
Abstract
Rapid progress has been made in the last decade related to stem cell-mediated pulp–dentin regeneration, from characterization of dental pulp stem cells (DPSCs) to the first-ever reported clinical case in humans. However, many challenges still need to be addressed before such technology can become a common clinical practice; therefore, further rigorous research is needed. Animal study models are very important to test new ideas, concepts, and technologies. This review summarizes and discusses several key animal models that have been utilized to investigate pulp–dentin regeneration. From a tissue regeneration perspective, we categorize the animal model by the location where the regenerated pulp tissue is formed: ectopic, semiorthotopic, and orthotopic. Several animal species are discussed, including mouse, ferret, dog, and miniswine. Mouse is used for ectopic pulp–dentin regeneration in the dorsum subcutaneous space. A commonly tested approach is hydroxyapatite/tricalcium phosphate (HA-TCP) granules model used to observe ectopic pulp–dentin complex formation. The semiorthotopic model includes tooth slices or fragments with which de novo pulp regeneration in a root canal space can be tested in the mouse subcutaneous space. For orthotopic pulp regeneration, the canine teeth of ferrets are large enough for such purposes. As nonprimate large animal models, dog and miniswine teeth have many aspects quite similar to those of humans, allowing researchers to perform experiments that mimic clinical conditions in humans. The protocols established and the data obtained from large animal studies may directly relate to and apply to future human studies. Complete orthotopic pulp regeneration has been demonstrated in dogs and miniswine. The use of allogeneic and subpopulations of DPSCs for pulp regeneration, and testing of the periapical disease model and aging model have been performed in miniswine or dogs. In sum, all these animal models will help address challenges that still face pulp regeneration in humans. We need to thoroughly utilize these models to test new ideas, technologies, and strategies before reliable and predictable clinical protocols can be established for human clinical trials or treatment.
Impact Statement
Animal models are essential for tissue regeneration studies. This review summarizes and discusses the small and large animal models, including mouse, ferret, dog, and miniswine that have been utilized to experiment and to demonstrate stem cell-mediated dental pulp tissue regeneration. We describe the models based on the location where the tissue regeneration is tested—either ectopic, semiorthotopic, or orthotopic. Developing and utilizing optimal animal models for both mechanistic and translational studies of pulp regeneration are of critical importance to advance this field.
Introduction
Developing and utilizing optimal animal models for mechanistic studies and preclinical simulation are key to advancing regenerative medicine. The research progress in the past decade has indicated that stem cell-mediated regeneration of lost pulp and dentin in the root canal space is no longer an impossible task.1–4 A pilot clinical study and a phase I clinical trial in humans have also been recently reported and shown to be a safe therapeutic approach.5,6 Both cell-based and cell-free approaches have been investigated using animal models. The cell-free approach has been practiced clinically for the past several years and has been suggested as an appropriate treatment option for suitable cases. 7 However, the cell-free approach has not consistently shown pulp–dentin regeneration when tested in human teeth and large animals. 7 The cell-based approach is relatively more complex, as it deals with isolating and culturing stem cells in the laboratory; therefore, stem cell transplantation studies rely heavily on animal models to investigate regeneration. The cell-based approach has utilized mice, rats, rabbits, ferrets, dogs, and miniswine to test pulp–dentin regeneration.2,3,8–12 The use of small animals has much to do with their ease of handling and economical nature. Nonetheless, large animals will eventually be needed to test conditions that highly mimic those in humans, especially considering work in the root canal space. Larger animals with tooth sizes comparable with humans allow for testing not just for orthotopic pulp regeneration but also for clinical simulation.
In this review, we outline and critically analyze key characteristics of established animal models in the field of stem cell-based pulp–dentin regeneration research. We describe the models based on the location where the tissue regeneration is tested—either ectopic, semiorthotopic, or orthotopic—in which small and large animal models are perused.
Ectopic Regeneration Model
It is usually the first step and typically involves small animals that are easier to manage. Whether isolated stem cells possess the ability to form pulp–dentin in vivo can be determined using a hydroxyapatite/tricalcium phosphate (HA/TCP) model, which is commonly used for studying ectopic bone regeneration.
HA/TCP model for ectopic pulp–dentin complex formation in mice
This mouse model utilizes osteoinductive HA/TCP granules to drive stem cells differentiation toward osteogenic lineages. 13 Cells mixed with HA/TCP are subcutaneously transplanted into an immunocompromised mouse. If the original cells are bone marrow stromal/stem cells (BMMSCs), they become osteoblast-like cells; if those are dental pulp stem cells (DPSCs) or stem cells of apical papilla (SCAP), they differentiate into odontoblast-like cells. These differentiated cells produce mineral tissues on the surface of HA/TCP granules, and the space between them is filled with soft tissue. With this model, BMMSCs would form ectopic bone and bone marrow, whereas DPSCs or SCAP would form pulp–dentin complexes.
A typical pulp–dentin complex is shown in Figure 1, exhibiting pulp-like tissue containing odontoblast-like cells in the periphery against the mineral dentin-like tissue they produced. When conditions are optimal, the pulp–dentin complex contains well-aligned odontoblast-like cells against the dentin-like structure. Both human (h) and miniswine (s) DPSCs can form a good quality pulp–dentin complex in such a model.12,13 These odontoblast-like cells express nestin, dentin sialophosphoprotein (DSP), and dentin matrix protein-1 (DMP1) (Fig. 1D–F). The dentin-like mineral-contained dentinal tubule-like structures (Fig. 1C–E, yellow arrows) and some odontoblast-like cells showed polarized cell bodies (Fig. 1F).
