
Abstract
In a native bone marrow tissue microenvironment, mesenchymal stem/stromal cells form an important constituent of the stem cell niche, which is composed of extracellular matrix (ECM), niche cells, biophysical cues, and growth factors. These microenvironmental elements play a major role in guiding stem cell behavior, such as growth, stemness, maintenance, and lineage differentiation. The rapid progress in biomaterials advancement and stem cell biology has opened new directions in stem cell therapy. Ideally, biomaterials intended for tissue regenerative medicine applications should perform the structural and biochemical functions of the native ECM, which provides cells with physical (structural/morphological), chemical, and mechanical cues through its three-dimensional (3D) architecture, until the cells' own ECM takes over. Advanced techniques related to material synthesis are significantly important for designing temporary tissue engineering scaffolds, which not only support cell growth but also facilitates 3D tissue formation. This review concisely describes the types of natural and synthetic polymeric biomaterials and the different approaches of designing and developing porous scaffolds for the expansion of hematopoietic stem cells (HSCs). It also illustrates the relationship between material science and tissue engineering and reviews the most frequently used materials and some exciting recent advancements in scaffold manufacture technologies. Furthermore, this review focuses on an introduction to existing HSC niche concepts and summarizes approaches to mimic them in vitro.
Impact Statement
This review discusses designing novel biomaterial-based hematopoietic stem cell (HSC) expansion strategies that would contribute to the field of hematopoiesis and also proposes possible approaches for HSC expansion using interpenetrating network hydrogels, emulsion templated polymers poly(HIPEs) (high internal phase emulsion templated polymers), and three-dimensional cell printing, which could provide optimal environment for HSC attachment, proliferation, and differentiation. These novel approaches could improve the efficacy of bone marrow transplantation and also offer new insights in the field of regenerative biology.
Introduction
Regenerative medicine, tissue engineering, and stem cell-based therapies are some of the most promising approaches for modern medicine, offering the prospect of developing new treatments with the possibility to replace missing, injured, or diseased tissues and organs.1–3 However, their success depends on finding the most effective and clinically suitable strategies to engineer biocompatible scaffolds and microenvironments suitable for cells to expand, proliferate, and differentiate.
To exploit the efficacy of stem cell-based treatments by transplanting cells into patients or stimulating expansion of endogenous cells, it is important to comprehend the microenvironmental (niche) signals, such as physical and mechanical cues4–7 and cell–cell 8 and cell–matrix 9 interactions that regulate stem cell behavior (Fig. 1). The components of the engineered extracellular matrix (ECM) dictate the mechanical performance (stiffness) and porosity of matrix, and also the types of cell–cell and cell–matrix adhesions and interactions. The matrix porosity regulates the ease of nutrients and oxygen diffusion to tissues, and the ability of cells to move throughout the matrix.
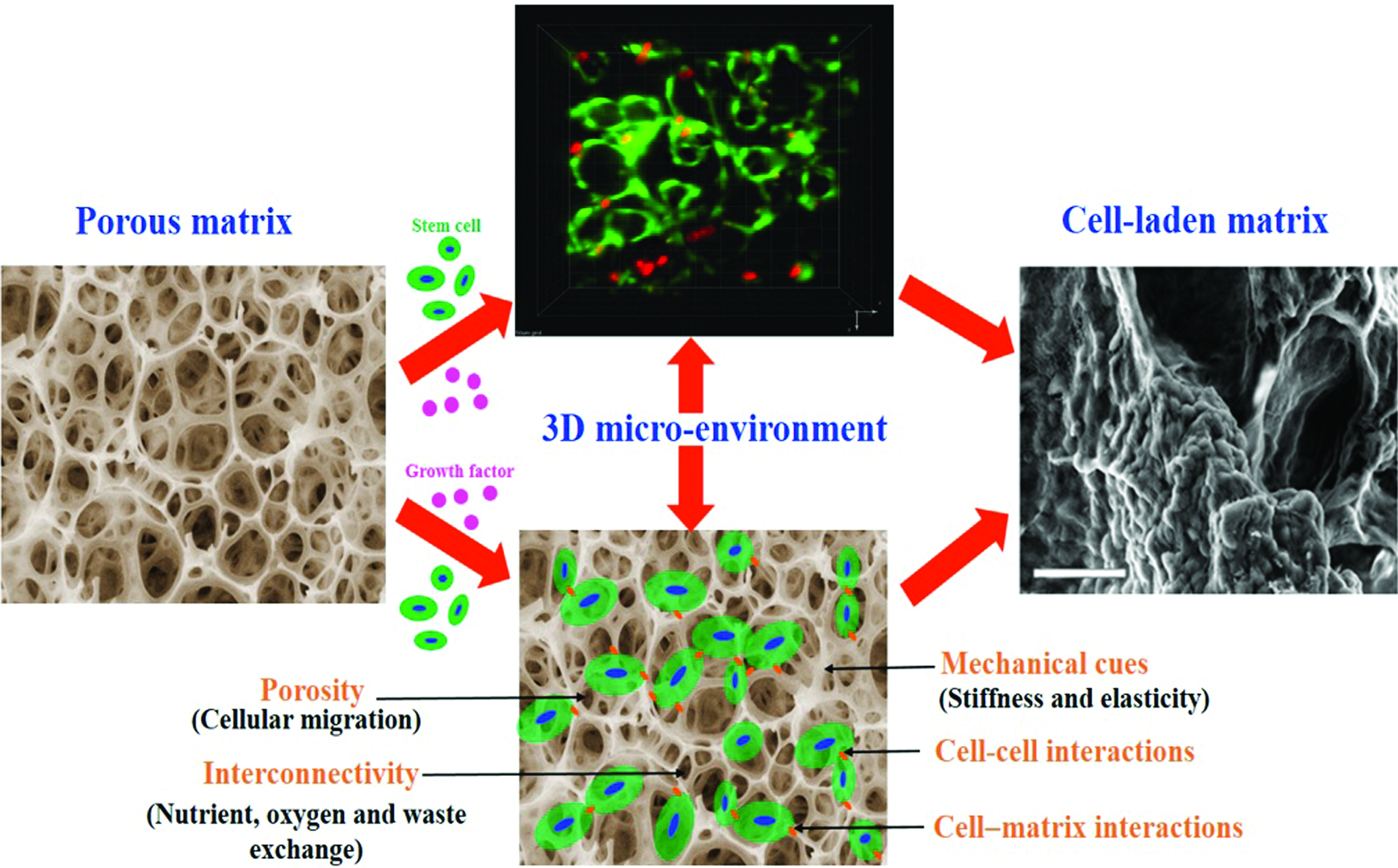
An engineered 3D biomaterial environment provides mechanical and physical cues that guide cell growth and function. 3D, three-dimensional. Color images are available online.
