
Abstract
The clinical relevance of perichondrium was recognized more than a century ago. In children and adolescents, perichondrium is essential for the formation and growth of the cartilaginous part of craniofacial features and must be considered during reconstructive surgery in the head and neck area. Also in adults, perichondrium must be preserved during surgical intervention for adequate postoperative healing and cartilage maintenance. Furthermore, the regenerative function of perichondrium in the ribs enables the harvesting of the rib cartilage tissue for reconstruction of craniofacial features. With the advancement of tissue engineering, renewed attention has been focused on the perichondrium, because without this crucial tissue, the function of cartilage engineered for craniofacial reconstruction is incomplete and may not be suitable for long-term reconstructive goals. Furthermore, interest in the perichondrium was revived owing to its possible role as a microenvironment containing stem and progenitor cells. Here we will revisit seminal studies on the perichondrium and review the current literature to provide a holistic perspective on the importance of this tissue in the context of regenerative medicine. We will also highlight the functional significance of perichondrium for cartilage tissue engineering.
Impact statement
All adult cartilage tissues, with the exception of articular and fibrocartilage, are lined by a stratified tissue called the perichondrium. The perichondrium contributes to growth, structural stability, and regeneration and maintenance of the organ, but the cellular mechanisms underlying these processes are not well understood. This review provides a comprehensive summary of past and recent studies on perichondrium from the vantage point of tissue engineering and regenerative medicine. Of particular relevance is the evidence that perichondrium might contain chondrogenic progenitor cells. Cartilage tissue engineering holds great promise for novel treatments of craniofacial defects, and a better understanding of the function and structure of the perichondrium could contribute to improved therapies for head and neck reconstructive surgery and beyond.
Introduction
The perichondrium is possibly among the first tissues to attract clinical interest for its regenerative potential, with initial reports on its role in cartilage regeneration made as early as the 19th century.1,2 At that time, it had already attracted the attention of researchers and medical practitioners who believed that cartilage itself did not have the capacity for healing, but that damaged cartilage could be regenerated from the overlying perichondrium. 2 In the early 1970s, the interest in the perichondrium was renewed thanks to a series of studies performed by a group of researchers at the University Hospital of Uppsala in Sweden. 3 These studies were motivated by the hypothesis that a characteristic disfiguration observed in wrestlers called “cauliflower ear” (Fig. 1) occurred owing to the formation of neocartilage between the perichondrium and ear cartilage when the two were detached by a posttraumatic hematoma. 3 The hypothesis was confirmed experimentally in 2-month-old rabbits, when a blood clot placed between the perichondrium and auricular cartilage led to the generation of ectopic cartilage, whereas a clot placed between the skin and perichondrium did not. 4 Retraction of the detached perichondrium caused the cartilage to fold, thus forming a characteristic deformation over the hematoma.
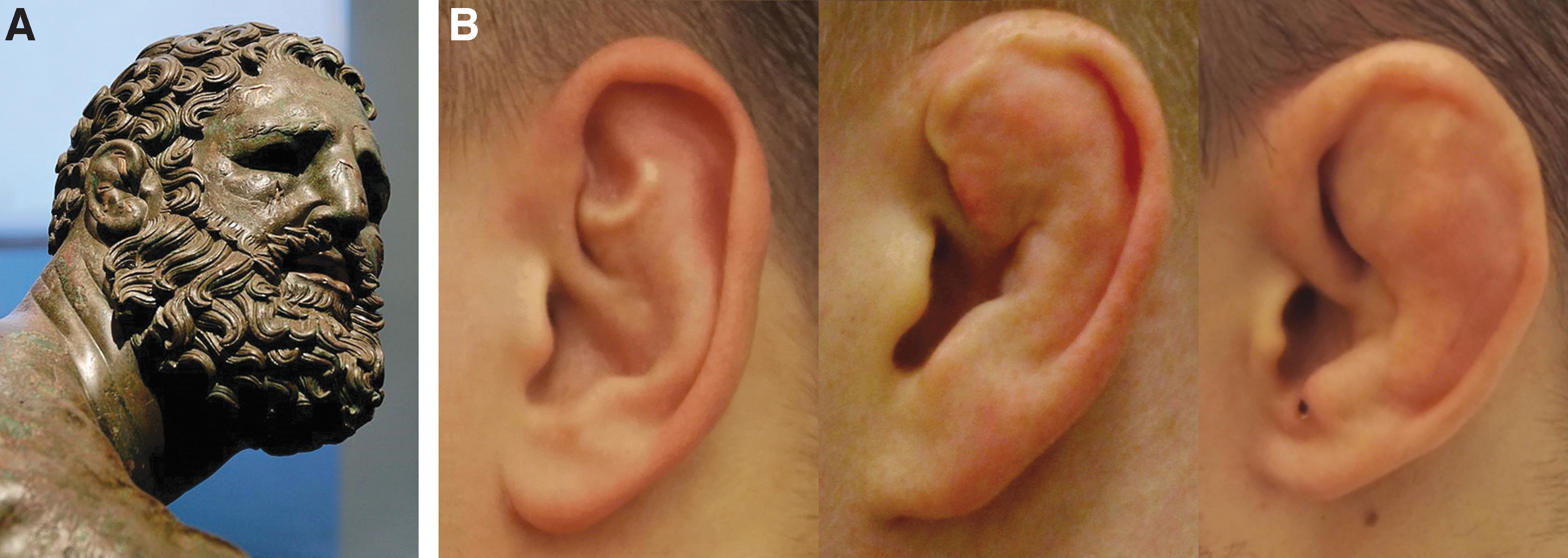
Cauliflower ear.