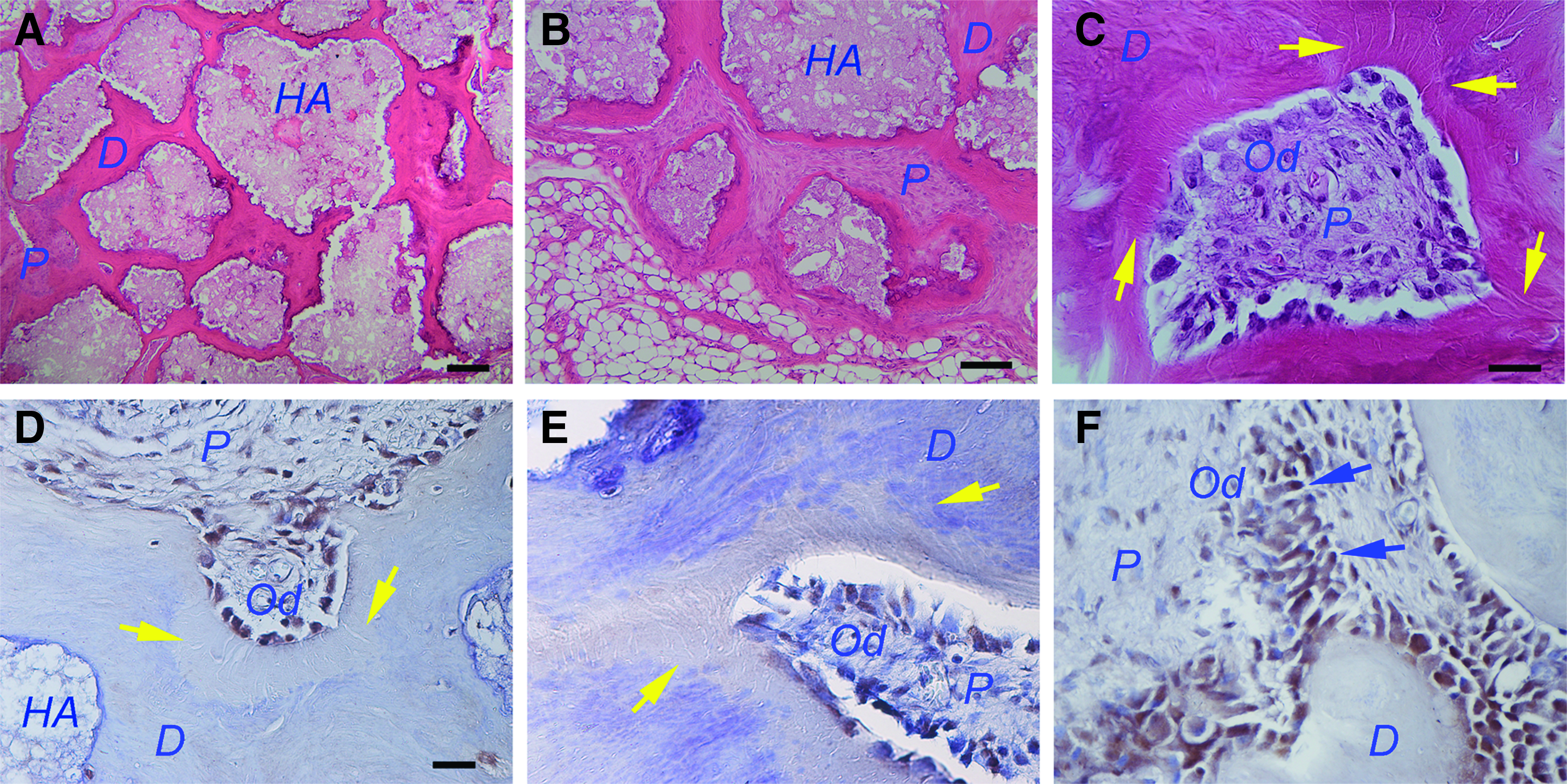
HA/TCP model for in vivo formation of ectopic pulp–dentin complex. Swine DPSCs (passage 3) were mixed with HA/TCP and transplanted into SCID mice. Samples were harvested after 3 months.
Semiorthotopic Regeneration Model
Tooth slice and tooth fragment models are described here. Both can be considered as semiorthotopic for pulp regeneration. Although the tooth is implanted into an ectopic location using immunodeficient mice, the regeneration occurs inside a real tooth.2,8
Tooth slice/scaffold model
The tooth slice/scaffold model 14 was inspired by seminal publication by the Smith laboratory at the University of Birmingham (United Kingdom), which described the development and characterization of the “tooth slice organ culture model.” The original model consisted of tooth slices prepared from incisors extracted from 28-day-old rats that were embedded in agar and cultured at the liquid–gas interface for up to 14 days. 15 This was the first demonstration that dentin–pulp complex of mature rat teeth can be cultured in vitro as a study model for dentinogenesis. A natural extension of this in vitro model was described by Gonçalves et al., who prepared human tooth slices from third molars and transplanted them in the subcutaneous space of the dorsum of immunodeficient mice. 8 They showed for the first time that the dental pulp of permanent teeth survived well the sectioning and transplantation processes. Further, this human dental pulp was found to remain viable for at least 7 days, showing few apoptotic cells. This experimental model is useful for studies of the biology of the human dentin–pulp complex in vivo, as well as for studies that address the impact of treatment [e.g., vascular endothelial growth factor (VEGF)] on the revascularization of severed dental pulps. 16
Stimulated by these early results, the Nör Laboratory began exploring the possibility of using the human tooth slice as a method to understand the mechanisms of differentiation of DPSCs. The reasoning here is that we knew that the differentiation of DPSCs into fully functional odontoblasts requires the physical contact of these cells with dentin and is driven by dentin-derived proteins.17,18 As such, we worked on the development of a tissue engineering-based approach for DPSC transplantation. Here, we were inspired by the characterization of porous poly-
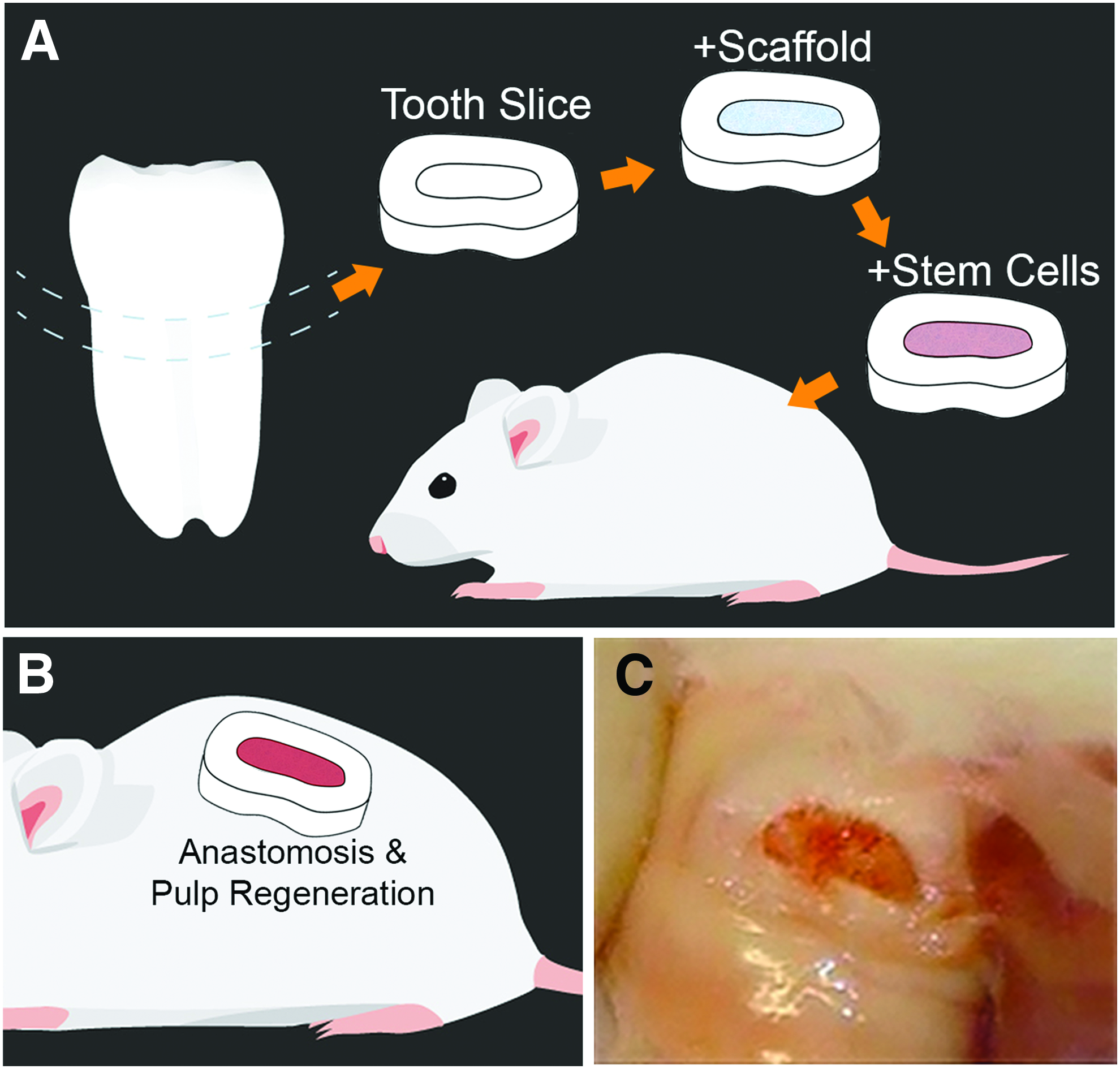
Schematic diagram of tooth slice in vivo model.
