
Abstract
Biomaterial-guided in situ tissue regeneration uses biomaterials to stimulate and guide the body's endogenous, regenerative processes to drive functional tissue repair and regeneration. To be successful, cell migration into the biomaterials is essential, which requires angiogenesis to maintain cell viability. Neutrophils, the first cells responding to an implanted biomaterial, are now known to play an integral part in angiogenesis in multiple tissues and exhibit considerable potential for driving angiogenesis in the context of tissue regeneration. In terms of biomaterial-guided in situ tissue regeneration, harnessing the proangiogenic potential of the neutrophil through its robust secretion of matrix metalloproteinase 9 (MMP-9) may provide a mechanism to improve biomaterial performance by initiating matrix reprogramming. This review will discuss neutrophils as matrix reprogrammers and what is currently known about their ability to create a microenvironment that is more conducive for angiogenesis and tissue regeneration through the secretion of MMP-9. It will first review a set of ground-breaking studies in tumor biology and then present an overview of what is currently known about neutrophils and MMP-9 in biomaterial vascularization. Finally, it will conclude with potential strategies and considerations to engage neutrophils in biomaterial-guided angiogenesis and in situ tissue regeneration.
Impact statement
This review draws attention to a highly neglected topic in tissue engineering, the role of neutrophils in biomaterial-guided tissue regeneration and angiogenesis. Moreover, it highlights their abundant secretion of matrix metalloproteinase 9 (MMP-9) for matrix reprogramming, a topic with great potential yet to be vetted in the literature. It presents strategies and considerations for designing the next generation of immunomodulatory biomaterials. While there is literature discussing the overall role of neutrophils in angiogenesis, there are a limited number of review articles focused on this highly relevant topic in the context of biomaterial integration and tissue regeneration, making this a necessary and impactful article.
Introduction
The goal of biomaterial-guided in situ tissue regeneration is to design biomaterials that harness the body's endogenous, regenerative processes to drive functional tissue repair at sites of tissue loss, injury, or disease. Rather than develop viable replacement tissues in vitro, which is problematic due to cell sourcing issues, time constraints, and cost, in situ tissue engineering relies on instructive biomaterials that effectively control the host microenvironment and endogenous cells for functional tissue regeneration. 1 The key steps that must occur for successful in situ regeneration are rapid cell migration into the biomaterial, integration with the surrounding extracellular matrix (ECM), robust angiogenesis, and eventually neotissue formation. Therefore, the success of in situ tissue regeneration is largely dependent on the efficiency of the first step, biomaterial-guided cell migration.
One promising avenue to encourage rapid cell migration for in situ regeneration is matrix reprogramming, or a resetting of the ECM to a less dense state that is more conducive to cell migration, proliferation, and angiogenesis.2,3 Compared with fetal tissue, adult tissues are dense and hypocellular, reducing their healing capacity. 4 Through matrix reprogramming, the physical impediments to cell migration and proliferation are removed while ECM-sequestered growth factors are simultaneously activated and released to support angiogenesis and tissue repair. Given the body's highly orchestrated innate immune response to implanted biomaterials, neutrophils can be considered a readily available and easily recruited pool of matrix reprogrammers to drive biomaterial-guided in situ regeneration. 5
Neutrophils, or polymorphonuclear leukocytes, are largely responsible for scavenging debris around a biomaterial through phagocytosis, generation of reactive oxygen species, and release of neutrophil extracellular traps (NETs). 6 Because neutrophil numbers peak within the first 24–48 h after biomaterial implantation, the field of tissue engineering has focused on the second wave of cells, predominantly macrophages, as the drivers of tissue regeneration. While macrophages are undoubtedly critical for biomaterial-guided angiogenesis and tissue repair, research in tumor biology suggests that the tissue engineering perspective is inherently limited by a lack of understanding of neutrophil dynamics. 7 Particularly, biologists have found that neutrophils contribute to the initiation of tumor vascularization. This ground-breaking work suggests that the field of tissue engineering should shift its focus from macrophages to neutrophils in the potential for biomaterial-guided angiogenesis.
This review highlights the significance of the neutrophil in biomaterial-guided in situ tissue regeneration beyond its classical antimicrobial actions. First, it will discuss matrix reprogramming as a strategy to enhance biomaterial integration and angiogenesis. Then, it will focus on the neutrophil's contribution to inflammation, biomaterial-guided matrix reprogramming, and angiogenesis through the secretion of matrix metalloproteinase 9 (MMP-9). Last, the most recent work in biomaterial–neutrophil interactions will be discussed and methods to engage our body's matrix reprogrammers for biomaterial-driven angiogenesis will be proposed. Ultimately, this review aims to bring attention to a largely unexplored area in tissue engineering that may hold the key to critical design considerations for enhanced biomaterial-guided in situ tissue regeneration.