A synthetic mimic of the native ECM is a crucial requirement for in vitro expansion of primary cell with user-tunable composition, including physical and mechanical cues that would permit their long-term viability and other cellular functions.10–13 Physical interaction of the cell with its surrounding tissues is important for adhesion-dependence cellular functions such as shape, survival, migration, expansion, survival, and differentiation. 14 Cell–matrix interactions permit a cell to probe into many properties of the ECM and the cells, but to study such systems systematically, an ideal microenvironment with precise parameters are desirable. Traditional monolayer cell culture models involve use of two-dimensional (2D) substrates that are flat and rigid for the expansion of cells. Cells acclimatize to this engineered 2D environment, become flattened, and act in an anomalous and nonphysiological manner. Moreover, traditional 2D culture substrates made up of plastic and gel cannot mimic geometric, anatomic, and topographic features that are only possible in three-dimensional (3D) cultures, making them less relevant to tissue regeneration approaches.15,16
Surprisingly, the cells residing in soft-tissue microenvironment are very much sensitive to the elasticity of that microenvironment. 17 Therefore, there is significant need for an ideal 3D cell culture model that would enable the cells to expand and acclimatize to their surrounding environment in a way that more intently mimic their native counterparts. There are various benefits in allowing cells to gain a natural 3D phenotypic characteristic, including improved cell proliferation, differentiation capability, and function. Physical and mechanical properties of cellular microenvironment play a major role in guiding cell behaviors such as cell adhesion, migration, growth, survival, proliferation, and differentiation.13,18,19 Cells naturally reside in a specialized and dynamic 3D niche in their biological environment, and collecting evidence specifies that cells may react diversely when placed in 2D and 3D microenvironments.20–22 As the physical forces in the native ECM environment are known to have a significant influence on cell behavior, tissue engineers have developed many material-based systems to probe these interactions.
Hematopoietic stem cells (HSCs) have the distinctive property of self-renewal and differentiation toward all functional blood cells, which makes them a promising tool to be used in patients. 23 There is a constant rise in demand for HSC transplantation (autologous and allogenic) because of the increasing incidence of hereditary and hematologic malignancies. 24 Understanding the mechanisms that regulate the fate of HSCs has been challenging for several years and till today several questions remain unanswered. 25 Such challenges impose a major constraint in the medical use of HSCs. The number of deceased patients who required HSC transplantation surpasses that of matching donors. 26 One of the strategies that has been implemented to prevail over this issue is to use HSCs from umbilical cord blood units27,28; however, the dose of stem cells in one cord blood unit is too little for effective reconstitution of the hematopoietic system. 26 The second strategy was to focus on ex vivo expansion of these cord blood units using different methods. 29 With the emergence of latest gene editing tools like Crisper-Cas9, expansion of gene-edited stem cells has become another important forefront area of research.
HSCs have a tendency to quickly differentiate when cultured in vitro, hence one of the main challenges of ex vivo expansion of HSCs is to be able to expand them without losing their stem cell-like characteristics. 25 Several studies have reported that coculturing HSCs with mesenchymal stem/stromal cells (MSCs) mimics their natural microenvironment in the bone marrow (BM) and at the same favors proliferation of HSCs while preserving their stem cell phenotypic characteristics.30,31 Artificial remodeling of niche constituents in a single biomaterial is therefore an important approach to simulate the in vivo HSC niche with the motive to control its behavior in vitro. 26
In the past decade, several groups have demonstrated the advancement of biomaterial-based 3D cell culture platforms to facilitate in vitro interactions or exchanges between the HSCs and their local microenvironment. 32 A wide range of 2D and 3D culture-based biomaterial experimental approaches have already been utilized extensively for in vitro HSC expansion.24,26,30,31,33–35 These approaches capture the physiological conditions of native niches 36 based upon the current information of the in vivo situation. 33 The basic criteria for such design systems include ease of fabrication, ability to recognize a combination of physical and biological cues, ease of scaling-up for clinical use, and ease for high-throughput analyses that together display a minimal level of complexity, but at the same time also enable a more articulated understanding of HSC regulation and function.24,32,33
This review primarily focuses on proposing novel approaches for HSC expansion using emulsion templating, 3D cell printing, electrospun nanofiber and interpenetrating network (IPN) hydrogel-based scaffolds as the next promising strategy for the ex vivo expansion of HSCs, and also explores the existing approaches taken for the purpose.