A significant advantage of this model is that it enables parallel in vitro and in vivo experiments for both mechanistic and translational studies using a singular platform, thus minimizing the number of experimental variables. For example, to understand the impact of dentin-derived proteins on the odontoblastic differentiation of DPSCs, Casagrande et al. performed parallel in vitro and in vivo studies using the tooth slice/scaffold model. 18 They concluded that dentin-derived proteins are necessary and sufficient to induce odontoblastic differentiation, and subsequently showed that dentin-derived BMP-2 (not BMP-7) was necessary for such differentiation. Moreover, this model is amenable for testing genetically modified DPSCs to understand the impact of a certain signaling pathway on their differentiation capacity. Bento et al. used a shRNA-based approach to silence two major receptors of VEGF; that is, VEGFR1 and NP-1. 21 They observed that VEGFR1 signaling is required for endothelial differentiation of DPSCs, while NP-1 signaling has a more modest impact on this process. More recently, Zhang et al. performed a similar approach to understand the impact of beta-Catenin signaling on DPSCs. 22 They observed that silencing of beta-Catenin rendered DPSCs incapable of fully differentiating into endothelial cells in vivo. Furthermore, in the same study, DPSCs were stably transduced with green fluorescence protein to allow for the unequivocal determination of cell fate upon transplantation. 22
To further improve the translational potential of data derived from the use of the tooth slice/scaffold model, Rosa et al. tested injectable scaffolds such as Puramatrix™ peptide hydrogel, and full human roots became a natural extension of the tooth slice/scaffold model. Rosa et al. demonstrated that transplantation of DPSCs mixed with Puramatrix consistently enabled the generation of pulp-like tissues with histological features that closely resemble those of natural human pulp. 23 Further studies demonstrated that a concentration of 15–20% Puramatrix is ideally suited for the survival and proliferation of DPSCs. 24 In summary, the tooth slice/scaffold model is most suited for mechanistic and translational studies with DPSCs, including the understanding of signaling events regulating DPSC fate or designed to optimize stem cell transplantation conditions for regenerative endodontics procedures.
Tooth fragment model
This model was developed based on R.B. Rutherford's testing of dental tissue regeneration using the apical portion of human roots transplanted into immunocompromised mice (unpublished data). To simulate clinical pulp–dentin regeneration, Huang's Laboratory further extended the concept by sectioning a segment of a human root up to 10 mm long with one end of the canal sealed with mineral trioxide aggregate (MTA) cement. 2 Similar to the concept of the tooth slice model, the tooth fragment model is a semiorthotopic regeneration model that more closely resembles clinical conditions, because not only pulp–dentin regeneration occurs inside a real tooth but also the length of the tooth fragment approximates the actual root. When one end of the root canal space is sealed with cement, the dead-end root canal space is identical to the orthotopic condition that blood supply can only enter from one end. The schematic illustration demonstrates the reliance of nutrition and blood supply from the subcutaneous space of immunocompromised mice to support the inserted cells in the root canal space to survive and regenerate tissues (Fig. 3A). Such an approach was first reported using tooth fragments with a wide canal and large apical opening (2–3 mm in diameter) with which the transplanted DPSCs or SCAP loaded into the canal space may well survive, leading to complete de novo pulp regeneration in the entire canal space with a layer of dentin-like mineral on the canal walls and below the MTA. 2
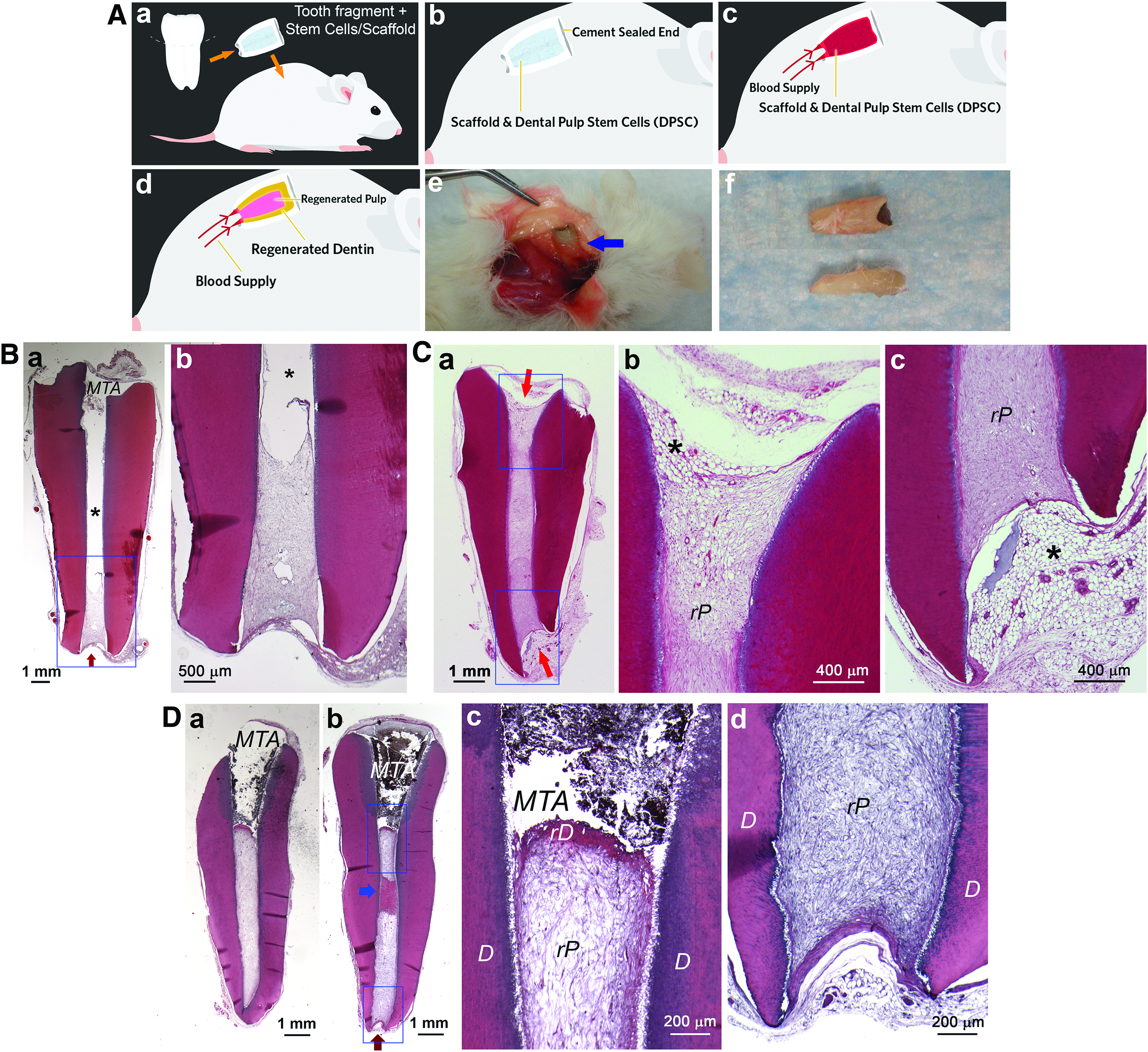
Tooth fragment in vivo mouse model.
However, when longer tooth fragments with narrower canals and small apical openings (≤1 mm) were used, regeneration became inconsistent, as the blood and nutrition supply from the mouse subcutaneous space had difficulty reaching deeper into the canal (Fig. 3B). 12 In such a condition, both the coronal and apical ends of the canal need to be kept open to allow complete pulp regeneration (Fig. 3C). When one end was sealed, pulp regeneration only reached to the middle or apical third of the canal in most samples tested, although complete regeneration did occur in a very small number of samples (Fig. 3D).
