects that mechanical stimulation has on the mammalian target of rapamycin pathway to regulate gene and protein expression 69 ; it is thought that muscle behaves similarly in vitro.
Because proper alignment of skeletal muscle fibers is crucial for function, tissue engineers must also develop strategies to align skeletal muscle myotubes as they fuse in vitro. Methods to align cells in vitro include topography methods,70,71 surface patterning methods, 72 mechanical stimulation,73–75 and electrical and magnetic stimulation.76,77 Topography methods use surface features to manipulate cell behavior. For instance, growing myoblasts on parallel, micropatterned grooves on tissue culture plates promotes myoblast alignment 78 and myotube fusion in organized, end-to-end configurations. 79 Custom bioreactors, 80 which contain lined, cylindrical microwells, can be used to promote cell alignment and myoblast fusion using topographical principles. The same concepts are applied in surface patterning. Surface patterning is an umbrella term for a variety of techniques that promote cell alignment, including soft lithography. 81 ECM proteins, including collagen, fibronectin, and laminin, can be printed onto cell surfaces by soft lithography to promote proper myotube alignment. 61 As summarized in several review articles,82,83 mechanical stimulation can also be used to promote myotube alignment in vitro by using custom, dynamic bioreactors to add a cyclic strain, which aligns myotubes parallel to the direction of strain. 84 As summarized in a useful review by Ahadian et al., 85 electrical stimulation works by applying an electric field to muscle cells in vitro; because these cells are electroactive, applying an electric field regulates cell behavior and induces muscle tissue alignment parallel to the direction of an applied electric field.86,87 Similarly, magnetic stimulation applies a magnetic field to align myoblasts and elongates them as they differentiate along the axis of the applied magnetic poles 88 ; however, it must be noted that magnetic stimulation is presently a clinically infeasible process, as it requires that cells be electroporated with magnetic nanoparticles.
Satellite Cells in SMTE for VML
Several studies have tested satellite cell-based SMTE in VML animal models.89,90 In 2011, Machingal et al. 13 created and tested the first tissue-engineered muscle repair (TEMR) construct. The researchers created the TEMR by performing an enzymatic dissociation isolation of rat muscle to obtain muscle progenitor cells (MPCs), which is a more general category of Pax7+ cells that includes satellite cells. They cultured the MPCs to obtain rat myoblasts, and seeded the myoblasts onto a porcine bladder acellular matrix (BAM) scaffold to produce the TEMR. Afterward, Machingal et al. used a custom bioreactor for a week after seeding to provide mechanical stretch, thereby promoting differentiation and alignment within the TEMR. Fluorescent microscopy and scanning electron microscopy analysis of the TEMR after bioreactor preconditioning confirmed that the scaffold contained aligned, multinucleated myotubes. After implanting final TEMRs into a murine model for VML injury in the lattissimus dorsi (LD) muscle and allowing the mice to recover for 2 months, they measured isometric tetanic force and observed significant functional recovery of the injured LD in mice treated with TEMRs compared to no repair, although function was not returned to preinjury levels.
This TEMR protocol was further refined by Corona et al. 91 by experimenting with three different varieties of TEMR. The first type of TEMR, termed TEMR-1SP, was seeded with rat-derived MPCs and allowed to proliferate for only a short period of time without bioreactor preconditioning. The second TEMR variety, called TEMR-1SPD, was given a longer cellular proliferation and differentiation period and was conditioned with a bioreactor; this is the TEMR variety that was used in the original Machingal et al.'s 13 study. The third TEMR variety, named TEMR-2SPD, was given a treatment similar to TEMR-1SPD, but additional MDCs were seeded onto the scaffold during bioreactor preconditioning to simulate functional recovery during exercise, wherein MPCs are typically recruited to injured fibers. After implanting these TEMRs into a murine model for VML injury, they measured in vitro isometric tetanic and twitch force. Results showed that TEMR-1SPs and TEMR-1SPDs conferred similar functional recovery, while TEMR-2SPDs induced functional recovery that had twice the magnitude of TEMR-1SPs and TEMR-1SPDs.
Corona et al. 14 later used the same protocol to generate TEMR-1SPDs in a Lewis rat model of VML. When VML rats were treated with TEMRs, results were binary; the rats were either responsive or nonresponsive to the tissue repair. Nonresponsive rats with TEMR implants had no significant improvements in structural or functional recovery compared to untreated rats, while responsive rats had significant structural and functional recovery compared to untreated rats, as assessed by histology and in vivo torque measurements. The researchers noted that responsive and nonresponsive rats had different immune responses in their injury site, which could have contributed to the observed variability.
VanDusen et al. 16 published a method for engineering scaffoldless, three-dimensional, functional skeletal muscle units (SMUs) and tested them in a rat VML model. The researchers used enzymatic digestion of rat soleus muscles to isolate satellite cells, cultivated them using a muscle growth medium, and differentiated them in vitro using a medium with low serum content. After the muscle monolayers rolled up into a 3D skeletal muscle tissue due to passive tension, they were pinned to the plate and engineered bone/tendon anchors were added to either end, generating the final SMU. In this specific study, the researchers measured isometric tetanic force of their SMUs to ensure functionality. The SMUs were then implanted into a VML site in the rat's TA muscle. After 28 days of recovery, they observed a significant increase in isometric tetanic force for VML muscles treated with SMUs compared to rats with VML receiving no treatment, and a force per unit mass for SMU-treated muscles that was equivalent to native muscle.