HSCs and Their Niches
HSCs are multipotent, long-term self-renewing cells that initiate and maintain life-long hematopoiesis. 37 These cells are not haphazardly disseminated throughout the body but rather localized to specific tissue environment or niches in the BM. 38 The quiescence, adhesion, migration, proliferation, and differentiation of the HSCs during hematopoiesis are primarily regulated by complex multidimensional BM niche composed of biochemical (cells, chemokines, cytokines, and growth factors) and physiochemical (mechanical stiffness, micro/nano topography, of tissues, and O2 concentration) factors 24 (Fig. 2). A well-controlled native microenvironment (niche) exists in the BM that controls the quiescence, proliferation, maintenance, and differentiation of HSCs. 39 The BM niche exudes regulatory signals or cues including secreted and membrane-bound molecules from adjacent cells that are essential for HSC maintenance.40,41
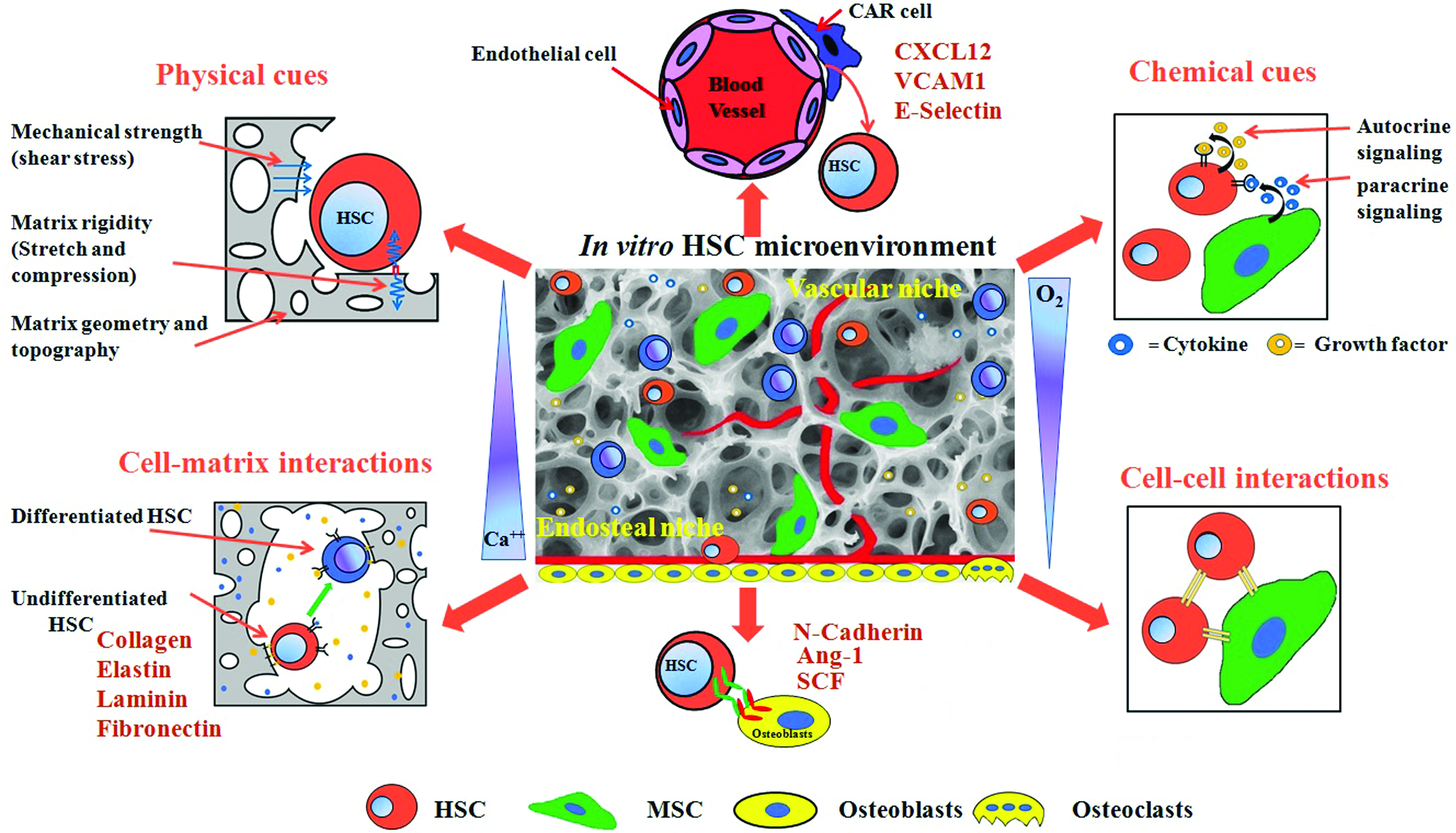
The cartoon illustrates the component of the HSC microenvironment in the bone marrow. Microenvironmental cues such as soluble chemical factors (growth factors and cytokines), cell–matrix interactions (collagen, laminin, fibronectin, and hyaluronic acid), cell–cell interactions (cell adhesion molecules, cadherins, and notch signaling), mechanical forces (matrix rigidity, shear stress, and compression) converse in an intricate way to influence various signaling pathways and eventually regulate HSC fate. HSC, hematopoietic stem cell. Color images are available online.
Furthermore, physical cues such as mechanical shear stress, surface rigidity, elasticity, oxygen tension, and ECM topography play a major role in HSC maintenance.9,42 The niche environment consists of ECM proteins such as laminin, fibronectin, and collagen (types I, III, and IV), and the ECM remodeling proteins such as matrix metalloproteinases that modulate HSC functions during hematopoiesis. HSCs express a range of integrins and the integrin-mediated HSC–ECM interactions have been implicated in determining the HSC behavior.32,43 In addition, the expression of cell-regulating molecules such as ECM protein transforming growth factor-β-inducible gene h3 (BIGH3), adhesion molecule (Endolyn/CD164), activated leukocyte cell adhesion molecule (ALCAM/CD 166), biglycan, and embigin may differ thereby altering the functional capacities of the HSCs. 43 The BM microenvironment also contains many different cell types including osteolineage cells, adipocytes, MSCs, C–X–C motif chemokine 12 (CXCL12)-abundant reticular (CAR) cells, adventitial reticular cells, megakaryocytes (MKs), endothelial cells (arteriole and sinusoidal) and is affected by chemokines, cytokines, and other physical factors, such as mechanical shear stress and local oxygen tension, all of which regulate the fate of HSCs. 44
It is fairly recent that an awareness regarding HSC niche and its general significance as well as implications in disease and medicine has developed because of the advancement in the field of stem cell biology.45–47 Identification of markers that reliably identify the HSCs and niche cells was the first step toward understanding the job of the niche in regulating the HSCs.39,47 It has been reported that the BM consists of two niche compartments namely the endosteal niche and the vascular niche and that both these compartments house HSCs at distinct developmental state.31,38 The endosteal niche is thought to favor HSC quiescence, whereas the vascular niche allows differentiation and movement of the HSCs into the peripheral circulation. 31
Although endosteal and vascular niches contribute in hematopoiesis, the difference between the two is not totally comprehended. One probability is that despite the fact that the two compartments of niches function in an essential way, they perhaps spatially are isolated in the BM. Another school of thought is that each niche has a discrete responsibility during hematopoiesis, and therefore, each signifies a part of several niches in the hematopoietic system. 48 Furthermore, Itkin et al. have shown that the differential permeability properties and reactive oxygen species (ROS) levels of the sinusoidal endothelial cells of the BM regulate the HSC maintenance and the trafficking of the differentiated blood cells out of the BM. 49 The less permeable sinusoids with less ROS promote HSC maintenance, whereas the more permeable sinusoids with more ROS state promote HSC activation and their movement.