Matrix Reprogramming
Matrix reprogramming is the concept of reverting the ECM back to a former state that is more conducive for functional tissue regeneration (Fig. 1).2,3 Similar to the idea of cell reprogramming, the objective is to erase the intrinsic hurdles that arise during tissue maturation and revert the tissue back to a state capable of rapid healing, as observed in fetal tissue repair. Fetal tissues heal with a hypercellular response resulting in complete healing without fibrotic tissue (Fig. 1A). This is partially attributed to unobstructed cell migration and proliferation due to a less dense ECM that is more rapidly remodeled as a result of low levels of MMP inhibitors.4,8 Alternatively, adult tissues normally heal with fibrosis because of increased ECM density and stiffness as a result of elevated collagen I content and low cellularity (Fig. 1B). 4

Fetal and adult wounds heal differently.
Therefore, in the context of in situ tissue engineering, which relies on cell migration from the surrounding tissue for integration, an advantageous approach is to design biomaterials that initiate matrix reprogramming (Fig. 1C). When the biomaterial is implanted, the notion is that the biomaterial will guide the inflammatory and innate immune responses to trigger a breakdown of the ECM. Reverting the ECM to a more fetal-like state of healing subsequently encourages cell migration and proliferation in a less dense environment. As a result, the degrading biomaterial is slowly replaced by functional tissue without fibrosis.
The premise of matrix reprogramming for enhanced tissue regeneration has been previously explored in the context of articular cartilage and meniscal repair.2,9 In a series of experiments aimed at improving articular cartilage repair, partially digested bovine cartilage explants were implanted subcutaneously to evaluate if partial digestion could enhance graft integration with surrounding tissue. 9 The authors found that compared with control grafts, the partially digested grafts had significantly higher cell counts at their borders, were nearly two times as integrated with the surrounding tissue as determined by matrix growth at the tissue–graft interface, and contained collagen type II intercalating with the wound surface. Similarly, Qu et al. investigated an electrospun poly(ethylene oxide) delivery system for collagenase to enhance meniscal repair.2,10 Through in vitro and in vivo experiments, they found that partial degradation of the meniscus at the wound interface significantly improved repair by accelerating cell migration, reducing ECM density, and enhancing graft integration.
Altogether, these examples represent a “worst-case scenario” for tissue engineers working with dense, avascular connective tissues with low cellularity. Success in such a challenging tissue environment through artificial manipulation of the ECM illustrates promise for matrix reprogramming in less-dense, vascularized tissues, which has yet to be explored. Therefore, designing biomaterials that instruct the patient's own cells, containing their own stores of degradative enzymes, to reprogram the ECM may bolster in situ tissue engineering outcomes. One excellent candidate that is readily available upon biomaterial implantation is the neutrophil.
Inspiration from Tumor Biology
In the tumor microenvironment, infiltrating inflammatory cells contribute significantly to the development of the vascular network, which supports the rapid proliferation of mutated cells and metastasis. 11 Tumor-associated macrophages have been considered the sole inflammatory maestro of tumor angiogenesis by secreting large amounts of proangiogenic factors, such as MMP-9, vascular endothelial growth factor (VEGF), interlueklin-8 (IL-8), epidermal growth factor, and basic fibroblast growth factor 2 to recruit endothelial cells. 12 However, nearly 10 years ago, several groups studying the role of neutrophils in tumor biology published a set of seminal studies illustrating that macrophages act in concert while the neutrophils turn on the angiogenic switch.
First, Ardi et al. identified that MMP-9 is uniquely released from neutrophils free of tissue inhibitor of metalloproteinase 1 (TIMP1), resulting in rapid activation in vitro and in vivo to induce angiogenesis.13,14 Using a modified angiogenic model, the authors showed that purified neutrophil MMP-9, which is secreted as a TIMP-free zymogen, undergoes efficient activation to its catalytic form. Contrastingly, macrophage MMP-9 is concurrently secreted with TIMP1, which inhibits some activity upon secretion. 15 They then showed that MMP-9 is a distinctly potent proangiogenic factor that exhibits angiogenic effects at subnanogram levels, resulting in a 5-fold increase in angiogenesis over vehicle controls as measured by the number of vessel sprouts in an in vivo collagen onplant model. This high level of potency is attributed to the neutrophil's inability to produce TIMPs, which reduces the odds of MMP-9 inhibition. 16 Thus, Ardi et al. were the first to provide biochemical evidence that neutrophils and their secretion of TIMP-free MMP-9 are central to tumor-induced angiogenesis.