Satellite Cell Limitations
Satellite cell-based SMTE is undoubtedly a promising field with demonstrated potential as a VML treatment, but using satellite cells has significant limitations. A skeletal muscle biopsy is painful and invasive, and only a small biopsy would be clinically feasible. Enzymatic dissociation is the only isolation method that is able to achieve sufficient satellite cell yields, yet it is a laborious process and produces impure populations of satellite cells. With respect to purification, FACS, MACS, and preplating have significant practical limitations, while microfluidic inertial separation is a promising, yet nascent method. Satellite cell preservation is not well studied, and satellite cell self-renewal is not unlimited, and satellite cell differentiation potential declines rapidly in vivo. To address these issues, it is important to consider alternative stem cell sources, including ADSCs, BM-MSCs, umbilical cord-derived mesenchymal stem cells (UC-MSCs), PVSCs, iPSCs, and embryonic stem cells (ESCs).
Adipose-Derived Stem Cells
ADSCs are an abundant, adult, multipotent, mesenchymal stem cell (MSC) variety with myogenic potential. MSCs are a general category of nonhematopoietic stem cells with specific cluster of differentiation (CD) protein surface markers that reside in many tissue types, including adipose tissue. MSCs have the capacity to differentiate into tissues of mesodermal lineage, including cartilage, bone, adipose tissue, and skeletal muscle.92–94 A useful ADSC tissue engineering review by Zuk 95 contains a comprehensive discussion of human ADSC surface markers.
ADSCs are attractive to tissue engineers not just for their abundance, but because cultured ADSCs express very low levels of major histocompatibility complex class I (MHC I) and do not express major histocompatibility complex class II (MHC II). 96 Thus, ADSCs from allogeneic or xenogeneic donors might be able to evade the patient's immune system after implantation, potentially eliminating the need for lifelong immunosuppression in patients. 96 It is important to note that the low levels of MHC I and MHC II observed in ADSCs are present before differentiation; there has not yet been a study that investigates MHC I and MHC II levels in ADSC-derived cells after myogenic differentiation.
ADSC Isolation and Purification
ADSC isolation protocols begin with an adipose tissue biopsy. 97 As noted in the Forcales 98 review of ADSCs for muscle regenerative therapies, adipose tissue is a highly abundant, cosmetically favorable tissue type to biopsy in most individuals, and a simple liposuction procedure to collect lipoaspirate typically suffices. Moreover, ADSC isolations are generally efficient, with an average of 404,000 ± 206,000 cells isolated per milliliter of lipoaspirate. 99 Importantly, ADSC yield does not depend on the donor site, 100 so fat can be taken from the most convenient location possible. Thus, ADSCs offer significant advantages over satellite cells with respect to biopsy.
After biopsy, ADSC isolations typically use enzymatic digestion of the biopsied adipose tissue or lipoaspirate with collagenase type 1A. 98 After digestion, the mixture is centrifuged to obtain the stromal vascular fraction (SVF) pellet, which contains many cell types, including preadipocytes, pericytes, fibroblasts, red blood cells, and smooth muscle cells. 98 The SVF is plated in ADSC media, which can vary by protocol, but typically contain Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS). The cells that adhere to the plate are considered to be ADSCs. 98 FACS and MACS can be used to purify ADSC populations, but since ADSCs lack a universal marker, these processes are complicated and imperfect. Cryopreservation is a suitable method for ADSC preservation, as summarized in a review by Miyagi-Shiohira et al. 101
Several specific ADSC isolation methods may offer advantages over satellite cell enzymatic dissociation. Like satellite cells, explant culture can be used for ADSC isolations, and Priya et al. 102 developed an explant culture method that achieved equivalent to superior human ADSC yields when compared to enzymatic dissociation. Because explant culture tends to result in purer cell populations than enzymatic dissociation, 40 using explant culture could eliminate the need for further purification. There is also a novel protocol developed by Francis et al. 103 to isolate human ADSCs in just 30 min. Future SMTE studies could consider using one of these protocols to obtain a starting ADSC population.
ADSC Induction
There are several different methods for inducing ADSCs into myoblasts, including using appropriate myogenic media with myogenic biochemical factors, coculturing ADSCs with myoblasts, adding biophysical stimuli, and genetically manipulating the ADSCs. Interestingly, ADSC myogenesis is similar to satellite cell myogenesis in terms of when Pax7, Myf5, and MyoD are expressed. 104
Di Rocco et al. 105 observed that a small proportion of mouse ADSCs are able to sporadically convert into a myogenic lineage, which suggests that certain ADSCs do have inherent myogenic potential. They also experimented with 50–50 cocultures of myoblasts and ADSCs, and found that ADSCs will fuse into myotubes when cultured with myoblasts, but only when the ADSCs and myoblasts are in direct contact. Afterward, they tested ADSC regenerative capacity in vivo and found that when uncultured ADSCs were injected into a mouse with ischemic skeletal muscle, they were able to fuse with muscle fibers to contribute to structural recovery in vivo. Thus, simply placing ADSCs into the site of injury can induce some degree of induction and differentiation toward a myogenic lineage, although it is unknown whether this would contribute to meaningful functional recovery.
Biophysical stimuli can also induce ADSCs to a myogenic lineage, and Huri et al. 106 found that adding a cyclic strain to human ADSCs (hADSCs) treated with muscle induction media not only encourages proper cell alignment but also enhances myogenic differentiation into myotubes. In fact, after adding the cyclic strain, the researchers observed a 4.5-fold increase in nuclei per cell, which suggests enhanced myogenesis. They also observed significantly higher myogenic protein expression compared to cultures not given the cyclic strain.
Altering gene expression has also been explored as a method to differentiate ADSCs, and Goudenege et al. 107 developed an ADSC differentiation protocol that uses a mouse MyoD lentiviral vector to forcibly induce MyoD expression, thereby causing myogenic induction. MyoD is an essential myogenic regulatory factor that induces an open chromatin structure at myogenic genes. 108 The researchers found that using their protocol to induce MyoD expression in hADSCs was a fairly efficient way to achieve myogenic induction, with 30% of treated hADSCs expressing myogenic markers, compared to just 2% of untreated hADSCs. Notably, Goudenege et al. also found that when injected into TA cryoinjuries in a murine model, the transfected hADSCs were capable of fusing with regenerating fibers in the injury site to contribute to structural and functional recovery in vivo. Sung et al. 109 tested a safer method for induction by engineering and applying MyoD protein to hADSC cultures to induce myogenesis in vitro. From a clinical perspective, applying MyoD protein is a far more practical and safe method than altering MyoD gene expression, and would be a more feasible method to try in SMTE.