Such a tooth fragment semiorthotopic approach has the advantage of having relatively simple procedures while providing an orthotopic-like environment and space for pulp and dentin regeneration. There are, however, obvious disadvantages to such mouse models: (1) blood supply in the mouse subcutaneous tissues is very different from periapical tissues, (2) operating procedures on the tooth samples are very different from working in the clinic, and (3) the regenerated tissues are also produced and populated by mouse cells.
Orthotopic Regeneration Model
Ferret model (Mustela poturios furo)
The ferret canine has been used in endodontic research for many years.25–29 Male descented animals, ∼4 months of age, are typically used for dental research. The mature canine is ∼15–17 mm long, and the clinical crown is sufficient for performing clinical pulpal and endodontic procedures in a realistic manner, even after removing 2–3 mm from the incisal edge to expose the pulp and access the root canal. Earlier work on the ferret canine investigated the biocompatibility of dental materials to pulpal and apical tissues.27,28,30,31 Pulp–dentin regeneration induced by growth factors, such as BMP-7, was described in the ferret canine.32,33
The clinical indication of dental pulp regeneration frequently involves control of microbial factors. Inducing apical periodontitis in the mature ferret canine has been found to be a reproducible disease model.29,34 Mature ferret canine has some apical arborization of the canal system that harbors bacteria after pulp necrosis but does not permeate effective disinfection protocols. The 70-day-old ferret has four erupted but immature canines 35 having open root apex, which lacks the arborization. This allows the study of infection and disinfection in a manner similar to that of human immature teeth. Care should be taken with complete pulp removal on the lingual walls of the canal, beyond the lingual ridge. The reason this is necessary is that in the immature tooth, the pulp has ample vascular supply, and protected portions of the tissue may survive infection and disinfection protocols, thus leading to confounding data.
An earlier report using the ferret model for regenerative procedures showed that the use of blood clot or platelet-rich plasma resulted in the formation of mineralized tissue that mimics osseous or cemento-osseous tissues. 36 Recent studies have isolated and characterized DPSCs from the ferret canine (fDPSCs), 37 which were found to express STRO1, CD90, and CD105, but not CD146, and were highly osteogenic but weakly adipogenic. Quantitative reverse transcriptase-polymerase chain reaction (RT-PCR) experiments revealed that the cells expressed osteopontin, DMP1, DSP, and alkaline phosphatase (ALP) comparable with their human counterparts.
Allogeneic fDPSCs were prepared in oxidized alginate-fibrin hydrogel microbeads, CaCl2, and thrombin. 11 This scaffold had been developed and was shown to release stem cells within 3–5 days. 38 This stem cell-scaffold construct was used to investigate whether its ability to regenerate pulp–dentin was more effective than blood clot-based revascularization protocols in ferret canines after the root canals had been infected and disinfected. The results showed no differences in the percentage of cases in which the healing of apical periodontitis and continued development of the root occurred between the two groups. Importantly, however, this study also showed that there was a highly statistically significant association between the presence of persistent bacteria and the lack of root development. Likewise, the presence of intrapulpal inflammation was significantly correlated with a lack of root development (Fig. 4). These experiments demonstrated that the targeted stem cell/scaffold approach did not improve the process of pulp revitalization, and that the persistence of bacteria interfered with root development and tissue healing, which underscores the importance of developing effective disinfection protocols.

Ferret canine in which the allogeneic stem cell/hydrogel construct was used to regenerate dental pulp. In this case there was incomplete elimination of bacterial infection that caused persistence of necrotic tissue (N), an inflammatory response (I), incomplete root development and apical cemento-osseous repair tissue (B). Color images are available online.
Dog model (Canis lupus familiaris)
Dogs have been widely used as a study model for dental research. Here, we focus on the review of stem cell-mediated complete pulp–dentin regeneration.