More recent studies have pointed out that osteogenic progenitors (and not osteoblasts) influence HSC counts in the BM50–54 and that the HSC progenitors have a tendency to preferentially get distributed along the endosteal region. 55 The interactions between the HSCs and spindle-shaped N-cadherin+ CD45− osteoblastic cells are critical for maintaining the quiescent nature of HSCs residing in the endosteum. Although several articles still discuss the possible role of N-cadherin in HSC maintenance,56–58 a knockout study on mouse showed evidently that N-cadherin has no influence on HSC levels whatsoever.54,59 Other osteoblast and HSC surface molecules that have been shown to promote HSC quiescence in the endosteal niche include the angiopoietin-1 and angiopoietin receptor-2 (Tie 2) interaction. Ang-1 has been shown not only to promote HSC quiescence, but also to enhance HSC survival and protect them from various stresses.60,61 Sacchetti et al. showed that CD146+ osteoprogenitors, and not their differentiated osteoblastic progeny, expressed Ang-1 and were responsible for the vascular remodeling of the hematopoietic microenvironment. 62
Another vital factor that has been shown to guide and favor HSC localization in the endosteal region is the oxygen gradient. HSCs are typically situated at the lowest side of the gradient because the regionally defined functional hypoxia plays a major role in modulating stem cell function in the BM cavity. 63 Moreover, several other factors in the endosteal ECM such as parathyroid hormone (PTH), osteopontin, and stromal cell-derived factor-1 (SDF-1) further contribute toward controlling HSC quiescence and activation in the endosteal niche.32,38 Kuznetsov et al. demonstrated that the expression of PTH and PTH-related peptide receptor also regulated the establishment of skeletal stem cells in the BM and hematopoiesis in the bone. 64
During embryonic development the HSCs arise from the perivascular progenitors and hence it is not surprising that the vasculature play a significant role in the formation and expansion of HSCs. 31 Like the osteoblast progenitors, the endothelial cells promote HSC maintenance and normal endothelial function is a requisite for normal hematopoiesis. 39 In this regard, the endothelial cells intrinsically secrete endothelial cell adhesion molecule E-selectin, vascular cell adhesion molecule 1, and chemokine protein such as CXCL12 that are essential for HSC homing, mobilization, and engraftment. 31 In addition, adventitial cells in a small population with long processes termed CAR cells express highest amounts of CXCL12.38,46 These CAR cells are perivascular MSCs that secrete 100-fold higher levels of CXCL12 than endothelial cells and 1000-fold higher levels of CXCL12 than osteoblasts. Several studies provide strong evidence that HSCs live in a perivascular niche in which endothelial cells and MSCs each secrete various soluble factors that promote HSC survival, localization, and maintenance.31,38,39
Mesenchymal stromal cells also called mesenchymal stem cells constitute an important element of the hematopoietic niche. 65 They are defined as multipotent cells equipped for differentiating into adipogenic, chondrogenic, and osteogenic lineages. 66 MSCs have low immunogenicity, and because of their immunomodulatory characteristics, which influence a wide range of distinctive and adaptive immune system responses, make them highly attractive for clinical applications in cell-based therapies. 67 They colocalize with HSCs in BM niche and produce factors that orchestrate HSC fate in the BM niche. 68 Several studies have shown how intercellular communication between the MSCs and HSCs affects the functionality of HSCs.69,70
Accumulating evidence suggests that the therapeutic effects (regenerative properties) of MSCs are mainly attributable to their paracrine effects.71,72 True HSC maintenance and self-renewal in the initial population has not been achieved when MSCs cocultured with HSCs using 2D systems simply because the geometry is not adequate for cell–matrix and cell–cell interactions and exchanges.73,74 It is evident that 3D culture results in more in vivo-like environment65,75 and the 3D MSC–HSC coculture approach led to statistically significant higher expansion of cord blood CD34+ cells, compared with 2D cocultures.34,76 Therefore, a 3D MSC–HSC culture environment that mimic the native in vivo hematopoietic compartment would be more beneficial for guiding HSC maintenance and expansion.
It is now recognized that, apart from biologically active components like cytokines, MSCs also secrete a large number of extracellular vesicles (EVs). Current ex vivo and in vivo studies have shown that MSC-EVs could not only expand HSCs in vitro, but could also improve outcome of hematopoietic stem cell transplantation and reduce graft versus host disease.77–79 Therefore, MSC-EVs are considered to be promising candidates (as cell-free biologics) for mediating tissue regeneration.77,80
Constructing Biomimetic BM Microenvironment Using Biomaterials for HSC Expansion
Mimicking the native in vivo microenvironment under which HSCs can self-restore and multiply while preserving their stem cell characteristics is a promising approach for in vitro growth and expansion of HSCs. A unique approach to mimic the native BM microenvironment in an in vitro set up is to use different 2D and 3D biomimetic biomaterials derived from natural and synthetic sources. Nevertheless, the success of biomimicking materials in a laboratory setting is dependent upon several factors and interactions, such as the selection of the initial material, type of target cell for which the 3D biomaterial will be used, and other biochemical and biophysical cues or signals that are integrated in to the constructed material.
Natural versus synthetic biomaterials for HSC expansion
A large number of biomaterials have been used because of the advantage of user-defined tunability of cell–matrix interaction as an artificial matrix platform for HSC expansion. Several efforts have been made to create engineered BM-mimicking platforms using natural and synthetic polymers for HSC expansion (Table 1). However, most of these studies did not address the engraftment ability of the cultured HSCs using in vivo experimental studies, the most important parameter indicative of HSC functionality.
Various Biomaterial-Based Strategies Used to Guide Hematopoietic Stem Cell Fate In Vitro to Improve Hematopoietic Stem Cell Culture and Expansion and Their Hematopoietic Stem Cell Outcome
3D, three-dimensional; 2D, two-dimensional; HSC, hematopoietic stem cell; MSC, mesenchymal stem/stromal cell; UCB, umbilical cord blood; HSPC, hematopoietic stem and progenitor cell; ECM, extracellular matrix; AEMA, 2-aminoethyl methacrylate hydrochloride; PCL, polycaprolactone; PLGA, poly(lactic-co-glycolic acid); PES, polyethersulfone, SCF, stem cell factor; FL, Flk2/Flt3 ligand; TPO, thrombopoietin; IL, interleukin; sIL-6R, soluble IL-6 receptor; TCPB, tantalum-coated porous biomaterial; WSS, wall shear stress; EDA, ethylenediamine; LTC-IC, long-term culture-initiating cells; CFU, colony-forming unit; LTR-HSCs, long-term repopulating hematopoietic stem cells; PEG, poly(ethylene glycol); FN, fibronectin; BMSC, bone marrow-derived stromal/stem cell; HPC, hematopoietic progenitor cell; CB, cord blood.