Just 2 years after neutrophil-derived MMP-9 was found to be TIMP free and potently proangiogenic, another crucial discovery was made that described two distinct phenotypes of tumor-associated neutrophils. 17 Similar to macrophages, neutrophils were found to exist as either antitumoral N1s that suppress tumor growth or protumoral N2s that promote tumor growth. Polarized by transforming growth factor beta (TGF-β) within the tumor microenvironment, N2 neutrophils produce significant amounts of MMP-9 and VEGF, which together promote angiogenesis and tumor growth by degrading the ECM and reprogramming its components to support endothelial cell migration. Alternatively, N1 neutrophils, which result from TGF-β blockade, are characterized by an increased expression of tumor necrosis factor alpha (TNF-α) and a decreased production of VEGF and MMP-9, resulting in an antitumoral, antiangiogenic neutrophil phenotype. This work by Fridlender et al. demonstrated the previously unrecognized, phenotypic plasticity of neutrophils, suggesting a fundamental role for neutrophils in tumor vascularization through microenvironment polarization. 17
Strikingly, Deryugina et al. also highlighted the role of neutrophils in tumor-induced angiogenesis by showing that tumor-associated neutrophils secrete 40- to 50-fold more MMP-9 within 2 h compared with macrophages cultured over 48 h. 18 Using several different tumor models, the authors showed that this upregulated secretion of neutrophil-derived MMP-9 induced substantially greater in vivo angiogenesis compared with macrophages, which was attributed to the magnitude of secretion and its TIMP-free status.13,19 Finally, this study also showed that MMP-9 was essential for pericyte recruitment in the tumor microenvironment for vascular stability and cancer metastasis, with a lack of MMP-9 in MMP-9 knockout mice leading to a 50% reduction in pericyte density. These findings suggest that neutrophil-derived MMP-9 also functions as an important signaling molecule for vessel maturation and stabilization, although that is not the focus of this article. Ultimately, the magnitude of neutrophil-secreted MMP-9 and its effects on endothelial cell migration and pericyte recruitment demonstrate the impressive impact of neutrophils on tumor-related angiogenesis and highlight their immense importance in tissue development.
Taken together, these benchmark publications in tumor biology shifted the paradigm from macrophage-driven to neutrophil-driven tumor vascularization and suggest that the neutrophil may be pivotal for biomaterial-guided angiogenesis and tissue regeneration through similar mechanisms. Ideally, a biomaterial could be designed to harness the unique and abundant release of TIMP-free MMP-9 from neutrophils to drive angiogenesis. This may include achieving an ideal ratio between N1 and N2 neutrophils, analogous to regulating the ratio of proinflammatory M1 macrophages and anti-inflammatory M2 macrophages known to play an integral role in tissue regeneration.17,20,21 Furthermore, if the milieu of factors in the tumor microenvironment can regulate the neutrophil phenotype, similar factors may also regulate the neutrophil phenotype in the environment surrounding a biomaterial. The field has shown that macrophage phenotype can be regulated through biomaterial design, so it is highly likely this principle can be applied to neutrophils as well.20,22 Lastly, while initiating angiogenesis is critical for in situ tissue regeneration, vascular maturation and stabilization by pericytes are essential for long-term tissue viability. Thus, the secretion of neutrophil-derived MMP-9 may also contribute to tissue remodeling and maturation through signaling for pericyte recruitment and capillary stabilization. 18 Ultimately, these paradigm-shifting data in tumor biology provide inspiration to explore the role of neutrophils and their MMP-9 in in situ biomaterial-guided tissue regeneration.
Neutrophils in Biomaterial-Induced Inflammation
The implantation of a biomaterial (Fig. 2) results in tissue damage, which triggers inflammation, an innate immune response, and neutrophil recruitment. 5 Signals such as damage-associated molecular patterns (DAMPs), IL-8, and leukotriene B4 are released from the ECM and necrotic cells to recruit neutrophils from the circulation (Fig. 2A).23–25 These signals establish a chemotactic gradient that can be immobilized in the ECM to direct neutrophil migration. Aided by the increase in vascular permeability from the acute inflammatory response, there is a rapid amplification of phenotypically plastic neutrophils in the local microenvironment, referred to as neutrophil swarming (Fig. 2B). 26 Within 25–40 min, neutrophils undergo extravasation from the vasculature and swarm infected or sterile wounds before clustering around the site of tissue damage. Leukotriene B4, released from endothelial cells after exposure to DAMPs, is also secreted by neutrophils to mediate swarming in a feed-forward manner.26,27 Furthermore, vascular damage resulting from the surgical insult will lead to pooled blood, containing neutrophils, around the biomaterial. As a result, biomaterial implantation establishes a dynamic and complex microenvironment that is acutely infiltrated by neutrophils, eager to interact with and respond to the biomaterial, which provides an inherent opportunity to harness their angiogenic potential.