Isolation and characterization of canine dental pulp stem cell subpopulations
The Nakashima laboratory has isolated and characterized canine (c) DPSCs, which are similar to their human counterparts. Several DPSC subpopulations have been identified, which not only have high proliferative and multidifferentiation potential in vitro but also possess high angiogenic/vasculogenic and neurogenic potential in vivo for pulp regeneration. Canine dental pulp CD31− side population (SP) cells and CD105/endoglin+ cells were isolated,39,40 which expressed high levels of angiogenic/neurotrophic factors.3,41 However, their isolation was met with challenges in the context of good manufacturing practice; thus, the granulocyte colony-stimulating factor (G-CSF)-induced stem cell mobilization method has been developed for obtaining clinical-grade DPSC subpopulations that circumvent the challenges.42,43 Canine-mobilized DPSCs (MDPSCs) express higher levels of CD105 and stem cell markers with higher proliferation and migration activities compared with the unfractionated pulp cells. The secretome analysis showed higher trophic effects on migration, antiapoptosis, neurite extension, and immunosuppression of MDPSCs than those from unfractionated pulp cells. 44
Complete pulp regeneration by harnessing DPSC subpopulations in canine mature teeth
Canine incisors and premolars are similar to those of humans in their anatomy and the developmental biology of the dentin–pulp complex. 45 Tissue regeneration in dogs may be similarly affected by some factors, including the immune system and genetics to humans46,47; therefore, they can be used as an effective, directly translatable model in the preclinical study of pulp regeneration in mature teeth. The apical part of the tooth root, however, has a “closed apical delta in the adult dog, distinct from the open apical foramen in the adult human” (Fig. 5Aa–d).
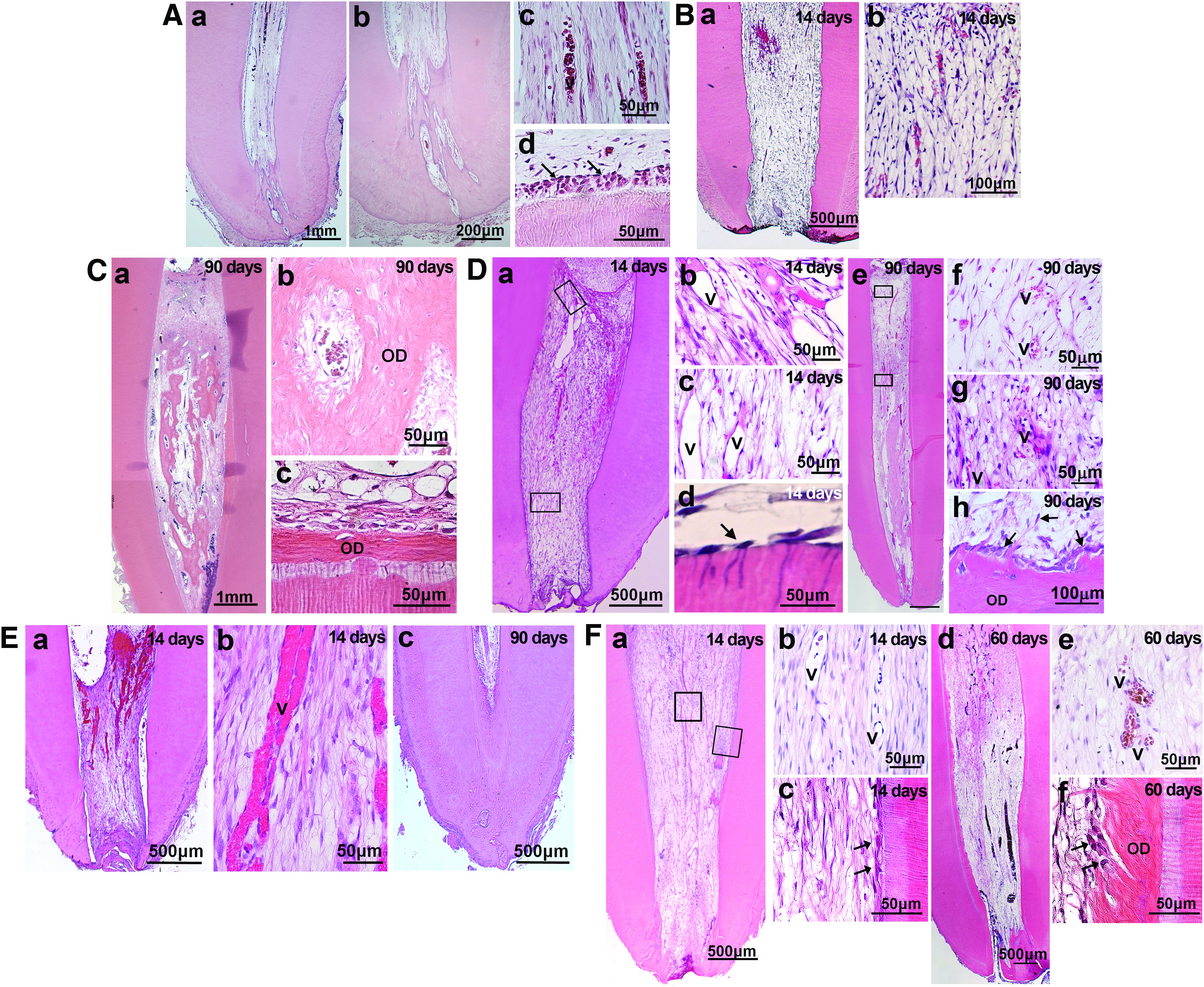
Dog model for orthotopic de novo pulp regeneration.