Naturally derived polymers that include proteins, polysaccharides, amino acids, and apatite, and acellular materials composed of ECM and tissue-derived materials have been used for tissue regeneration with the possible advantage of biological recognition that might support the development of cells. Recently, decellularized ECM from a murine BM stromal cell line was used as a substrate for the ex vivo expansion of HSCs. 81 This substrate was found to play a major role in regulating the HSC expansion potential of the matrices. Lai et al. used decellularized ECM, derived from osteogenically differentiating MSCs, as an in vitro model to explore HSC niche. 82 A significantly more number of HSCs and HSC–MSC pairs per unit area had been discovered in the decellularized bone-like matrix, as compared with collagen scaffold.
In addition, incubation with extra-neutralizing antibody to bone morphogenetic protein 2 (BMP-2) ended in a considerably higher number of HSC per unit area than that without in the decellularized matrix. 82 However, this study did not address in vivo functionality of the cultured HSCs. The usefulness of a fully defined all-recombinant protein-based culture system has been investigated for reproducible in vitro HSC culture and its potential to specifically contribute to the identification of bona fide factors accountable for cell fate decision, such as in vitro self-renewal, maintenance, survival, expansion, quiescence, and differentiation of stem cell populations. 83 In this study, addition of hemopexin was found to give twofolds higher peripheral blood chimerism in irradiated recipient mice, showing improved maintenance of HSCs. Fujita et al. investigated that implantation of hydroxyapatite scaffolds in vivo partly recapitulates the HSC niche in vivo and maintains HSC properties. 84
Many combinations of stromal cells 85 and osteoblasts 86 that are constituents of the niche are reported to be able to improve the long-term survivability, migration, proliferation, differentiation, and maintenance of hematopoietic stem and progenitor cells (HSPCs) in vitro. A twofold increase in CD34+ cells was obtained using stromal cells and a three- to fourfold enhancement in human long-term culture-initiating cells units was obtained using osteoblasts. Similarly, several cytokines 87 have also been investigated for the ex vivo expansion of HSPCs. In addition, the synergistic effect between the three dimensionality of electropsun nanofiber scaffolds and surface-immobilized ECM fibronectin and the capacity of this scaffold for the expansion of HSPCs has been investigated. 88
The physicochemical properties of the biomaterial plays a critical role in supporting cell adhesion typically through sequestration of cell-secreted ECM proteins and nonspecific adsorption of proteins from the culture medium when there is no adhesive moiety present on the biomaterials, such as peptide conjugation and protein. The amount and conformation of cell-secreted ECM proteins and biomolecules at the cell–matrix interface offer the biological cues to regulate the HSC fate. This has prompted active research in the finding of various peptides and components of recombinant proteins to replace feeder layers in support of in vitro expansion of stem cells. Templated synthesis of protein-engineered biological constructs makes it effortless to produce families of functional biomaterial scaffolds from a very few peptides building blocks. These protein-engineered materials mimic numerous important properties of natural ECM and can be particularly designed and optimized for different tissue engineering and clinical application.
Recombinant protein-based biomaterials have the advantage of being able to combine essential properties, such as biocompatibility from natural component and tunability from synthetic component. Furthermore, these materials are synthesized with accurate molecular-level precision as they are recombinantly engineered and therefore, offer greater control over the protein structure and function that natural biomaterials fail to provide. In addition, as these materials are made out of proteins, a concern about the toxicity of materials is narrowing down. When compared with natural biomaterials these protein-engineered materials provide batch-to-batch consistency and represent a highly tunable class of materials important for stem cell long-term survival, maintenance, and fate. The scalability of HSC-based bioengineering strategies depends on the supply of a reproducible and extensive cell source and requires well-defined in vitro expansion of HSCs for clinical translation.
Recently, notch ligand-modified biomaterials, such as immobilized polystyrene microbeads covalently immobilized with delta-like ligand (DLL)-4 and polyacrylamide hydrogel coated with DLL-1 by layer-by-layer assembly technique, have been evaluated for their potential to regulate HSC differentiation toward T cell lineage.89,90 Using a hematopoietic cell line 32D, Cuchiara et al. confirmed that integration of RGDs and in combination of either stem cell factor or SDF-1α-conjugated poly(ethylene glycol) (PEG) hydrogel supports significantly increased cell adhesion. 91
In addition, hybrid functional polymers, composed of natural and synthetic polymer blends and their derivatives, have also been explored. For example, Ferreira et al. proposed a combined use of four different biodegradable biomaterials [polyester poly(lactic-co-glycolic acid) (PLGA), polycaprolactone (PCL), fibrin, collagen] with defined geometries, pore sizes, and fiber diameter with efficient cytokine supplementation for improved growth, adhesive property and expansion, and multilineage differentiation of CD34+ HSCs. 34 Typical functional groups, such as primary amine and carboxyl, were most widely used for the successful immobilization of biomolecules on the nanofiber surface. Substrate topographical cues also supported ex vivo proliferation, mobilization, and differentiation of stem cells.9,92 Chua et al. demonstrated that polyethersulfone nanofiber meshes functionalized with carboxylic, hydroxyl, and amino groups influenced HSPC adhesion, proliferation, and phenotype maintenance differently and contributed in HSPC expansion. 93
Soft versus rigid biomaterials
Apart from biochemical cues, mechanical or physical cues/signals of the native microenvironment, such as material elasticity, stiffness, and shear stress play a fundamental role in the differentiation of embryonic and adult stem cells.36,94–98 The importance of mechanical cues, such as matrix elasticity and shear stress, have been explored by Holst et al. 97 They engineered soft tropoelastin biomaterials with unique mechanical properties to examine the effects of elasticity on the maintenance of ex vivo HSPC populations. 97 They observed that growing mouse or human hematopoietic cells on a tropoelastin matrix resulted in a two- to threefold expansion of undifferentiated cells. Their results suggested that during development, HSPCs continuously sense and respond to the physical cues/signals supplied by their local niche environments. Lee-Thedieck et al. synthesized a biocompatible fibronectin-functionalized poly(ethylene glycol) diacrylate hydrogel substrate with tunable elasticity to test if the cultured HSCs in contact with the osteoblasts can actually sense the variations within the elasticity of the underlying matrix. 33 In the presence of chemoattractant (SDF-1α) on hard (>38 kPa) gels, they observed improved HSC adhesion and faster migration compared with soft (≤20 kPa) gels, indicating the importance of matrix elasticity on HSC expansion.