ADSCs in SMTE for VML
In 2016, Kesireddy 110 published the first comparative study that used rat ADSCs to create a TEMR construct to treat VML injury in a murine model and compared it to MPC TEMRs. To make a TEMR, he isolated ADSCs and MPCs from donor rats, cultivated them, and seeded either ADSCs or MPCs into BAM scaffolds. Afterward, he implanted them into a murine VML injury model in the LD muscle. Histological analysis of the injury site 2 months postrepair revealed that ADSC and MPC TEMRs displayed similar regeneration potential, and both showed signs of neotissue formation, which is characterized by scaffold remodeling, fiber formation, and cell fusion in the VML site. 110
Kesireddy also did qualitative observation of lentivirus-Cherry-labeled donor cells to determine the origin of the structural recovery that was observed, and discovered that ADSCs contributed fairly little to new myofiber formation compared to donor MPCs. However, ADSCs contributed greatly to vascularization in the injury site, and histological images revealed greater vascularization after being treated with ADSCs than MPCs, 110 which is a positive sign of healing. Unfortunately, this experiment did not conduct force testing to assess functional recovery, but the results nevertheless suggest that ADSCs could be useful for treating VML.
Bone Marrow-Derived MSCs
BM-MSCs are another notable MSC variety with myogenic potential. BM-MSCs are a multipotent, adult stem cell population that resides in the bone marrow (BM) on the abluminal side of vessels and can differentiate into mesodermal tissues like skeletal muscle, bone, and cartilage. 111 It is important to note that BM-MSCs are distinct from BM hematopoietic stem cells and do not directly participate in hematopoiesis; rather, BM-MSCs play an immunomodulatory role, and provide microenvironment support for hematopoietic stem cells in vivo. 112
BM-MSCs are a heterogeneous stem cell population, and individual subsets have significant differences in markers, as detailed in a review by Boxall and Jones. 113 Because of the similarities between BM-MSCs and ADSCs, which are both MSCs with myogenic potential that are found in vessel walls in their respective tissues of origin, Bayati et al. 114 compared rat BM-MSCs and ADSCs and discovered that the two cell populations had differential surface marker expression, but did not significantly differ in terms of proliferative capacity or myogenic potential. Also, like ADSCs, BM-MSCs have poor ability to stimulate an in vitro allogeneic T cell response due to absent MHC II and low MHC I expression, 115 which makes them well suited to transplantation.
BM-MSC Isolation and Purification
BM-MSC isolation begins with a BM biopsy, which is a painful and invasive process; however, it is still advantaged over satellite cell biopsies in that more tissue can be taken without introducing a functional deficit. After a biopsy, typical BM-MSC isolations use enzymatic digestion, just like ADSC and satellite cell protocols. 116 BM isolates are filtered and washed, and enzymes are used to digest the BM isolate to retrieve the BM-MSCs. Similar to ADSCs, explant culture has also more recently been proposed as an efficient method for BM-MSC isolation. 117 Generally speaking, BM-MSC isolation from BM samples is a fairly low-yield process, because there are less than. 01% BM-MSCs present in total BM cells. 92 FACS and MACS can be used to purify BM-MSC populations if that is deemed necessary, and cryopreservation methods for BM-MSCs have been developed.118,119
BM-MSC Induction
BM-MSC induction methods are similar to what was already discussed for ADSCs and satellite cells; most involve muscle induction media, coculture, biophysical stimuli, or some combination of these methods. Treatment with 5-azacytidine is a common method for induction of BM-MSCs to a myogenic lineage in vitro, although this is not unique to BM-MSCs, and can also be used for induction of ADSCs. 120 Although using 5-azacytidine for myogenic induction is a useful research technique, it is known that 5-azacytidine is a carcinogen, 121 and is therefore impractical for SMTE. A study by Stern-Straeter et al. 122 evaluated the efficacy of six different muscle induction medium formulations for hADSCs and human BM-MSCs (hBM-MSCs) to offer guidance on which induction media work best. Beier et al. 123 perfected a rat BM-MSC/myoblast coculture method by using basic FGF (bFGF) and DEX to maximize myogenic potential. Egusa et al. 124 were the first to discover that adding a cyclic strain to mouse BM-MSCs accelerates myogenic induction and promotes proper fiber alignment during the terminal differentiation process.
An important study by Meligy et al. 125 compared proliferation and myogenic differentiation capacity between ADSCs, BM-MSCs, and skeletal muscle tissue MSCs (MC-MSCs), which are found in skeletal muscle, but have a MSC phenotype and are distinct from satellite cells. After isolating and cultivating rat MSCs from their respective tissues of origin, they treated the cells with 5-azacytidine to promote induction to a myogenic lineage. ADSCs had the highest observed proliferation rate and BM-MSCs had the lowest proliferation rate, 125 although all were fairly high. The researchers measured differentiation potential by myogenin expression and observed that BM-MSCs had higher myogenic differentiation capacity than ADSCs, although both were inferior to MC-MSCs. 125 From this, Meligy et al. concluded that ADSCs were the most easily accessible and highly proliferative MSCs. Although MC-MSCs had the highest observed myogenic differentiation potential, the researchers noted that MC-MSCs isolated from a skeletal muscle biopsy are no more accessible than satellite cells, suggesting that ADSCs and BM-MSCs could still be a viable starting source for SMTE, regardless of their lower myogenic differentiation potential.