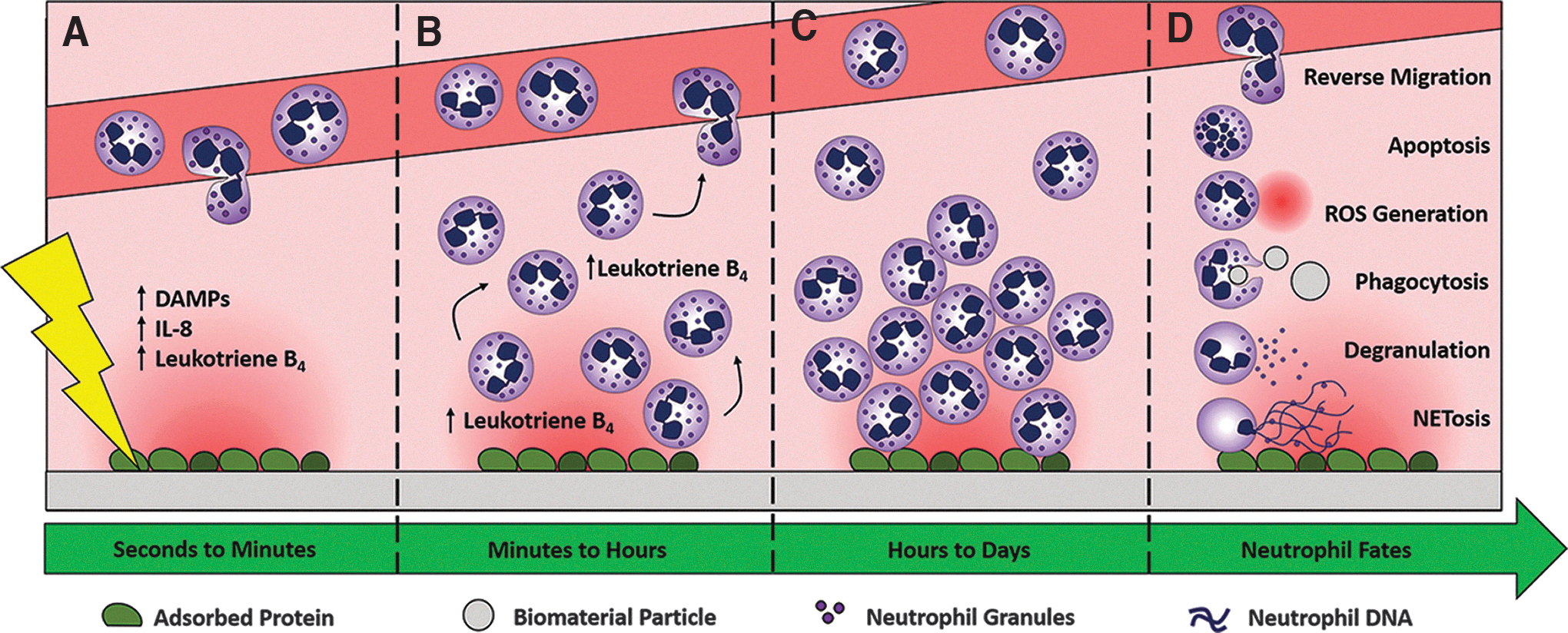
Implantation of a biomaterial triggers an innate immune response and neutrophil recruitment.
The swarming and clustering neutrophils either interact with the biomaterial directly through binding sites on the material surface or indirectly through binding sites on surface-adsorbed proteins (Fig. 2C). Natural polymer biomaterials, made from collagens, gelatin, and fibrinogen, are derived from the ECM and are inherently bioactive. They contain binding sites for integrins, which can induce outside-in signaling within neutrophils resulting in a multitude of effects. 28 In addition, synthetic materials present binding sites on their surface through the adsorption of blood plasma proteins. 29 Surface-adsorbed proteins may contain binding sites in their native conformation or may reveal cryptic binding sites through conformational alterations due to adsorption. 30 Direct or indirect interactions with the biomaterial results in a dynamic interplay between the material, the neutrophils, and the response.
The biomaterial-interacting neutrophils can respond through alternative pathways to modulate the inflammatory state of the tissue (Fig. 2D). Tools in their arsenal of responses include the production of reactive oxygen species through oxidative burst, the ability to clear pathogens and debris through phagocytosis, the capacity to release NETs through NETosis, and the exocytosis of granules through degranulation. Importantly, these processes, which have all been reviewed in terms of biomaterial–neutrophil interactions elsewhere, determine the outcome after biomaterial implantation by regulating the extent of inflammation and the rate of resolution. 31
Final resolution of the innate immune response and removal of neutrophils from the injury site are essential for biomaterial-guided in situ tissue regeneration. In the classical wound healing response, neutrophil recruitment peaks within the first 48 h and declines rapidly over the next several days while macrophages infiltrate the inflammatory site. This rapid response and subsequent decline suggest an important preconditioning function for neutrophils in the acute inflammatory phase. However, some studies have shown that neutrophils can persist for several days and weeks in the presence of inflammatory signals that decrease the levels of proapoptotic proteins.32–34 For example, neutrophils have been found in the peritoneal cavity 2 weeks after the implantation of alginate microcapsules, indicating their potential prolonged participation in the chronic response to a biomaterial, which may have positive or negative impacts on tissue regeneration. 35 Removal of neutrophils can occur through efferocytosis, where apoptotic neutrophils are cleared by macrophages, or can occur through reverse migration, where neutrophils return to the vasculature during resolution.36,37 The efferocytosis of apoptotic neutrophils by macrophages is an anti-inflammatory process that is essential for wound healing, implicating this as an important clearance mechanism to resolve inflammation.38,39 Ultimately, the biomaterial must be designed to direct the neutrophils to a restorative phenotype that promotes resolution of inflammation followed by functional tissue regeneration, both of which may be dependent on matrix reprogramming.