Choi and Harley showed that collective effect of matrix elasticity along with collagen density resulted in increase in the shape, spreading, and cytoskeletal organization of HSCs. 99 For ex vivo expansion of HSCs, optimal modulus has been reported in the range of 0.8 kPa 100 to 30 kPa 101 similar to that of the extracted BM, where varied range of stiffness was found along with vascular and osteoblastic niche. Recent finding of Choi and Harley's work indicated that matrix biophysical properties (matrix-associated stiffness) and cell–matrix (adhesive-associate protein) interactions are critical design considerations in biomaterial development to guide in vitro HSC performance. 102 They showed that polyacrylamide hydrogel-based matrix corresponding to stiffer (44 kPa, fibronectin-coated) endosteal region fostered maintenance of primitive myeloid progenitors, whereas those corresponding to the soft (3.7 kPa, laminin-coated) vascular region preferred differentiation predominantly toward erythroid lineages. However, how substrate-associated elasticity could play a role in the niche of the HSC itself, where HSCs proliferate naturally, remains an open question.
Substrate architecture and geometry
Collectively, matrix physical properties (stiffness, softness, roughness, and hydrophilicity), microstructure (porosity, pore shape, pore size, and interconnectivity), and geometry (porous biomimetic scaffold, hydrogel, and nanofiber) have a significant influence on the HSC performance. An essential part of signaling control for HSC long-term survival, proliferation, and differentiation is the interaction between HSCs and the components of the ECM. A range of substrates has been evaluated for HSC expansion based on pore size, shape, and their geometry. Recently, highly porous and interconnected scaffolds have been adapted as culture platform for HSC expansion to mimic native tissue architecture and BM microenvironment.35,103,104 Hematopoietic differentiation of embryonic stem cells (ESCs) depends heavily on physical characteristics of scaffolds like pore size, polymer concentration, and matrix mechanical stiffness. 105 It has been shown that smaller pore size and higher polymer concentration increases hematopoietic differentiation of ESCs. However, it has not been shown that selective changes in matrix physical properties (pore size or stiffness) influences HSC adhesion and migration. Further selective expansion of HSCs has been observed on fibrin scaffold, when a broad range of matrices such as PLGA, PCL, collagen, and fibrin is used. 34
Many researchers have explored cell encapsulating hydrogel-based platforms for HSC niche engineering. Small changes in hydrogel stiffness, density, composition, and orientation affect cell performance and phenotype. Hydrogel not only represents the physical and mechanical complexities of the stem cell niches 106 but also the encapsulating cells within it offers a realistic 3D microenvironment that supports many cellular mechanisms like cell adhesion, long-term survival, migration, 107 and cell–matrix mechano-interactions. 108 By mimicking few important features of the BM niche in optimized pullulan hydrogels supported delayed HSC differentiation into MKs and long-term HSC survivability. 109 Furthermore, RGD peptide-modified PEG hydrogel (fibronectin-mimicking hydrogel) greatly improved HSC proliferation. 110
Approaches in Designing Novel Biomaterials Systems with Unique Geometries and Morphologies
Emulsion templating
New findings and research methodologies in the design of biomaterials have also permitted guided differentiation of HSCs into specific lineages. The architecture and geometry of scaffolds used for tissue engineering is of critical importance. An interconnected porous structure is essential to motivate cell growth, nutrient and waste transport, and vascularization. 111 Some basic requirements for the design of polymer scaffolds have been broadly acknowledged. 112 The ideal 3D-engineered scaffolds should provide suitable porous and interconnected pore structures, pore sizes, acceptable mechanical properties, open surface area, biodegradability, and surface functional group chemistries supporting cell adhesion, proliferation, long-term survival, and differentiation. Conventional methods often produced scaffolds lacking interconnected porous structure and the required mechanical properties that are essential for optimal cell performance.
PolyHIPEs (high internal phase emulsion templated polymers), with their well-defined hierarchical interconnected porous structures (>75%) and highly adjustable void and interconnected window size, 113 can successfully support cell migration and growth when seeded in the matrix. 114 These polyHIPE materials have been used for human fibroblasts 115 and neurons derived from stem cells. 116 A hierarchical porous architecture can be introduced into scaffolds using water-in-oil (W/O) or oil-in-water (O/W) double emulsion technique, which are then subsequently seeded with stem cells and growth factors (Fig. 3). The thin, highly interconnected windows allowing exchange of nutrients and oxygen necessary for the cells to grow, migrate, and proliferate thus reduces the chances of cell necrosis. Recently, styrene-divinylbenzene 117 (synthetic) and gelatin-based 118 polyHIPE scaffolds have been successfully prepared and used for culturing hepatocytes in vitro. Biodegradable and hydrophilic vinyl-functionalized dextran polyHIPEs were shown to support neuronal migration, 119 whereas polyester-based polymers such as PCL 115 and PCL-polystyrene 120 have also been explored as tissue-engineered scaffolds. PLGA microspheres fabricated by W/O emulsion method with 3.4–5.1 μm pore size range could support ovarian cancer cells over 7 days culture when coated with type 1 collagen. 121 Drug-eluting porous and biodegradable ethylene glycol dimethacrylate microspheres were fabricated using a solvent-free multiple emulsion (W/O/W) HIPE method for controlled growth factor release of BMP-2. 122