BM-MSCs in SMTE
There has never been a published study that used BM-MSCs to engineer skeletal muscle tissue specifically for VML. However, there are several studies that have used BM-MSC injections to treat skeletal muscle injuries, and they offer clues as to how BM-MSCs would behave in SMTE specifically for VML. Natsu et al. 126 isolated BM-MSCs from green fluorescent protein (GFP) rats and injected them into lacerated TA muscles of Sprague-Dawley (SD) rats after implanting a fibrin scaffold. One month after transplantation, the rats treated with BM-MSCs showed significant functional recovery that was almost that of preinjury levels. Interestingly, histological analysis did not indicate that the GFP BM-MSCs actually differentiated or fused into myofibers in the SD rats, which suggests that BM-MSCs may play an indirect role in healing, possibly by promoting a healing immune environment in the injury site.
Merritt et al. 127 repaired skeletal muscle injury in a rat model by implanting an ECM scaffold into injured lateral gastrocnemius muscles and injecting BM-MSCs into a group of rats treated with the ECM scaffold a week after surgery. After 42 days, the injured muscles treated with ECM and BM-MSCs displayed significant structural recovery 127 and partial functional recovery that were superior to muscles treated with ECM alone. 127 BM-MSC-injected muscles also had more blood vessels and regenerating fibers. From this, the researchers concluded that BM-MSCs could be a viable therapy for skeletal muscle injury in the future. Andrade et al. 128 found that BM-MSCs administered through intramuscular injection into a rat model of successive injury significantly increased maximal skeletal muscle contraction, muscle fiber cross-sectional area, and the number of muscle fibers in the injury site 28 days after injection.
Helal et al. 129 found that BM-MSCs modulate the local immune system environment in the site of VML to encourage recovery. Specifically, administration of BM-MSCs through intramuscular injection into a VML injury in a rat was shown to improve structural and functional recovery by suppressing inflammatory cytokines, promoting anti-inflammatory cytokines, and directly contributing to myogenesis and angiogenesis. BM-MSCs also downregulate TGF-β in the injury site, which prevents fibrosis. As this study demonstrates, BM-MSCs show promise for SMTE applications not only because of their myogenic potential but also because of the role they play in immunomodulation to promote proper healing.
Umbilical Cord-Derived MSCs
UC-MSCs are derived from compartments of the umbilical cord, with UC-MSCs isolated from the cord-placenta junction region showing particularly high differentiation potential. 130 UC-MSCs are more similar to ESCs than adult stem cells, and they are capable of faster self-renewal than adult MSCs. 131 Collecting UC-MSCs is also a painless process, and UC-MSCs lack MHC II, which is useful for transplantation. 132 UC-MSCs can be isolated by enzymatic digestion 133 or explant culture methods.130,134 A good manufacturing practice (GMP) protocol for clinical-grade human UC-MSC isolation was recently published, 135 and a useful summary of UC-MSC markers, characteristics, and isolation methods is included in a review by Nagamura and He 136
Several myogenic induction protocols exist for UC-MSCs. Gang et al. 137 demonstrated that UC-MSCs are capable of being induced into a myogenic lineage by cultivating them in myogenic media that contained 5% horse serum, 0.1 μM DEX, and 50 μM hydrocortisone for 6 weeks. Kocaefe et al. 132 transfected human UC-MSCs with MyoD to force induction into a myoblastic lineage. They found that the resulting cells were capable of undergoing terminal differentiation by fusing with rat myoblasts to form myotubes, which suggests that UC-MSCs could be a viable starting source for SMTE in the future.
To date, UC-MSCs are best studied in the context of muscular dystrophy therapies. A study by Zucconi et al. 138 found that undifferentiated UC-MSCs injected into mouse and dog muscular dystrophy models did not differentiate into skeletal muscle in vivo, yet dystrophic animals injected with them displayed significant functional recovery compared to untreated controls. In contrast, a different study by Nunes et al. 139 was able to observe some degree of structural recovery, with a small number of mouse UC-MSCs able to differentiate into dystrophin-expressing myotubes in vivo when transplanted into mdx mice. While the exact reason for these conflicting results is not clear, it was shown in a study by Koponen et al. 140 that UC-MSCs are able to promote skeletal muscle regeneration in lacerated mouse skeletal muscle not by directly fusing with myofibers, but by promoting a healing cytokine environment. While it appears that UC-MSCs could be a useful source for clinical skeletal muscle treatments in the future, more research must be done to understand the mechanism and extent of healing in traumatic injury applications.
Perivascular Stem Cells
PVSCs are yet another MSC population that can be subdivided into pericytes and adventitial cells. 141 Adventitial cells are found in the tunica adventitia (Fig. 4, right), 142 which is the outermost layer of arteries and veins, 143 and they are defined as positive for CD34 and negative for CD31, CD146, and CD45. 143 Although adventitial cells share developmental features and general MSC properties with pericytes, they are a phenotypically and genotypically distinct population. 143 Their myogenic potential has not yet been studied, whereas pericytes have been studied in greater detail, discussed below.