Neutrophils in Matrix Reprogramming
The neutrophil response to a biomaterial influences the potential for angiogenesis by altering the ECM characteristics in the local environment. Key determinants of the outcome are contained within the neutrophil granules, which are filled with degradative enzymes that can reprogram the ECM for sprouting angiogenesis and tissue development. Neutrophils contain primary, secondary, and tertiary granules that are prepackaged for rapid degranulation and release upon activation. 40 The contents of these granules have been well characterized, and each has been identified as a reservoir of matrix-degrading factors that can function independently or synergistically. 41
Several factors contained within the granules, such as neutrophil elastase (NE), neutrophil gelatinase-associated lipocalin (NGAL), and MMP-9, are uniquely important to biomaterials because they affect the potential for angiogenesis. NE, which is contained in the primary, azurophilic granules, is an abundant serine protease that targets all ECM components. 42 Distinct from other serine proteases, NE colocalizes with NETs, concentrating its catalytic activity to the immediate area around NETing neutrophils. 43 The secondary granules, also referred to as specific granules, contain NGAL. While it is not a matrix-degrading factor, NGAL complexes with MMP-9 and has been shown to exert enzyme-activating effects and protect MMP-9 from degradation.44,45 Finally, the tertiary granules contain MMP-9, which at a regulated level, may greatly influence biomaterial-guided angiogenesis through matrix reprogramming and signaling mechanisms.
While neutrophils do secrete proangiogenic growth factors and other proteases, MMP-9, or gelatinase B, has the most profound effects on angiogenesis due to its diverse enzymatic activity. 46 MMP-9 is secreted from neutrophils in three distinct forms after degranulation: a 92-kDa zymogen, a 120–130 kDa MMP-9/NGAL heterodimer, and a 200–300 kDa MMP-9 multimeter. 16 In terms of matrix reprogramming, the regions of interest of MMP-9 are the amino-terminal propeptide domain, a catalytic domain, and a hemopexin-like C-terminal domain that recognizes the substrate. 47 Activation of MMP-9 requires the removal of the propeptide domain, which signifies the transition from proMMP-9 to catalytically active MMP-9. This functions to protect the neutrophil from its incendiary cargo until degranulation. The removal of the 80 amino acid propeptide domain can be accomplished by proteases, such as plasmin and MMP-2, and NE, which also functions in promoting activation by degrading TIMP1.48–51
The catalytic domain of MMP-9 has promiscuous fibronectin type II repeats that facilitate collagen binding as well as binding of additional ECM components, such as gelatins, aggrecan, elastin, and vitronectin. 52 Neutrophils do not secrete a MMP-9/TIMP1 complex, and furthermore do not synthesize TIMP1, so the Zn-binding motif becomes accessible after removal of the propeptide domain. MMP-9 is also able to target other ligands, such as growth factors and cytokines, processing them from zymogens to enzymatically active forms. 53 For example, TGF-β and VEGF are stored in the ECM and can be activated and released upon ECM degradation by MMP-9.54,55
Lastly, the C-terminal hemopexin-like domain of MMP-9 regulates substrate binding to the catalytic domain and facilitates the formation of secreted MMP-9 multimers. 56 It is also this domain that binds TIMP1 to inhibit enzymatic activity. 57 Due to its structure and diverse activity, neutrophil-derived MMP-9 plays a major role in many tissue remodeling processes, including angiogenesis.
Neutrophils in Angiogenesis
Neutrophils are involved in physiological angiogenesis and tissue growth and development in multiple tissues and organs. 58 During endometrial development, neutrophils are the most abundant during the early growth of the decidua, decrease during menstruation, and are not present during tissue stability. 59 In a model of hindlimb ischemia, wild-type mice treated with granulocyte colony-stimulating factor facilitate revascularization and tissue regeneration through enhanced recruitment of neutrophils compared with untreated controls. 60 These examples demonstrate that there are endogenous mechanisms to induce neovascularization and overcome the ischemic load in developing tissue, driven by the neutrophil. Unfortunately, dysregulated and aberrant angiogenesis can also be driven by the neutrophil, leading to a number of pathologies, including tumorigenesis and psoriasis.61,62
The ECM plays a critical role in the regulation of neutrophil-driven angiogenesis by communicating continually and dynamically with the interacting cells through physical and chemical cues. 63 Cells probe the biomechanical properties of the ECM by applying a force to it and experiencing a force in return. Neutrophils in particular are known to migrate slowly but more persistently on stiff substrates, which through outside-in signaling through integrins is likely imparting their activation and polarization. 64 In addition to sensing the mechanical properties of the ECM, the ECM is communicating with the cells through its bound reservoir of growth factors. TGF-β is first secreted in an inactive form and stored in the ECM until MMP proteolysis. 54 Since it polarizes neutrophils to a protumoral phenotype, TGF-β released from the ECM that is degraded by biomaterial-confronting neutrophils may play a critical role in promoting tissue regeneration. 17 Additionally, IL-8, a strong neutrophil chemoattractant, is a proangiogenic factor in the tumor microenvironment that is upregulated through MMP-9 degradation of the ECM and secreted from activated neutrophils. 65 Normally, physiological angiogenesis is regulated with great accuracy, but in the case of biomaterial implantation, the balance between ECM production and degradation, in part mediated by the processing of growth factors, is often lost.
The angiogenic paradox is that the ECM is needed to keep cells from migrating sporadically to maintain homeostasis, yet the same restrictive matrix must be temporally degraded to guide endothelial cell sprouting during angiogenesis. A recent study by Trappmann et al. found that MMP-mediated ECM degradation regulates the multicellular, stand-like invasion of endothelial cells required for angiogenesis. 66 Moderately crosslinked hydrogels significantly enhanced the number of multicellular sprouts compared with lightly and highly crosslinked hydrogels. The lightly crosslinked hydrogels were too degradable and compliant while the highly crosslinked gels were resistant to degradation and stiff.