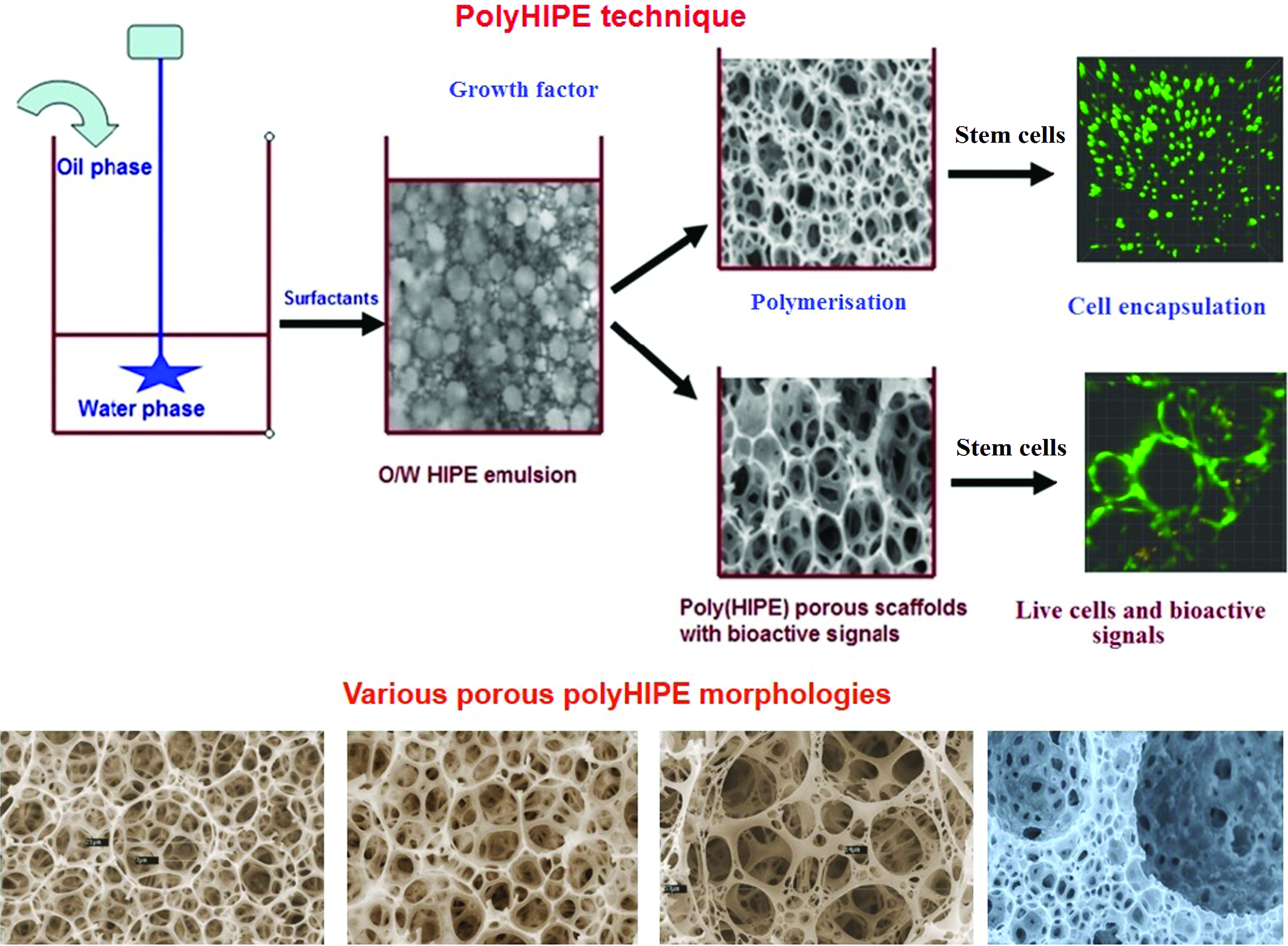
PolyHIPE technique and various HIPE scaffold morphologies obtained by HIPE polymerization. PolyHIPE, high internal phase emulsion templated polymer. Color images are available online.
Recently, 3D porous protein scaffold synthesized using poly-pickering HIPE method templated from O/W HIPEs costabilized by completely natural materials, gelatin, and its nanoparticles showed enhanced L929 cell adhesivity, growth, and functionality. 123 Ion-responsive methacrylate-modified alginate hydrogels with a well-defined hierarchical and unique porous structure were produced using O/W HIPE templating approach. To promote osteoblastic differentiation and viability of human MSCs, osteoconductive particles such as calcium phosphate nanoparticles and collagen-rich demineralized bone matrix particles were integrated into injectable polyHIPE. 124 This advanced emulsion templating approach allows the hierarchical porous structuring of scaffold in which micro and nanostructural arrangement is managed by W/O or O/W emulsion templating. It is possible to control the distribution of pore size and interconnectivity throughout the matrix by controlling window sizes using HIPE synthesis parameters, which are perhaps suitable for investigating the association between 3D porous surface and stem cell adhesion, migration, proliferation, and differentiation. The use of HIPE scaffolds for HSC expansion is still at a preliminary phase. Optimizing effect of highly porous nature of HIPE on HSC proliferation and differentiation will enhance the performance of the HIPE scaffolds toward HSC expansion.
3D cell printing
Traditional strategies mainly focused on soluble factors such as chemical and growth factors for predetermined HSC maintenance and fate in vitro. However, biotechnological and bioengineering advancements in biomaterials science have evidenced that other factors such as matrix geometry, topography, and stiffness can also contribute in regulating cell attachment, mobilization, long-term survival, proliferation, and differentiation of the HSCs. 125 In addition to these factors, a highly porous architecture of scaffold is another important factor that plays a significant role in cell infiltration and diffusion of nutrients (or waste materials) and oxygen.
Scaffold having defined mechanical, physical, and biochemical properties, and porous architectures for HSCs can be prepared by 3D printing. Polymeric biomaterial-based 3D cell printing offers stem cells the native tissue mimetic porous architecture and microenvironment that directs the functionality of the cell and improves cell viability. 3D cell printing has the potential to augment the regenerative efficacy of stem cell therapy and deliver stem cells to the target site while preserving their functions and long-term survivability. 126 Furthermore, 3D cell printing also facilitates the construction of a 3D complex native tissue with a predefined porous biological structure with mechanical and physical cues necessary for growing stem cells. In 3D cell printing system, bioinks refer to hydrogel (cell-encapsulating biomaterials) that contains stem cells and growth factors and cytokines that allow well-defined porous printed mass to be produced into a stable 3D form (Fig. 4), and also offers a cell matrix to mimic native tissue.127–130 As a result, the selection of bioinks can control a suitable microenvironment that offers biophysical and biochemical cues/signals for promoting stem cell functions to accelerate tissue formation.131,132 One of the foremost challenges within 3D cell bioprinting is getting the stem cells that make up the bioink to survive during the 3D printing process.

A diagram depicting bioprinting of HSCs. The bioink consists of biomaterials mixed with live cells and additives (growth factors and signaling molecules), which are printed to develop 3D tissue structures. Color images are available online.
To maintain cell integrity and long-term cell survivability within the cell-printed scaffold, it is necessary that bioink should have some biological (biocompatibility), mechanical (structural integrity), and rheological (viscoelasticity) requirements during the resuspension and ejection through the print head for delivery on collector. In addition, it should also be able to provide an appropriate environment for cell growth and function inside the printed 3D construct. Natural polymers such as collagen, fibrin, and gelatin hyaluronic acid have been explored for cell printing bioink that provides a natural-like environment for the printed cell.133–139 No studies were conducted to investigate the effect of this 3D cell-printing technique on the maintenance and expansion of printed HSCs. It is still not very clear how the HSCs in the 3D cell-printed scaffolds would attach, grow, migrate, and differentiate and communicate with the host tissue during hematopoiesis.