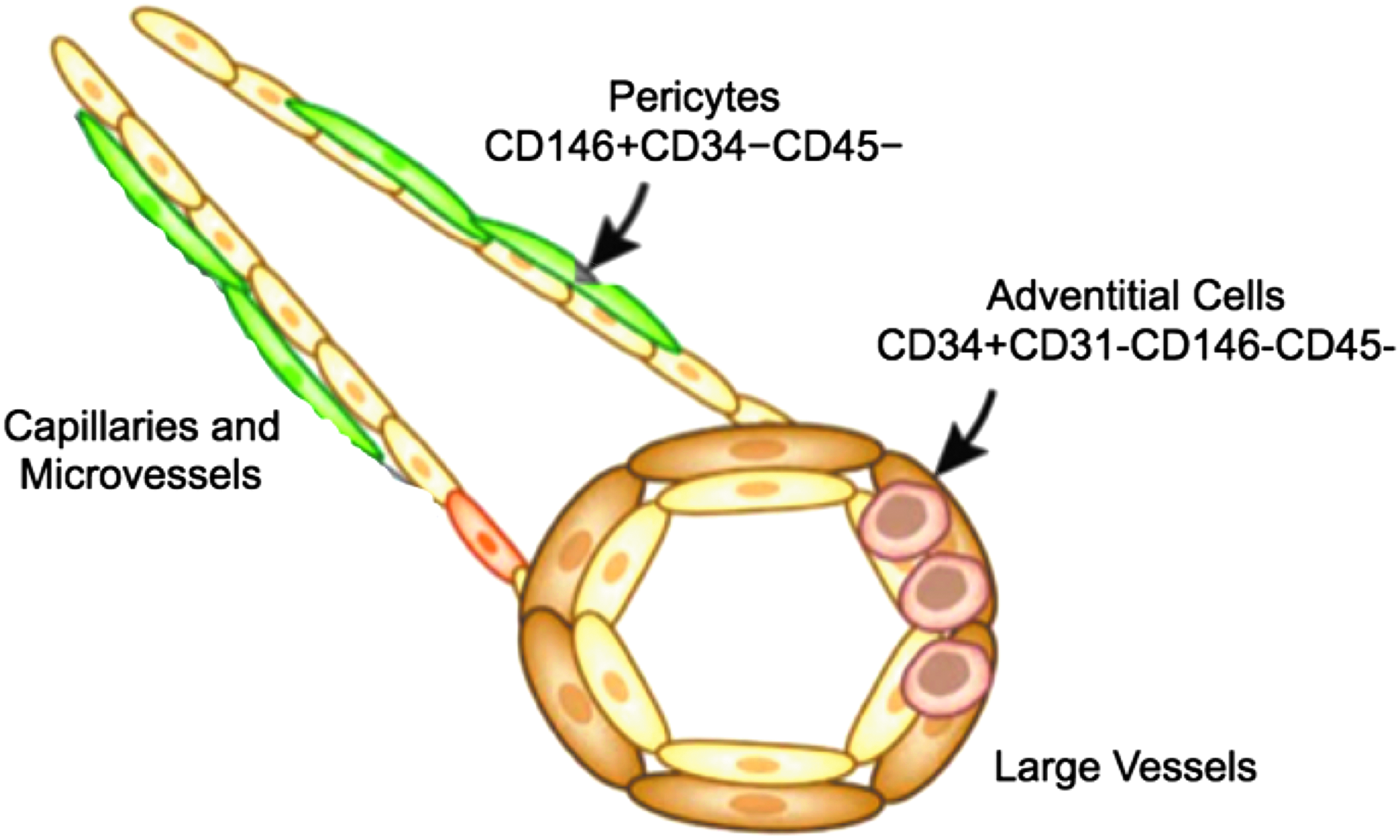
Visual summary of the relative locations and basic properties of the two varieties of perivascular stem cells: pericytes and adventitial cells. Pericytes are located in capillary and microvessel walls and are defined as positive for CD146 and negative for CD34 and CD45. Adventitial cells are located in the walls of large vessels (veins and arteries) and are positive for CD34 and negative for CD31, CD145, and CD45. Figure adapted from Bautch. 142
Pericytes
Pericytes are a stem cell population found in capillary and microvessel walls 142 underneath the basal lamina (Fig. 4, left) 142 in a variety of tissues, including skeletal muscle 144 and adipose tissue. 141 Pericytes isolated from skeletal muscle, white adipose tissue, pancreas, and placenta have myogenic potential, 145 and adipose tissue in particular is an attractive biopsy site to target. Dellavalle et al. 146 discovered that mouse pericytes actually participate in normal postnatal skeletal muscle development by differentiating into muscle fibers and infiltrating the satellite cell niche to generate satellite cells. Thus, pericyte myogenesis is a natural process that occurs during development and increases in response to acute injury. Pericytes are defined as positive for CD146 and negative for CD34 and CD45. 141 Most pericyte isolations involve enzymatic digestion, although explant culture has also been used, 147 and Bryan and D'Amore 148 wrote a useful review of methods to isolate and culture pericytes. After isolation, pericytes can be purified using FACS or MACS.
An important study by Dellavalle et al. 147 first discovered the promise of pericytes for skeletal muscle therapies. Dellavalle et al. began by isolating pericytes from human skeletal muscle biopsies with and without Duchenne Muscular Dystrophy (DMD), and then expanded them out to 20 population doublings, which allowed them to obtain at least 1 × 109 cells from a single biopsy, an amount they considered sufficient for clinical DMD therapy. After culturing the pericytes, they promoted induction to a myogenic lineage and differentiation using both a myogenic differentiation medium and coculture with myoblasts, and observed that pericytes formed differentiated myotubes spontaneously with high efficiency using both differentiation methods. 147 In fact, the differentiation potential they observed for pericytes was strikingly high, and up to an order of magnitude higher than most other myogenic stem cells they compared them to, excluding satellite cells. Dellavalle et al. also injected cultured human pericytes into a severe combined immune-deficient–X-linked, mouse muscular dystrophy (scid–mdx) mouse model, and found that pericytes could successfully engraft with dystrophic muscle fibers to contribute to structural recovery in vivo. The qualities of pericytes Dellavalle et al. observed—adequate proliferative capacity, high myogenic potential, and ability to contribute to fiber regeneration in vivo—coupled with the fact that pericytes can be isolated from other, more convenient tissue types to biopsy, suggests that pericytes could be a highly useful starting cell for SMTE. Moreover, pericytes do not express MHC II or essential T cell activating costimulatory molecules like CD80 and CD86, which suggests that pericytes are a promising candidate for allotransplantation. 149
To date, pericytes have never been used in an SMTE study specifically for VML; however, a study by Fuoco et al. 150 offers guidance to tissue engineers looking to develop a pericyte SMTE protocol. Fuoco et al. isolated skeletal muscle-derived pericytes (MPs) from young and old pigs using enzymatic digestion with collagenase II and induced myogenesis through media with low serum content. Myogenic potential of pericytes declines with age, and the researchers found that old MPs had reduced ability to form myotube and capillary-like structures in vitro on their own. However, when the old MPs were cultured on mixtures of polyethylene glycol, fibrinogen, and water called PF hydrogels, they displayed enhanced myogenic potential in vitro and in vivo. 150 PF hydrogels are believed to rejuvenate myogenic potential because they better mimic the stem cell niche in vivo, and they could be a useful scaffold material to try in a pericyte-based SMTE protocol in the future.