Although the authors were not evaluating neutrophil-derived MMP-9, this study demonstrates the significant role MMP-mediated degradation may play in overcoming the intrinsic ECM hurdles to in situ tissue repair. Moreover, this study also highlights the importance of regulating MMP-9 secretion to prevent a continuum of matrix destruction, which disrupts strand-like invasion and may result in chronic inflammation. In fact, elevated levels of MMP-9 predict poor wound healing in chronic pressure ulcers and diabetic foot ulcers as well as acute lung injury.67–69 As a double-edged sword, MMP-9 secretion should be temporally regulated with an initial upregulation for rapid matrix degradation to expedite angiogenesis, followed by downregulation to prevent chronic inflammation and poor wound healing.
Contrastingly, Aldabbous et al. recently examined the angiogenic potential of NETs, as opposed to MMP-9, in pulmonary hypertension and chronic thromboembolic pulmonary hypertension. 70 The authors found that NETs promoted angiogenesis in vitro in a two- and three-dimensional (3D) angiogenesis assay and in vivo in a Matrigel plug assay. By inhibiting Toll-like receptor 4 (TLR4) on the endothelial cells, tube formation and spheroid sprouting were abolished, indicating that activation of TLR4 is responsible for driving angiogenesis in their proinflammatory model.
While well designed, these experiments should not be taken out of context as NETs are known to induce endothelial cell dysfunction.71–73 The authors even state that when their experiments were performed with 10- to 30-fold higher NET concentrations, the NETs enhanced endothelial cell apoptosis. Moreover, NETs are associated with tissue fibrosis and impaired tissue healing.74–76 We have shown that the presence of NETs on the surface of electrospun biomaterials inhibits their integration with the surrounding tissue and results in the development of a fibrotic capsule. 77 Some NETs may have a proangiogenic effect in terms of biomaterial-guided angiogenesis by concentrating ECM remodeling factors. However, the general body of literature suggests that an overabundance will be detrimental to functional in situ tissue regeneration, suggesting that MMP-9 is an important proangiogenic factor for matrix remodeling.
Neutrophils in Biomaterial-Guided Angiogenesis and Tissue Regeneration
While there is strong evidence from other fields that neutrophils and their stores of MMP-9 are endogenous matrix reprogrammers initiating angiogenesis, tissue engineers working with biomaterials are just beginning to understand the significant contribution of neutrophils in biomaterial vascularization and in situ tissue regeneration capacity. In an exciting study, Christoffersson et al. evaluated the signals attracting neutrophils to avascular transplanted pancreatic islets and their subsequent physiological actions. 78 After transplanting the pancreatic islets into either the cremaster muscle or the abdominal external oblique muscle, the authors found that the pancreatic islets rapidly increased expression of VEGF-A, which resulted in the recruitment of proangiogenic neutrophils to the graft.
As a result, the VEGF-A-recruited neutrophils facilitated angiogenesis in the pancreatic islets and functional integration of the grafts. Contrasting to graft-recruited neutrophils, neutrophils recruited to a control inflammatory stimulus expressed 10 times less MMP-9. Moreover, revascularization of the pancreatic islets was severely impaired in MMP-9-deficient mice, which the authors attribute to the ECM remodeling effects of MMP-9. This insightful study by Christoffersson et al. was one of the first to illustrate the proangiogenic potential of neutrophils in biomaterial-guided angiogenesis. 78
In a follow-up study, Christoffersson et al. further investigated the distinct contributions and interactions of the proangiogenic neutrophils with the transplanted pancreatic islets using intravital microscopy. 79 First, the authors found there is an initial insurgence of neutrophils within the graft that rapidly diminishes over a 5-day period as vasculature and blood flow is restored. Next, the authors showed that while VEGF-A secretion from the islets aids in the recruitment of proangiogenic neutrophils, the graft itself was not the primary target. In fact, the neutrophils were recruited to angiogenic hotspots, or areas of endothelial sprouts, in the adjacent tissues.
Upon recruitment to the angiogenic hotspots, the neutrophils associate with the sprouting tip cells and seem to bridge the gap between new vessels. These data suggest that chemotactic signals from the endothelial cells override the hypoxic signals from the islets. All of these neutrophil data were juxtaposed with parallel macrophage experiments showing that macrophages accumulate in perivascular positions, indicating their role as support cells in developing vasculature. 80 Ultimately, this follow-up study highlights the elaborate contribution of neutrophils in biomaterial-guided angiogenesis.