In a series of follow-up experiments, defects created in rabbit ears regenerated when perichondrium was preserved but not when cartilage was excised together with the adjacent perichondrium. 5 Modifications of the operation, such as increasing the defect size or fixing the defect edges with a metal ring were made to ensure that neocartilage was indeed originating from the perichondrium and not from the sides of the resected cartilage. Consistent with the “cauliflower ear” hypothesis, placing a blood clot between the cartilage defect and perichondrium enhanced cartilage regeneration.
After these initial observations, a number of studies were performed to investigate using perichondrial grafts for reconstructive purposes. Autologous perichondrium from ear and rib was transplanted into remote anatomical sites, aiming to show the potential of perichondrium grafts to form ectopic cartilage6,7 and to reconstruct articular cartilage defects in animal models.8,9 In an exemplary study in rabbits, 10 regeneration of functional articular cartilage from rib perichondrial grafts was observed over the course of 6 weeks, with no signs of resorption after a 3-month follow-up time. To transplant the graft, the researchers first created a deep defect down to the subchondral bone in the articular cartilage of femur condyles. The rib perichondrial grafts were then fixed on the subchondral bone using a fibrin glue, with the chondral surface of the graft facing the space of the joint. The knee joints were immobilized in a semi-flexed position for 13 days using a K-wire, thus keeping the graft pressed against the bone surface in the defect.
The authors reported in detail on the dynamics of the regenerative process, observing an active proliferation of “mesenchyme-like” cells originating from the perichondrium and forming a thick layer of matrix. The fibrous part of the perichondrium disappeared by the 9th day and the fibrin glue by the 16th day after the surgery. Of note, neither capillary perforations nor vascularization of the graft from the bone surface were observed. After 6 weeks, the newly formed tissue had transformed to resemble the appearance of the surrounding cartilage. The neocartilage was, however, thicker, had an uneven surface, and showed higher basophilic staining. It did not form a connection with the surrounding cartilage tissue, remaining separated from it by fibrous connective tissue. In a small group of animals from the same experiment, the cartilage graft was placed with the chondrogenic side facing the defect and the fibrous side facing the joint surface. Here too, proliferation and matrix deposition by the “mesenchyme-like” cells were observed, with eventual resorption of the fibrin glue and disappearance of the fibrous layer of the perichondrium. The authors hypothesized that “mesenchyme-like” cells were fibroblasts, which proliferated and differentiated into chondroblasts under the influence of factors in the synovial fluid. The presumed stimulating effect of synovial fluid was supported by previous observations, where transplantation of free perichondrial grafts into the joint space without fixation caused the formation of a free cartilaginous body.8,11
Overall, transplantation of perichondrial grafts has been performed in several animal species and at various anatomical sites.1,12 After the initial confirmation of the effectiveness of perichondrial grafts in animal models, a number of further studies reported the use of perichondrium to treat various cartilage-associated conditions in humans, with mixed success. These included, but were not limited to, arthroplasty of hand and finger joints,13–15 knees, 16 wrist, 17 and the temporomandibular joint. 18 Despite the success of these experiments and initial clinical use, properties of the newly produced cartilage tissue generated from perichondrium were considered unpredictable and varied in both quality and quantity, often displaying heterogeneity in histologic appearance and the amounts of extracellular matrix (ECM) components (e.g., chondroitin sulfate).10,19 Overall, however, a prevalent view had developed that perichondrium could facilitate and was essential for cartilage regeneration both in vivo and in vitro, and had the clinical potential to treat various cartilage-associated diseases and traumas.