Induced Pluripotent Stem Cells
iPSCs are produced in the laboratory by culturing adult cells and forcing them to upregulate transcription factors that induce pluripotency. Because they are pluripotent, they are capable of unlimited self-renewal in culture, which is a major advantage over satellite cells. Nishikawa et al. 151 wrote a useful review of the promises and present limitations of iPSCs.
Making iPSCs
Technically, any tissue type could be used to make iPSCs; however, in practice, making iPSCs for use in skeletal muscle therapies (Fig. 5) 152 typically begins with a simple skin biopsy to isolate epithelial cells and fibroblasts. 152 These cell types are chosen for their abundance and convenience; thus, iPSC biopsy offers a clear advantage over satellite cell biopsy. After biopsy, pluripotency must be induced in the cells. Takahashi and Yamanaka's 153 pioneering article determined four essential transcription factors, termed Yamanaka factors, which must be transduced into starting cells using a viral vector to reprogram the cells into iPSCs: KLF4, c-MYC, OCT4, and SOX2. Although fibroblasts and epithelial cells are abundant, iPSC generation is not efficient, with the percentage of adult cells successfully reprogramming to pluripotency often somewhere in the range of 0.1–3%.153,154 This is partially attributable to the fact that cells retain epigenetic markers from their tissue of origin that make them resistant to reprogramming.155,156
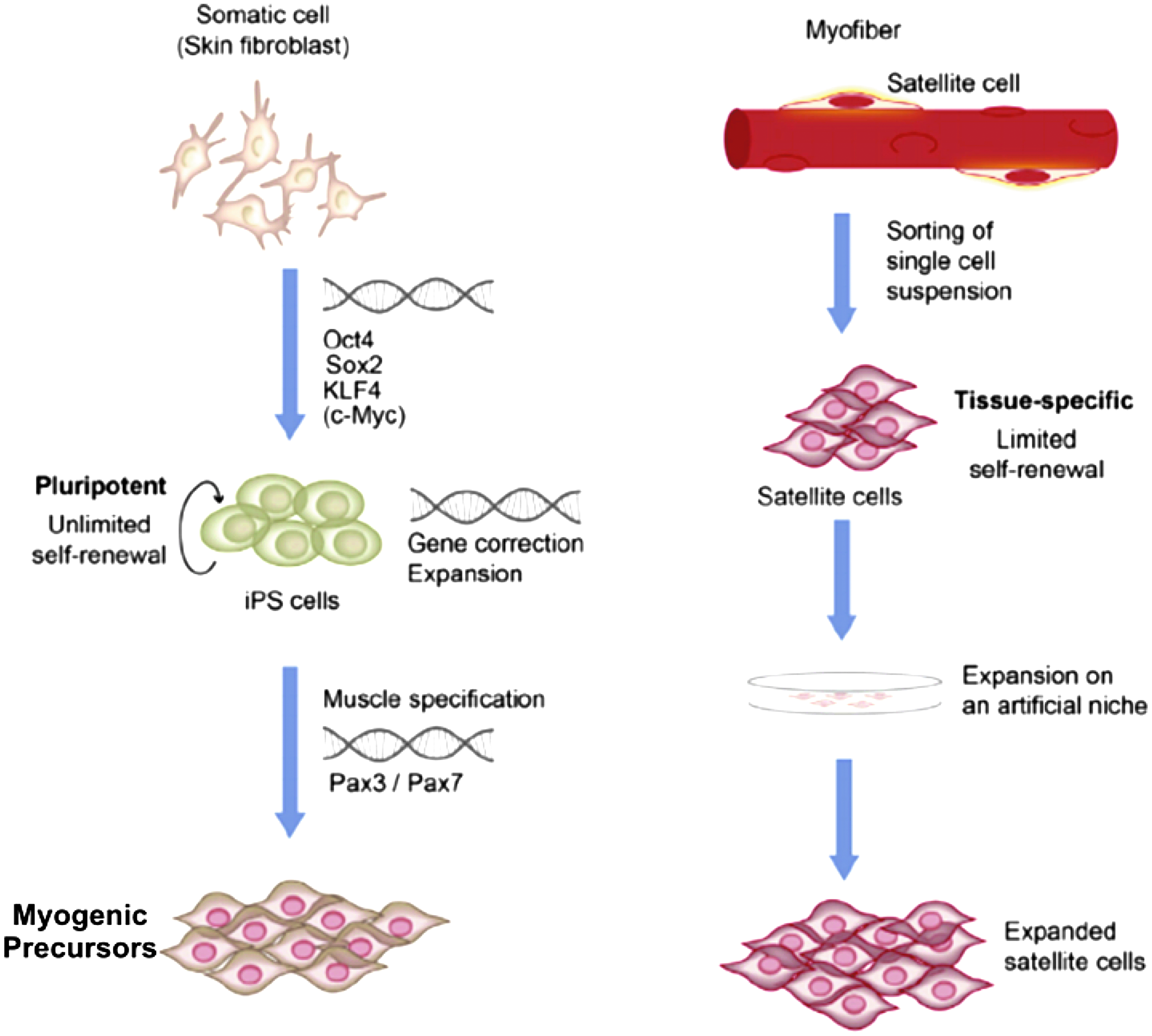
Visual summary of iPSC generation for muscle therapy applications with a comparison to satellite cell methods. Making iPSCs starts with taking a skin biopsy to isolate fibroblasts. The four Yamanaka transcription factors (Oct4, Sox2, KLF4, and c-Myc) are added to induce pluripotency. iPSCs can become muscle precursor cells by inducing muscle-specific genes like Pax7 and Pax3. Then, the myogenic precursors can be transplanted into a diseased patient. Unlike iPSCs, satellite cells are isolated from skeletal muscle biopsy and must be expanded in culture, but do not have the capacity for unlimited self-renewal. After expansion, satellite cells can be transplanted back into a patient. Figure is modified from Ref. 152. iPSC, induced pluripotent stem cell.