Electrospun nanofibers
HSCs reside in a complex BM ECM, which provides structural, mechanical, biological, biochemical and biophysical signals/cues that regulate stem cells' functions such as adhesion, self-renewal, migration, proliferation, and differentiation. It is thought that not only local growth factor and direct cell–cell interactions manage the balance between expansion and differentiation of HSCs, but also that the geometry of substrates plays a significant role in maintaining HSCs. Both matrix topographical and biochemical cues could work synergistically with the addition of cytokines to improve the ex vivo expansion of HSPCs. Interactions (cell–cell and cell–matrix) among HSCs and their surrounding niche microenvironment are essential for signaling control of HSC maintenance and functionality. 140 The lack of cell–matrix and cell–cell interactions is one of the factors responsible for the failure to maintain HSC's ability to self-renew and differentiate during in vitro culture.141–143 To develop 3D models of hematopoiesis, it is necessary to apply electrospinning techniques to formulate an artificial nanofibrous matrixwhich most closely resembles the native ECM (Fig. 5A).

Electrospinning has emerged as an enormously promising technique which offers numerous advantages such as fabrication of nanofiber scaffolds resembling the fibrillar structures of ECM, availability of large surface areas, ease of functionalization, and tunable mechanical and biological properties, 144 all of which may lead to enhancements in the ability to provide a true biomimetic microenvironment for HSCs maintenance. A wide range of flexible polymers can be electrospun, including natural, synthetic, semisynthetic, composite, or co-polymers integrating these features. Recently, modified electrospinning technique also demonstrated electrospinning of unspinnable polymers in core-sheath, triaxial/multichannel and emulsified nanofiber formulations (Fig. 5A), which makes it a superior technique out of all the methods applied so far. 144
Electrospinning can produce nanofibers of diameter in the range 3–1000 nm. In addition, wide-ranging pore sizes make these matrices suitable for HSC expansion when cultured alone or cocultured with MSCs by supplementing nanofibers with growth factors and conjugating them with cell adhesive peptides (Fig. 5B). Moreover, aligned architecture in 3D morphology can be easily produced using electrospinning. The understanding of structural and molecular constituents of HSC niche and their corresponding functions within 3D microenvironment is crucial for the development of successful HSC manipulation methodologies.
Interpenetrating network hydrogels
Single network biomaterials based on hydrogel have traditionally suffered severe constraints because of limited control of physicochemical and biological properties, impeding their usefulness in tissue-engineering applications. Making 3D microenvironment with a single network hydrogel (highly hydrated polymeric chain, with structural and mechanical properties like native tissue) has advanced enormously in the most recent decade. These artificial microenvironments can be precisely customized to mimic native tissue-like structures in vitro. However, small alteration in material composition would often change mechanical properties such as substrate stiffness and elasticity, which subsequently affect cell performance. To overcome these limitations of single-network hydrogel, an interpenetrating network (IPN) hydrogel-based biomaterial is designed for guiding cell performance with multiple functionalities.145–147 An IPN is a mixture of at least two polymeric networks (natural or synthetic) possessing distinct characteristics, produced by a two-step process involving diffusion of a second network monomer in a presynthesized network, followed by UV crosslinking (Fig. 6).148–150 Physical characteristics of material (elasticity) and biological cues or signals play an imperative role in guiding hematopoiesis. It is possible to use an IPN hydrogel platform with independently adjustable physicochemical and biological properties as a 3D biomimetic cell niche for HSCs. Moreover, this technique enables the simultaneous and independent formation of physicochemical and biological networks that do not harm the viability of cells.145,151

Steps involved in interpenetrating network hydrogel process. Cells and growth factors can be incorporated during the formation of first network. Color images are available online.
This versatile IPN biomaterial platform can create multifaceted patterns of niche-inspired signals to study the effects of physicochemical and biological cues/signals and BM niche cell paracrine signaling on HSC fate. An IPN could be an ideal platform to amalgamate the numerous components of the HSC niche and offer flexibility in manipulating the consequent interactions that dictate the performance. It has been reported that culturing HSCs (direct contact or noncontact) with other type of cells residing in the BM environment augmented the proliferation capability of the repopulating HSCs.87,152–155 The development of double-network hydrogel having heterotypic microenvironments containing one or more niche cells and HSCs will enable the coculture of HSCs with various niche constituents found within the native niche.
Future Challenges, Opportunities, and Concluding Remarks
Recent advances in materials science, bioengineering, and biotechnology facilitates microscale and nanoscale details to mimic the multifaceted HSC niche microenvironments. The rate at which biomaterials approaches are applied to address issues in stem cell biology ensures new insights into the mechanistic regulation of HSC fate. However, although there are plenty of resourceful biomaterial platforms available to examine the effect of the biophysical and biochemical properties of HSC niches, these multifunctional biomaterial-based approaches have just started to be used to guide HSC fate. Collective roles of material scientists/engineers, cell biologists, and biotechnologists are important in addressing the key biological intricacies and for nurturing interdisciplinary translational level of HSC research. Although the importance of microenvironment in controlling HSC fate has been shown through in vitro experimental systems, such complex environments have not been possible to recapitulate in vitro. Consequently, the hunt for an experimental system that closely and precisely resembles in vivo counterpart of the HSC niche continues.
The approaches explored above represent a key advancement in the fabrication of 3D microenvironment-inspired cell culture systems. These emulsion templating, 3D cell printing, nanofiber and IPN hydrogel-based technologies, along with cell-instructive biomaterials, are expected to make exciting contributions to the HSC biology field. For example, an IPN hydrogel approach will enable researchers to consider a huge collection (library) of natural and/or synthetic biomaterials that could be used as potential platforms for culturing HSCs along with different kind of niche cells and their pertinent growth factors. The success of IPN has been efficiently demonstrated in bone engineering 156 and cartilage tissue engineering145–147,151 and it is only a matter of time that the verdict on its efficacy on the expansion of HSCs will be appreciated.
Footnotes
Acknowledgments
The authors thank Symbiosis Centre for Research and Innovation (SCRI), Symbiosis International (Deemed University), Lavale, Pune, India for providing financial support for this research work.
Disclosure Statement
No competing financial interests exist.