The Structure of Perichondrium—Layers and Cellular Composition
During the several decades of heightened interest in perichondrium, some authors attempted to characterize basic perichondrial structures and pinpoint the area from which regeneration occurred. Initially, studies divided perichondrium into three zones chiefly based on the orientation of ECM fibers and resident cells, by the use of polarized light microscopy and visual observation. The first, or the fibrous, zone was characterized by the horizontal arrangement of collagen fibers (Fig. 2A, B). Most authors distinguished the second zone by the presence of elongated cells, arranged parallel to the fibers in the fibrous perichondrium.1,10,20 The proliferative process during neocartilage formation was thought to take place in this zone, as evidenced by the dynamic observations of transplanted perichondrial grafts and autoradiography using tritiated thymidine.1,9,10,20 The third and final zone was defined by the gradual shift of ECM fiber orientation in a vertical direction, where cells would transition from elongated to a more roundish and ellipsoid shape, acquiring a characteristic chondrocyte phenotype. Different authors used different terminology to refer to these three zones—Ohlsen and Widenfalk referred to them as fibrous, median, and transition zones, 10 whereas Bruns et al. used the terms fibrous, proliferation, and transition zones. 1 Yet another study divided perichondrium into outer, intermediate, and inner layers based on the same morphological characteristics. 20

Zones and cellular composition of the perichondrium.
By the 1990s, however, the zonal characterization of perichondrium was redefined altogether, and a more definitive terminology was introduced in an ultrastructural study with the use of electron microscopy.
21
The study examined the perichondrium of several different mammalian cartilage tissues, including auricle of mouse, rat, and rabbit; tracheal cartilage of rat and rabbit; rib and xiphoid cartilage of rabbit; and human nasal cartilage (Fig. 3A

The structure of the perichondrium.
Another remarkable feature of this study was the detailed description of the cellular composition of perichondrium. Especially interesting was the observation of extremely flat cells located between fibrous perichondrium and the overlying connective tissue in the rat auricle (Fig. 3B). Unlike fibroblasts in the fibrous perichondrium, these cells possessed long interconnected cytoplasmic processes, which would occasionally connect with the neighboring cells, thereby forming a layer. In addition, the cells displayed a unipolar arrangement of vesicles and electron-dense structures distributed toward the perichondrial side and a smooth and regular surface facing the connective tissue. Because of these distinguishing features, this previously unknown cell type was termed perichondrial lining cell (PLC).
Overall, Bairati et al. defined the following structural parts of perichondrium—(1) the external, or the vascularized loose connective tissue (LCT) layer, (2) the central layer, or the fibrous zone, and, (3) the internal layer, or the subchondral zone where perichondrium fuses with cartilage ECM (Fig. 2A). The authors emphasized that they could not detect any stem cells in the adult perichondrium, while acknowledging that stem cells might be present during embryonic development, residing in the subchondral zone.21,23 The cellular composition of the adult perichondrium according to Bairati et al. was reduced to perichondrial fibrocytes (referred to as perichondrocytes throughout the rest of this review) and, in auricle and trachea, PLCs (Fig. 3B). The zonal characterization and the structural framework of perichondrium delineated in this study had not been challenged and is still the existing paradigm.
Metabolic and Homeostatic Function of Perichondrium
Perichondrium is an intermediate tissue between cartilage and the overlying anatomical structures, such as skin in the auricle or the mucosa in the nasal septum. Unlike the underlying cartilage, perichondrium is vascularized and its proposed role in providing nutrients to the cartilage is well known and clinically relevant.3,24,25 The presence of blood vessels was shown in the LCT of perichondrium 21 (Fig. 2A, B). Of interest, PLCs were seen to occasionally envelop blood vessels at the perichondrial surface with their cytoplasmic processes (Fig. 2A). Finding a morphological resemblance of PLCs with the bone lining cells (Fig. 3B), Bairati et al. proposed a metabolic barrier as their possible function and suggested that PLCs might be necessary for the prevention of cartilage regression and calcification. This assumption was supported by the observation that PLCs were only found in cartilage types where calcification typically does not occur, namely in auricular and tracheal cartilage. In contrast, costal cartilage and the nasal septum, which are prone to calcification, do not contain the PLC layer. Indeed, characteristics of PLCs, such as contact with blood vessels and distribution of vesicles toward the cartilaginous side, are suggestive that perichondrium might be involved in the regulation of the metabolism of the cartilage tissue. The presence of vesicular structures toward the perichondrial side could be indicative of the active transport of metabolites to the underlying tissues. Furthermore, Bairati et al. implied that appositional growth happens by proliferation of chondrocytes beneath the perichondrium and put into question whether perichondrium could be the source of new cells for cartilage growth. In their view, the sole role of perichondrium in appositional growth is to provide the factors for proliferation of the underlying chondrocytes.
Paradoxically, the function of PLCs has not been studied in any further detail since the initial description of this cell type, as far as the authors of this review know. This could be because of the fact that research interest in recent years has shifted mostly toward the search for stem and progenitor cells in the perichondrium, as described in detail in the following section. Furthermore, a detailed exploration of cellular components of perichondrium is limited by the scarce availability of the tissue. Moreover, in their report on PLC, the authors noted that integrity of PLC layer was sensitive to fixation procedure, 21 which could be another reason why this layer has been overlooked. Whatever the reason, follow-up studies exploring PLCs and confirming the presence of this type of cells in humans remains to be performed.