In the years since Yamanaka's pioneering study, other scientists have worked to develop more efficient iPSC protocols. Warren et al. 157 optimized the original iPSC method using synthetic modified mRNA to transfect cells, causing them to be reprogrammed to pluripotency at a significantly higher efficiency, while Yulin et al. 158 and Kunisato et al. 159 have developed more efficient iPSC generation protocols that use BM-MSCs as a starting source; given that BM-MSCs have inherent myogenic differentiation potential, it is presently unclear whether these would have any relevance to SMTE. Malik and Rao 160 compiled a more comprehensive review of human iPSC generation methods. Regardless, the development of more efficient iPSC protocols suggests that iPSCs could eventually be a more practical therapeutic option in SMTE applications. After iPSC generation, cryopreservation is an efficient method for iPSC storage. 161
iPSC Induction
iPSCs must be properly induced into a myogenic lineage before implantation. A variety of different protocols have been developed to induce myogenesis in iPSCs, and most of these methods involve inducing myogenic gene expression. Shoji et al. 162 developed a reliable myogenic induction protocol for iPSCs that works by delivering a tetracycline-inducible MyoD piggyBac (PB) vector to human iPSCs, achieving robust differentiation efficiency that is as high as 70–90% in vitro. Chal et al. 163 developed a serum-free method for inducing iPSCs into muscle fibers and satellite-like cells by mimicking embryonic signaling events to induce myogenesis in vitro. Maffioletti et al. 164 demonstrated a complex, thorough method for myogenic induction and differentiation of human ESCs (hESC) and iPSCs, which used a four-stage system with different culture densities, media combinations, and protein coatings on the cell culture dish. First, they drove ESCs and iPSCs to a lineage that was similar to mesoangioblasts, and then they induced MyoD1 expression through a lentivirus vector with MyoD1 cDNA fused to an estrogen receptor, thereby driving the cells to a myogenic lineage. Darabi et al. 165 developed a protocol that makes iPSCs from fibroblasts derived from inducible Pax7 mice and then induces Pax7 expression to induce the iPSCs into myogenic progenitors.
It is important to note that iPSCs are ultimately only useful in SMTE if they are clinically safe. Reprogramming iPSCs to pluripotency and then to a myogenic lineage can induce genetic instability and abnormalities that cause cancers, and iPSCs are known to be tumorigenic.166,167 More emphasis should be placed on developing iPSC myogenic induction and differentiation methods that do not require genetic modification, and researchers have already begun this work. Hosoyama et al. 168 derived myogenic progenitors from human iPSCs by using a medium with fibroblast growth factor-2 (FGF-2) and epidermal growth factor to form free-floating spherical cultures that contained myogenic progenitors after 6 weeks. After culturing these progenitors in an MDM for 2 weeks, the researchers observed multinucleated myotubes. Xu et al. 169 identified three specific myogenic factors: a glycogen synthase kinase 3β (GSK3β) inhibitor called 6-bromo-indirubin-3′-oxime (BIO), FGF, and forskolin. By treating human iPSCs with a combination of these three factors, they were able to produce myogenic progenitors that could engraft into mdx mouse dystrophic muscle fibers in vivo. Any of these methods could be tried on iPSCs in an SMTE study.
iPSCs in Skeletal Muscle Therapies
To date, iPSCs have never been used in an SMTE study to treat VML, but iPSC injection therapy for DMD is better studied and offers clues as to how iPSCs might behave in SMTE. Darabi and Perlingeiro 170 found that human iPSC-derived myogenic progenitors restore dystrophin and improve contractility upon transplantation in dystrophic mice, and Xu et al. 169 found that human iPSC-derived myogenic precursors can engraft with mdx mouse muscle fibers in vivo. Quattrocelli et al. 171 found that iPSCs that have been induced to a mesodermal lineage are able to contribute to functional recovery of cardiac and skeletal muscle in an mdx mouse and dystrophic canines. Goudenege et al. 172 found that myoblasts derived from human iPSCs are able to efficiently fuse with existing muscle fibers following transplantation. These positive results suggest that iPSCs could integrate with existing tissue in an injury site in SMTE for VML.
Embryonic Stem Cells
ESCs are pluripotent stem cells isolated from the inner cell mass of a blastocyst. Beyond the ethical challenges that using ESCs pose, 173 ESCs are difficult and inefficient to isolate, and the isolation process requires high specialization and training. 174 ESCs are also tumorigenic. 175 Partially because of these factors, ESCs have not been used in muscle therapy studies to any meaningful degree.
Discussion
Although satellite cells remain the best-studied cell type in SMTE for VML, they can be difficult to isolate, purify, and biopsy. Alternative stem cells have characteristics that may provide solutions to these problems, although much is presently unknown about how alternative stem cell varieties will perform in SMTE studies.