Adult dog mature teeth with complete apical closure were useful as a pulpectomized tooth model without infection. Stringent criteria were provided for root canal enlargement. Initially, a cell transplantation method with tooth extraction and replantation: ex vivo cell injection in the root canal of extracted teeth before replantation, was used (Fig. 5Ba, b). A 1-mm section of the apical portion of the extracted tooth was cut out followed by whole pulp removal; enlargement of the apical foramen, 0.8 mm in width; biological filling ex vivo; and the tooth transplanted back into alveolar bone. 40 Subsequently, another cell transplantation method with cell injection into the root canal without tooth extraction was employed (Fig. 5C–F). Apical cementodentinal junction was shaped to 0.50–0.60 mm while maintaining the patency with the No. 15 reamer.3,41 Both approaches yielded similar results. Pulp CD31− SP cells and CD105+ cells contain a high rate of CXCR4+ cells with high migration and proliferation activities with stromal cell-derived factor-1 (SDF-1). 48 The amount of well-vascularized and innervated pulp-like tissue was much higher in the transplants of pulp CD31− SP cells or CD105+ cells with SDF-1 (Fig. 5Ba, b; Da–h) compared with unfractionated total pulp cells with SDF-1 (Fig. 5Ea–c). Transplantation of SDF-1 alone, pulp CD31− SP cells, or CD105+ cells alone with collagen scaffold yielded less regenerated pulp tissue. 40 A continuous layer of dentin-like mineralized tissue along the dentinal wall was demonstrated in the transplants of CD105+ cells with SDF-1 (Fig. 5De, h). On the contrary, formation of matrix and mineralized osteodentin-like tissue on day 90 was more prevalent inside regenerated tissue in the transplant of colony-derived DPSCs with G-CSF (Fig. 5Ca–c) and unfractionated total pulp cells with SDF-1 (Fig. 5Ec) compared with those of pulp CD105+ cells with SDF-1 (Fig. 5De). 3 The transplanted CD31− SP cells or CD105+ cells localized in the vicinity of the newly formed capillaries and nerve fibers released trophic factors without direct incorporation into vessels, nerves, or host cells, demonstrating their potent trophic effects. The protein and mRNA expression by two-dimensional electrophoresis, real-time RT-PCR, and microarray demonstrated that the regenerated pulp tissues were qualitatively and quantitatively similar in the transplants of pulp CD105+ or CD31− SP cells with SDF-1 to normal canine pulp.3,41
For preclinical efficacy and safety studies, clinical-grade autologous MDPSCs were transplanted. MDPSCs are enriched not only for CXCR4 but also for G-CSF receptor (G-CSFR) positive cells. 42 Enhanced tissue regeneration in the transplantation of MSCs, together with G-CSF, had been reported in other tissue regenerative therapy, including peripheral nerve injury, spinal cord injury, and cerebral ischemia. 43 Thus, G-CSF was used as a migration/homing factor, together with MDPSCs, in place of SDF-1, which had not been available for human use. Pulp-like tissue was completely regenerated with vasculature and innervation in the root canal (Fig. 5Fa, b, d, e). Dentin-like mineralized tissue was formed along the dentinal wall (Fig. 5Fc, f) and also in the coronal part to prevent microleakage. The regenerated tissue was further confirmed to be identical to normal functional pulp tissue by expression of pulp biomarkers, thyrotropin-releasing hormone (TRH)-degrading enzyme. Hierarchical cluster analysis of gene expression patterns among normal pulp, regenerated pulp, and periodontal ligament also demonstrated identification of regenerated tissue. 44 DiI-labeling demonstrated nerves extending to the trigeminal ganglion, suggesting that sensory signals may be transmitted from the regenerated tissue. There was no evidence of toxicity or adverse events for the 3-month study duration in dogs. 44
The underlying mechanisms of pulp regeneration by MDPSC transplantation with G-CSF were demonstrated in dogs. There were three critical criteria: (1) MDPSCs migrate to the apical part before vascularization due to high migratory activity, and survival by the antiapoptotic effect of G-CSF. G-CSFR-positive MDPSCs are confined in the root canal by G-CSF. (2) MDPSCs highly express various trophic factors to stimulate migration and proliferation of endogenous stem/progenitor cells from adjacent tissue and promote angiogenesis/vasculogenesis, reinnervation, and reduce apoptosis and inflammation. (3) G-CSF has combinatorial effects with MDPSCs on stimulated cell migration, neurite outgrowth, antiapoptosis, and immunosuppression. 44 In conclusion, these results of preclinical studies in dogs, and the scientific demonstration of the safety and efficacy, were established before clinical applications in humans.
Complete pulp regeneration with periapical disease
Recently, Nakashima's group demonstrated successful pulp regeneration by the stem cell therapy in a canine model of periapical disease similar to the pulpectomized tooth model in mature teeth. The root canal was kept open for >2 weeks, and infection was confirmed by microbiological examination of the extracting bacteria and periapical lesions by cone beam computed tomography. It took more time to regenerate whole pulp tissue in the periapical disease model (data not shown), in which the periapical tissue, including alveolar bone, was to be regenerated. It is a challenge to develop the optimal method for both pulp and periapical tissue regeneration in the canine model of periapical disease in mature teeth.
From a clinical standpoint, in necrotic immature permanent teeth, instrumentation must be avoided to further prevent weakening of fragile root dentinal walls. 49 Alternatively, root canal irrigation, along with placement of an antibiotic paste (triple antibiotic paste, TAP), has been the preferred disinfection strategy in blood clot-based revascularization protocols. 49 Regrettably, highly concentrated antibiotic pastes have been linked to severe cell toxicity,50,51 interference with dentin-embedded growth factors release, 52 crown discoloration, 53 and potential antiangiogenic activity, because of minocycline present in the conventional TAP. 54
To overcome the challenge of inherent toxicity of currently used disinfection agents, the Bottino group established a novel biocompatible disinfection strategy through the use of antibiotic-eluting electrospun polymer nanofibers.55–61 In electrospinning, the polymer solution can be loaded with one or multiple antibiotics to generate a three-dimensional (3D) drug delivery construct, which can be positioned inside the root canal system (Fig. 6). 62 The theory behind using antibiotic-eluting nanofibers as an intracanal drug delivery system is based on the fact that adding low antibiotic concentrations and slow drug release provided by these nanofibers will ablate infection and ultimately create a bacteria-free niche favorable to tissue regeneration. A series of studies have recently reported that those antibiotic-eluting nanofibers can eliminate dentin biofilm with minimal toxicity.58,59 In vivo studies using a canine model of periapical disease in immature permanent teeth are currently pursued. In a preliminary study, immature permanent double-rooted premolars have been used to test the antimicrobial capacity of the innovative drug delivery construct when compared with the antibiotic paste (TAP, 1 g/mL) and the regenerative response after the induction of bleeding. Three interventions were conducted: (1) periapical lesion induction, (2) disinfection, and (3) evoked bleeding through the introduction of a precurved K-file past the apical foramen to allow blood invasion. A collagen scaffold was placed over the blood clot before tooth restoration. Overall, the histological findings suggest that the drug delivery construct can ablate periapical infection and mediate formation of an osteodentin-like tissue at the apex.