Similar work by Lin et al. recently evaluated the ability of neutrophils to guide the formation of anastomoses with bioengineered microvessels. 81 In their work, the authors showed that the implantation of unassembled vascular networks resulted in anastomoses and patency, whereas assembled, mature vascular networks failed to connect with the host tissue. By depleting host neutrophils, the vascularization of the unassembled networks was abrogated, which suggests that the polarization of the neutrophil and its active engagement in the production of angiogenic factors is critical to the formation of anastomoses. The authors found that the endothelial cells in the unassembled grafts secreted nearly 10 times more IL-8 compared with the assembled grafts, which like the work by Christoffersson et al. suggests there is chemotactic recruitment of neutrophils toward angiogenic sites. 79 Altogether, these experiments demonstrate the angiogenic potential of the neutrophil for in situ-guided tissue regeneration in a different model and highlight the importance of biomaterial design in guiding these processes.
Herath et al. have also assessed the angiogenic potential of neutrophils in the context of bone tissue engineering. 82 Using a novel, triple cell coculture, the authors found that neutrophils upregulate the expression of angiogenic and osteogenic markers in human endothelial cells and osteoblasts, respectively, in a dose-dependent manner. Specifically, VEGF-A messenger RNA (mRNA) expression increased linearly with the number of neutrophils in culture. The authors also examined the potential temporal regulation of VEGF-A mRNA expression and found that it significantly increased from 4 to 48 h and decreased by 72 h. While they did not directly investigate the contribution of MMP-9, these data suggest that controlling the influx of neutrophils at an implanted biomaterial with the appropriate timing may be a potential strategy for improving in situ-guided regeneration.
Although each evaluated the role of the neutrophil in a slightly different context, the aforementioned studies all share a common theme: the neutrophil is able to reprogram the static ECM and contribute to biomaterial-guided angiogenesis. Dynamic manipulation of the wound environment for matrix reprogramming has previously been shown to increase the potential for guided tissue regeneration by significantly improving cell density at the wound margins and integration with the biomaterial.2,10,83 These previous implantable systems used the synthetic delivery of factors, like collagenase, to increase ECM porosity for accelerated cell migration. Alternatively, we propose focusing on the endogenous repair processes driven by neutrophils and their secretion of MMP-9. While the current literature has focused on cellularized, viable biomaterials, the same principles can likely be applied to and improve the performance of acellular biomaterials. Neutrophils are capable of promoting angiogenesis and tissue regeneration, and devising a strategy to utilize them may substantially improve biomaterial-guided in situ tissue regeneration.
Engaging Neutrophils in Biomaterial-Guided Matrix Reprogramming
Several levels of regulation could be targeted to engage neutrophils and their MMP-9 in biomaterial-guided matrix reprogramming (Fig. 3). The secretion of MMP-9 could be controlled at the intracellular level through MMP-9 synthesis and degranulation, at the extracellular level through the inhibition or activation of MMP-9 in the ECM, and at the level of cell recruitment. Each level could be targeted by engineering the properties of the biomaterial to elicit the appropriate immunomodulatory response. No matter the level of regulation, the overriding effect of the biomaterial should be to engage the neutrophils as drivers of matrix reprogramming through the optimal secretion of active MMP-9, resulting in a less dense and more fetal-like state of healing for functional tissue regeneration.

Potential levels of regulation to engage neutrophil MMP-9 in matrix reprogramming.
The first level of regulation is the intracellular level, where MMP-9 could be controlled at the level of synthesis and degranulation (Fig. 3A). MMP-9 synthesis is regulated at the level of transcription primarily by ERK1/2 in the MAPK kinase pathway.84,85 Activation of the pathway induces MMP-9 gene expression and could be used as a therapeutic target to regulate MMP-9 secretion.
In addition, integrin and TLR interactions can induce neutrophil degranulation and provide a second intracellular point of intervention. Mature neutrophils circulate with proMMP-9 packaged into secretory vesicles for rapid release. 86 Thus, engaging integrins (e.g., L-selectin and Mac-1) and TLRs (e.g., TLR2 and TLR4) could be used to regulate MMP-9 secretion.87–89
Various cytokines and chemokines binding to their receptors can also modulate neutrophil degranulation and further participate in autocrine and paracrine neutrophil feedback loops. As an example, neutralizing TNF-α suppresses MMP-9 secretion in a model of central nervous system tuberculosis. 84 All of these data suggest that MMP-9 secretion could be regulated at the intracellular level, through transcription or outside-in signaling, to increase the potential for biomaterial-guided angiogenesis.
Another level of MMP-9 regulation that could be targeted through biomaterial design involves the activation or inhibition of MMP-9 after secretion into the microenvironment (Fig. 3B). As previously discussed, MMP-9 is initially secreted as a zymogen that requires activation. 14 The availability of activators therefore affects the enzymatic potential of MMP-9. Multiple proteases are known to activate proMMP-9, including cathepsin K, MMP-2, MMP-3, and plasmin, which explains the rapid activation upon secretion.14,51,90,91 In addition, MMP-9 can be inhibited by TIMP1 before or after activation. Even without neutrophils producing it, TIMP1 could still be present in the microenvironment as it is constitutively expressed in many tissues. 92 Thus, devising a strategy to ensure efficient MMP-9 activation in the local microenvironment while minimizing or delaying inhibition may increase the potential for matrix reprogramming.