Judging on the biopsy alone (Table 1), ADSCs and pericytes are highly attractive starting sources that offer advantages over satellite cells, because both can be isolated from adipose tissue. Adipose tissue is the most promising location to target for pericyte isolation not only for convenience but also because pericytes make up 15% of the SVF that is obtained during normal ADSC isolation, 176 but constitute less than 0.5% of cells isolated from bone marrow. Adventitial cells make up 20% of the SVF, 143 but their myogenic potential is currently unknown, and should be explored in future studies. BM-MSCs are superior to satellite cells in that a larger biopsy can be taken without introducing a functional deficit. Yet, a BM biopsy is inferior to an adipose tissue biopsy, and BM-MSCs make up less than .01% of total BM cells biopsied. 92 UC-MSCs can be easily obtained after birth, but unless allotransplanted UC-MSCs become viable, UC-MSCs will not be an option for many current VML patients. iPSCs can be obtained from a convenient skin biopsy, but iPSC reprogramming is not highly efficient.
ADSC, adipose-derived stem cell; BM-MSC, bone marrow-derived mesenchymal stem cell; UC-MSC, umbilical cord-derived mesenchymal stem cell; ESC, embryonic stem cell.
With respect to isolation (Table 1), ADSCs and BM-MSCs may offer advantages over satellite cells in SMTE. ADSC isolation in particular is highly efficient, 98 and in recent years, explant culture has been developed as an efficient method for ADSC and BM-MSC isolation that result in purer cell populations than enzymatic dissociation. Explant culture may also decrease the cost, processing time, and labor required for cell isolation. 177 The Francis et al. 103 protocol to isolate ADSCs in 30 min is a unique and promising method that also does not use enzymatic dissociation, and it is far faster and simpler than satellite cell isolation. Pericytes can also use explant culture; although yields are fairly low, pericytes can be expanded for at least 20 population doublings in culture. 147 Notably, a clinically safe GMP protocol for UC-MSC isolation has also been developed. 135
No alternative stem cell is inherently easier to purify than satellite cells, and FACS and MACS remain the most common purification techniques (Table 2) across cell types. Alternative stem cell varieties have significant marker heterogeneity, so FACS and MACS can be particularly challenging, in addition to the inherent limitations of these methods. However, it must also be noted that purification is more commonly required when enzymatic dissociation is performed, because the resulting isolate is less pure. If explant culture methods to isolate ADSCs, BM-MSCs, or pericytes could be used effectively in SMTE, it is perhaps less likely that purification would be required, which is an important potential advantage that these cell types offer. Notably, a microfluidic inertial sorting method was recently developed to obtain purer satellite cell populations 51 ; this method is advantaged in that it is label free, but it has yet to be widely experimented with.
iPSC, induced pluripotent stem cell; MC-MSC, muscle tissue mesenchymal stem cell.
In terms of proliferative capacity and myogenic differentiation potential (Table 2), it is difficult to say which alternative stem cell is best, although pericytes have particular promise. As Dellavalle et al. 147 noted, pericytes have higher innate myogenic potential than most other stem cells, and their proliferative capacity is sufficient for clinical therapy. Pierantozzi et al. 178 found that pericytes have similar proliferative capacity to ADSCs and are able to differentiate toward myocytes more efficiently than ADSCs. Pericytes' proliferative capacity and myogenic potential compared to UC-MSCs and BM-MSCs should be determined in future studies. UC-MSCs also have great promise. It is known that UC-MSCs have higher proliferative capacity than adult MSCs like ADSCs and BM-MSCs, 179 but the myogenic differentiation ability of UC-MSCs compared to ADSCs and BM-MSCs is not well understood. Between BM-MSCs and ADSCs, it is difficult to determine which cell type is superior. Meligy et al. 125 found that ADSCs were more highly proliferative than BM-MSCs, but BM-MSCs had a higher myogenin expression than ADSCs, which correlates to myogenic differentiation potential. Conflicting with this finding, Bayati et al. 114 observed that ADSCs and BM-MSCs had similar differentiation potentials, although, as Bayati et al. noted, their protocol differed from Meligy et al.'s in terms of the rat species and differentiation method used. At this point, pericytes, ADSCs, UC-MSCs, and BM-MSCs all appear worthy of future exploration in SMTE VML studies, and many of them have already been tested to some extent in muscle therapy studies (Table 3).
DMD, Duchenne Muscular Dystrophy; SMTE, skeletal muscle tissue engineering; VML, volumetric muscle loss.
iPSCs and ESCs are presently less promising than other cell types. iPSCs can be generated from an abundant tissue like skin, can be patient matched, have unlimited self-renewal, and can have high myogenic differentiation capacity in culture. Yet, inducing pluripotency is not efficient, iPSCs are tumorigenic, and most differentiation methods rely on gene overexpression, which is not clinically feasible. Moreover, unlike pericytes, ADSCs, and BM-MSCs, iPSCs do not have immunological characteristics that are useful for allotransplantation.96,149 Although iPSCs are useful in experimental settings and offer some potential advantages over satellite cells, they have more significant barriers to overcome than other stem cells before they can be used in SMTE. ESCs have many of the same limitations as iPSCs, but they are also harder to obtain than iPSCs, carry enormous ethical and legal burdens, and are not patient specific.
Although there is no perfect alternative to satellite cells, based on what is currently known, ADSCs, BM-MSCs, pericytes, and UC-MSCs are particularly promising candidates for SMTE studies. Each offers potential advantages over satellite cells with respect to biopsy, isolation, and purification, while also having high proliferative capacities and significant myogenic differentiation potentials. Future studies should consider using these cell types to develop new SMTE protocols and compare them with current satellite cell-based engineering methods. In the process, it will be important to optimize isolation and differentiation methods across cell types, to work toward clinical-grade SMTE protocols that do not use gene overexpression, exogenous DNA, or media that contain animal products. Ensuring proper myogenic differentiation of alternative stem cells either in vitro or in vivo will be particularly crucial, because, unlike satellite cells, they are capable of differentiating into multiple tissue types. Despite the challenges ahead, alternative stem cells may be the impetus that advances SMTE toward highly functional and practical VML treatments, to give VML patients the quality of life they deserve.
Footnotes
Disclosure Statement
No competing financial interests exist.