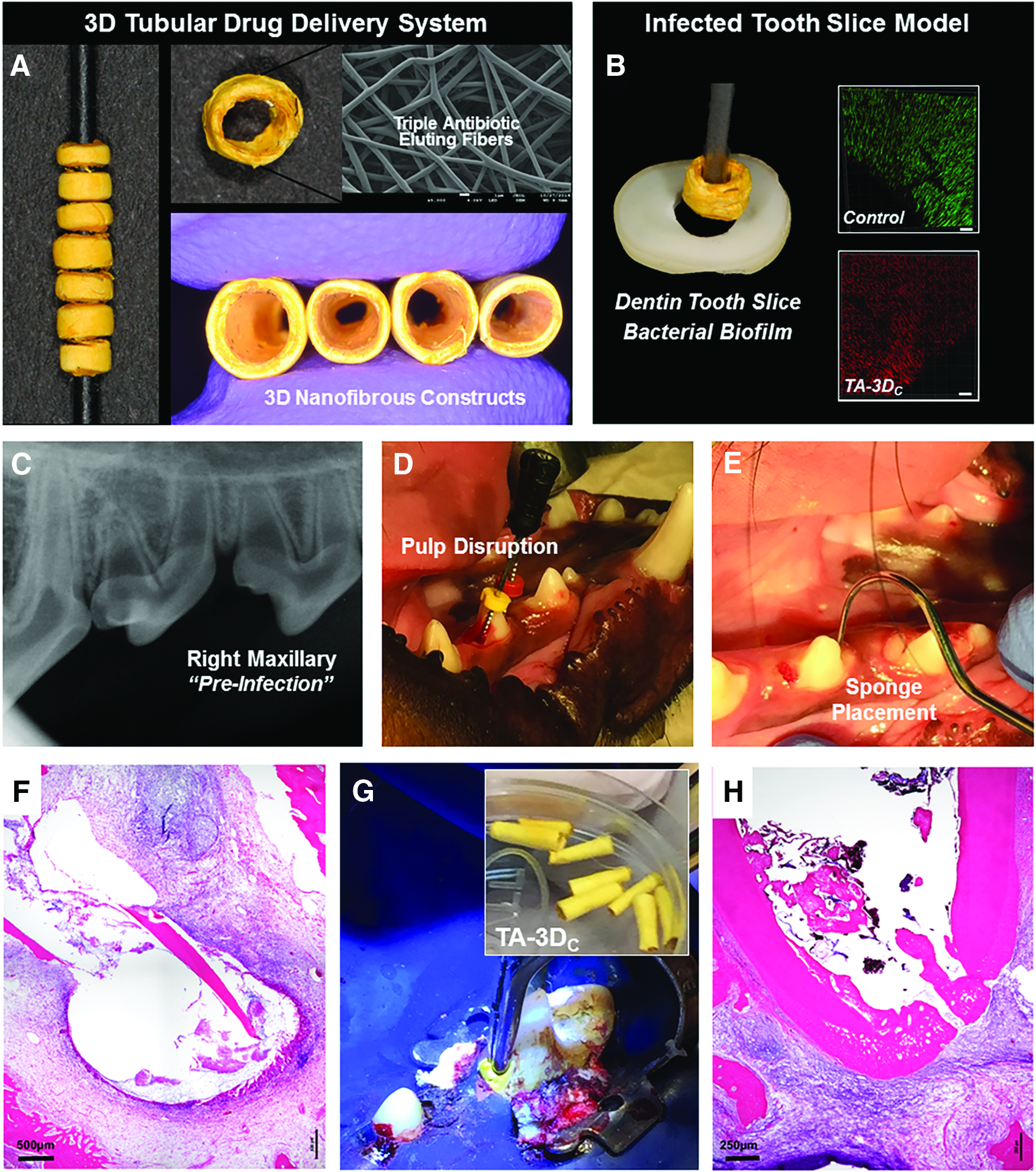
Summary of the clinical prospects associated with the use of TA-3DC for root canal disinfection before regenerative endodontics.
Complete disinfection by biocompatible intracanal medicaments is key to predictably achieve both pulp and periapical tissue regeneration. 11 Further improvement of the enhanced delivery of medicament deeper into the root canal system by nanobubbles with pressurized air (Nakashima and Iohara, unpublished); erbium:yttrium aluminum garnet laser (such as photon-induced photoacoustic streaming) 63 ; and improvement of the long duration of medicament by nanoparticles 64 are being studied.
Complete pulp regeneration for the aged
Pulp regenerative therapy in aged patients is a challenge. One plausible solution is using MDPSCs from aged donors that express high levels of trophic factors, similar to MDPSCs from young donors derived from humans and dogs.65,66 The regenerative potential of aged MDPSCs was similar to that of young MDPSCs, as demonstrated in a semiorthotopic tooth transplantation model. Of note, pulp regeneration was reduced after autologous transplantation of aged MDPSCs compared with transplantation of young MDPSCs in the canine pulpectomized model. Lower proliferation, migration, and antiapoptotic activities were demonstrated in the periodontal ligament stem cells derived from aged dogs, representative of migrating endogenous stem cells from the surrounding tooth tissue. The age-dependent decline in pulp regeneration might be attributed to the reduced regenerative capacity of endogenous stem cells.
Miniswine model (Sus scrofa)
Miniature swine resemble humans in many aspects, and they have been utilized as a large animal model in multiple fields of biomedical research, including dental research.67–71 Recently, the Huang laboratory demonstrated that miniature swine can be used as a large animal model to investigate orthotopic stem cell-based pulp–dentin regeneration, including the use of single- and multirooted teeth. 12 Swine (s) DPSCs are similar to their human counterparts with subtle differences in morphology and are indistinguishable in their ability to form pulp–dentin complex in the HA/TCP model. 12
De novo regeneration of pulp and dentin-like tissue in miniswine teeth
In this miniswine model, the heterogeneous population of swine (s) DPSCs were used to achieve complete pulp regeneration, including single-rooted and multirooted teeth. Isolated swine heterogeneous population of sDPSCs emerged from cultures showing typical MSC characteristics. Swine DPSCs have ∼4% STRO1, 40% CD146, 100% CD90, and <1% ALP-positive cells in the population, and are CD34 and CD45 negative. 12 They are highly odonto/osteogenic, while lacking adipogenic potential. The capacity of sDPSCs to form pulp–dentin complex was first tested using the aforementioned HA/TCP model with dentinal tubule-like structures formed in dentin-like mineral (Fig. 1).
When sDPSCs are transplanted orthotopically, de novo pulp regeneration, along with newly generated dentin bridge-like mineral underneath the filling material and dentin-like tissue along the canal walls, can be observed (Fig. 7Aa–g); however, dentinal tubules were hardly detected. A possible cause for this may be the lack of a sufficient number of new odontoblasts formed on the dentinal wall 72 or a lack of original signals, such as a preameloblast-derived factor, copine 7, which only exists during the early stage of tooth development when odontoblasts are maturing from their precursors. 73 Despite a lack of dentinal tubules, nestin, DSP, DMP1, and bone sialoprotein are detected in odontoblast-like cells 12 (Fig. 7Ba–c).