The last regulatory level that could be targeted to engage neutrophils in biomaterial-guided matrix reprogramming is neutrophil recruitment (Fig. 3C). The knockout of neutrophils is known to impair angiogenesis and tissue regeneration.60,81,93 Therefore, their recruitment to sites of inflammation is essential to wound healing, partly due to their delivery of growth factors and chemokines that promote the resolution of inflammation. Increasing the recruitment of a proangiogenic subset of neutrophils through VEGF-A secretion may be particularly beneficial for biomaterial-guided angiogenesis.78,94 Nonetheless, regulating angiogenesis at the level of recruitment may present additional challenges as the sustained recruitment of neutrophils and degradation of the ECM play a central role in chronic inflammation and poor wound healing.67,69,95 Due to multiple levels of regulation, several points of intervention exist for engineers to regulate neutrophil MMP-9 secretion to initiate matrix reprogramming.
In addition to regulating MMP-9 secretion to enhance matrix reprogramming, an immunomodulatory biomaterial would ideally direct balance in the polarization of confronting neutrophils to create a proregenerative microenvironment. Since it is known that the M1 to M2 phenotypic switch is necessary for tissue regeneration, we likely need a proportion of N1s for their proinflammatory effects and a proportion of N2s for proangiogenic effects. 96 Initially upregulating MMP-9 secretion from abundant proangiogenic N2 neutrophils, followed by their timely removal from the wound site to encourage resolution, may stimulate matrix reprogramming.
Furthermore, there is substantial crosstalk between neutrophils and the other cell types involved in wound healing and tissue regeneration. 31 Neutrophil–platelet interactions are central to thrombus formation and result in the release of NETs, which function as scaffolding for the developing thrombi. 97 After the initial influx of neutrophils, monocytes–macrophages invade the wound site to progress healing or fibrosis. 6 Signaling between neutrophils and macrophages is essential for cardiac repair after myocardial infarction with neutrophil-derived NGAL acting as the key inducer of highly phagocytic, proregenerative M2 macrophages. 98 Because NGAL and MMP-9 complex, which is thought to protect MMP-9 from proteolytic degradation, and closely correlate in amount, MMP-9 is likely acting in cooperation with NGAL and reprogramming the ECM to aid in repair. Our attention should be on understanding how neutrophils and their phenotypes set the stage for the ensuing response and the potential for biomaterial-guided in situ regeneration.
Importantly, nearly every aspect of the biomaterial design influences the progression of tissue repair, and likely influences the neutrophil response preceding it. For acellular electrospun templates, we have shown that the release of NETs is modulated by fiber diameter, pore diameter, and material composition.77,99 These same properties may also be regulating neutrophil polarization since they are known to regulate macrophage polarization. 7 Thus, architecture and surface chemistry are critically important material characteristics. In addition, the biomechanical properties of the biomaterial should be considered, not only for the purpose of withstanding the physiological forces upon implantation, but also for the purpose of facilitating neutrophil migration into the material to create cell highways for guided angiogenesis. Finally, the chemical signals given off by the biomaterial will greatly contribute to neutrophil recruitment and angiogenesis. Lin et al. illustrated the profound difference in neutrophil recruitment to engineered vascular networks and the formation of anastomoses with host vasculature due to the maturation state of the biomaterial. 81 Immature vascular networks were able to engage neutrophils and stimulate anastomoses whereas mature vascular networks failed completely. These data suggest that maturation of cellularized constructs ex vivo in a bioreactor may be detrimental to the success of the biomaterial. Instead, the human body and its own endogenous mechanisms may be the best bioreactor for biomaterial-guided angiogenesis and tissue regeneration.
Nevertheless, our current understanding of these neutrophil-driven mechanisms is limited in the context of tissue engineering, and more research is needed to evaluate neutrophil–biomaterial interactions. In vitro experiments using human peripheral blood neutrophils are thought be constrained by the neutrophil's short in vitro half-life, but culturing in more physiologically relevant, 3D microenvironments may delay apoptosis and prolong the neutrophil lifespan.77,82 Such studies may significantly enhance the ability to evaluate the neutrophil effector functions over the first critical hours following biomaterial implantation. Similarly, in vivo models using intravital microscopy, which have already substantially increased our understanding of the dynamic recruitment of neutrophils, may provide greater insight into complex crosstalk between cell types.26,79 Since we have only just begun to understand that neutrophils are engaged in matrix reprogramming and biomaterial-guided angiogenesis, the next phase of discovery will likely produce exciting results, implicating the neutrophil in in situ tissue regeneration.
Conclusion
In situ tissue regeneration has the potential to revolutionize the field of tissue engineering through the design of immunomodulatory biomaterials that harness the proangiogenic potential of neutrophils to drive matrix reprogramming. The current literature suggests the readily available neutrophil is of critical importance to wound healing in more than just the custodial duties previously assigned to it. Given that biomaterials are immediately bombarded by the body's highly orchestrated innate immune response upon implantation, we should utilize what physiology has provided as endogenous matrix reprogrammers. Studying the abundant, circulating neutrophils and the use of the body as the ideal bioreactor to vigorously drive in situ biomaterial-guided tissue regeneration will generate a new knowledge base to propel the field of tissue engineering into the next generation of biomaterials.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Science Foundation Graduate Research Fellowship Program under Grant No. 1451514.