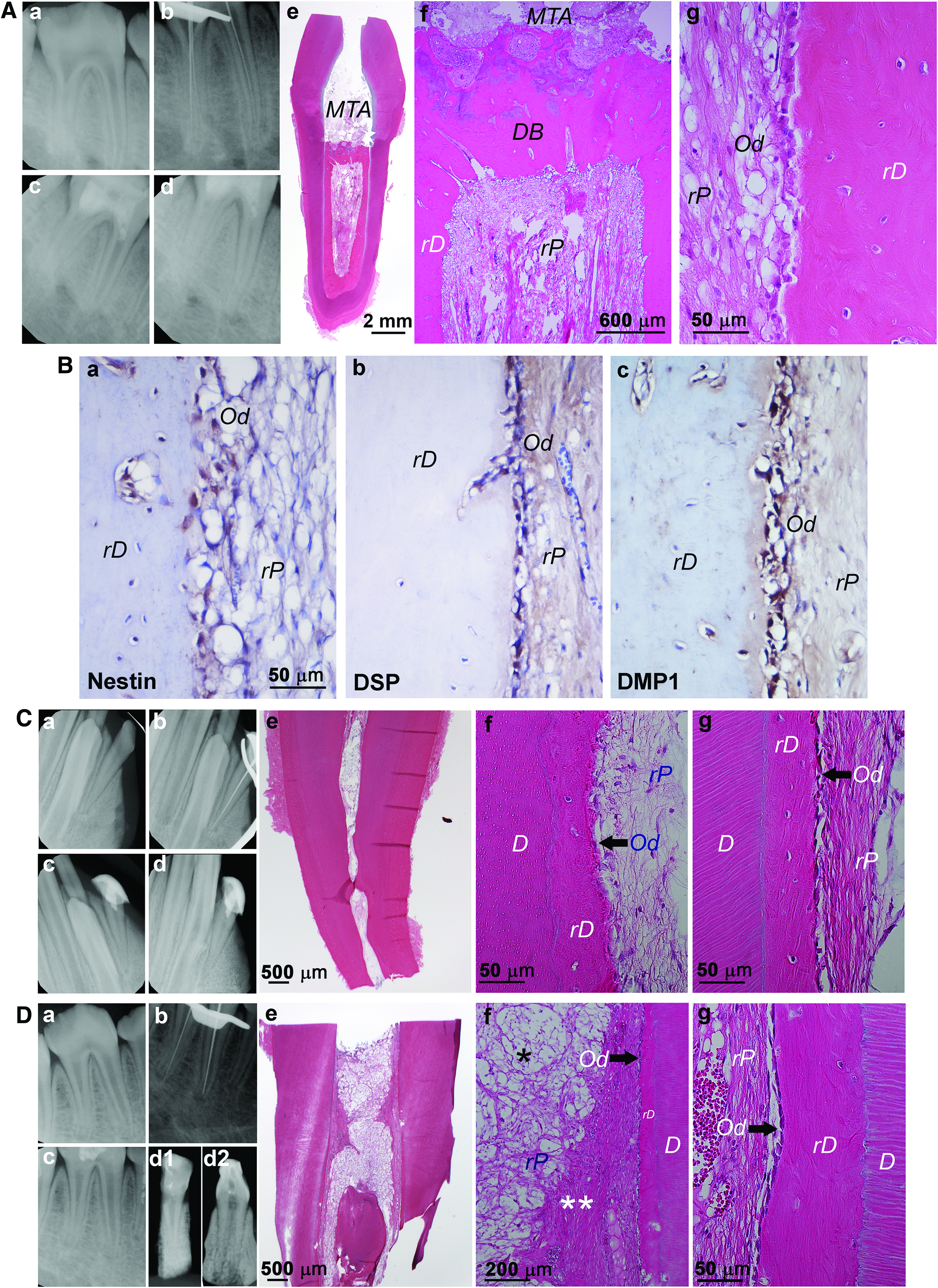
Orthotopic de novo pulp regeneration in miniswine.
In terms of the type of teeth suited for studying pulp regeneration in miniswine, second lateral incisors are similar to human maxillary lateral incisors or mandibular incisors. Molars are too complex and too posterior to manage. Premolars, in general, are similar to human premolars or molars in terms of size and root canal morphology while having two to three roots. First premolars are normally too small to operate. Second to fourth premolars are potentially suitable for experiments. Maxillary fourth premolars resemble human first molars with complex root and canal morphologies that may be quite challenging to operate. Mandibular second and third premolars represent multirooted teeth that are easier to operate. Mandibular fourth premolars are similar to human molars from a clinical management perspective.
Allogeneic DPSCs potentially be used for pulp regeneration
The use of an autologous cell source for tissue regeneration is considered the optimal approach, because such cells are less likely to be rejected by the host. A major drawback, however, is their availability at the time of clinical need. Even if available, the clinician needs to extract the tooth and send it for cell isolation, quality testing, etc., which takes at least a month with no control over whether the cells will be of sufficient quality for clinical use. In case allogeneic cells can be used, these cells would have been tested for their quality and safety, and can be cryopreserved as off-the-shelf products ready for use. If proven possible, stem cell-mediated pulp–dentin regeneration technologies may then become more relevant and prevalent for clinical practice.
Although preliminary, findings from the miniswine model suggest that allogeneic DPSCs may be useful for pulp regeneration. As shown in Figure 7C and D, one is a mandibular second lateral incisor receiving cells of a different strain (Sinclair to Yucatan) (Fig. 7Ca–g), and the other is a mandibular third premolar receiving cells of the same strain (Yucatan to Yucatan) (Fig. 7Da–g). Similar regeneration in the canal space can be seen compared to using autologous DPSCs. Recent report from Nakashima's laboratory demonstrated that complete orthotopic pulp regeneration in dogs using allogeneic MDPSCs is safe and can yield similar results to using autologous counterparts. 74 The tolerance of transplanted allogeneic DPSCs is likely to be the result of immunosuppressive properties that mesenchymal-type stem/stromal cells normally possess, which can suppress immune rejection from the host and be sustained.75–77 Despite this observation, a more stringent allogeneic transplantation study should be conducted by examining a long-term tracing of the transplanted cells.
Prospects
Pulp–dentin regeneration technology is the ultimate strategy to restore tooth functions and avoid tooth loss. While current research progress has indicated the impending arrival of pulp–dentin regeneration technology to clinical endodontics, there are still multiple challenges to overcome; for example, (1) disinfection efficiency and (2) blood supply during regeneration. Without more effective and biocompatible root canal disinfection protocols, such stem cell-based procedures are likely not to be suited for heavily infected root canals with periapical lesions. New technologies have to be developed and tested such as the aforementioned nanofibrous antibiotic-eluting strategy, laser activation, acoustic energy, and nanobubbles. The blood supply for long and multirooted teeth with narrow canal space is another significant challenge. The established animal models mentioned herein are likely to be very important for testing new ideas, concepts, or technologies and to provide relevant information to help establish optimal protocols for future clinical use.
Footnotes
Acknowledgments
This work was supported in part by grants from Council for Science and Technology Policy, chaired by Prime Minister (M.N.) and the Research Grant for Longevity Sciences (23–10) from the Ministry of Health, Labour and Welfare (M.N.) in Japan; the National Institutes of Health R01 DE21410 (J.E.N.), R01 DE026578 (M.C.B.), R01 DE019156 (G.T.-J.H.) and a Research Fund from the University of Tennessee Health Science Center.
Disclosure Statement
No competing financial interests exist.