The role of perichondrial fibroblasts in wound healing and regeneration of damaged cartilage tissue is currently unclear. To address this question, Duynstee et al. 26 studied wound healing in cartilage by resecting a portion of the rabbit auricle and culturing the tissue in vitro. In different modifications of the wound-healing models, the authors either preserved the perichondrium of the excised tissue, removed only the superficial fibrous part, or removed the perichondrium completely. Interestingly, in the sample where the complete perichondrium was preserved, the cartilage “wound” was overgrown by the fibrous tissue, which hampered further cartilage regeneration. When only the superficial part of the perichondrium was removed, a new cartilage tissue would form. In contrast, when perichondrium was completely removed, cartilage alone would show no signs of regeneration. It is not completely clear from the article whether by the superficial portion of the perichondrium authors were actually referring to the LCT, which covers the fibrous perichondrium. In any case, this interesting observation suggests that the matrix and/or fibroblasts in different layers of perichondrium might be characteristically and functionally different—fibroblasts in the fibrous part might stimulate production of new cartilage tissue, whereas fibroblasts in the LCT merely produce fibrous tissue.
In the same study, high amounts of transforming growth factor (TGF)-β in the perichondrium were reported. Remarkably, perichondrium is also rich in fibronectin (Fig. 2B). Interaction of fibroblasts with fibronectin is crucial in the regulation of TGF-β availability during wound healing and fibrosis27–29 and could be one of the mechanisms by which fibroblasts in the perichondrium regulate proliferation and maturation of the adjacent chondrocytes. Furthermore, whereas only a scarce presence of PG aggregates has been reported in the perichondrium, 21 these might also contribute to the bioavailability of the growth factors such as TGF-β in the fibrous zone,30,31 especially at the cartilage–perichondrium interface, where PG were described to occasionally protrude into the perichondrium. 21 However, the existence and nature of such ECM-growth factor interactions in the perichondrium remain a subject of further study.
Perichondrium as a Stem Cell Niche
The presence of chondroprogenitors has been reported at specific anatomical sites in different types of cartilage, such as in the surface zone of articular cartilage32–34 and on the groove of Ranvier in the knee joint 35 and intervertebral disc. 36 Some authors have defined these regions as stem cell niches.35,36 Furthermore, in some cases, chondroprogenitors were reported to fall under the minimal accepted criteria defining mesenchymal stromal cells (MSCs) and are thereby considered MSCs (reviewed in Vinod et al. 37 and Candela et al. 38 ). This review will mainly focus on presumed progenitors in the perichondrium of the auricle.
With increasing knowledge and refinement of research methods on stem cells, several groups have attempted to identify stem and progenitor cell population in the perichondrium. Various studies have tested multipotency, clonogenic potential, and MSC marker expression of perichondrocytes. A few researchers have attempted to localize stem and progenitor cells in the perichondrium with the use of DNA label retention assays. The premise of this method is based on the observation that adult stem cells are often quiescent, divide infrequently to prevent stem cell pool exhaustion, and will thereby retain a DNA label, such as bromodeoxyuridine (BrdU). 39 Actively proliferating cells, in contrast, will dilute the label over time. Togo et al. reported the presence of a scarce number of label-retaining cells (LRCs) in the rabbit ear at the junction between perichondrium and cartilage, 4 weeks after injecting 4-week-old animals with BrdU 40 (Fig. 4A). Kobayashi et al. injected pregnant mice with BrdU and traced the progeny for up to 24 and 48 weeks into adulthood, finding LRCs in the perichondrium but not in the cartilage of the adult animals 41 (Fig. 4C). Similarly, Srour et al. reported retention of the DNA label in costal perichondrium of neonatal mice for up to 8 months 42 (Fig. 4B). The regrowth of the resected costal cartilage originating from the perichondrium shown in this study further suggested the presence of putative stem cell population in the rib perichondrium. There is therefore evidence of the existence of LRCs in perichondrium in more than one species and at more than one anatomical site. It should be noted, however, that although suggestive of stem cell phenotype, label retention is not sufficient for stem cell identification, because some fully differentiated cells might also divide infrequently and retain DNA labels. 39 Furthermore, in contrast to dormancy, the active proliferation of some types of stem cells (e.g., in the gastrointestinal tract) has also been demonstrated.43,44

LRCs at the perichondrium–cartilage interface.
Most studies on perichondrocytes also suggest higher proliferation, in addition to the colony-forming capacity (CFC) of these cells in contrast to chondrocytes as an indication of the presence of progenitors in the perichondrium. Particularly, high CFC in perichondrocytes derived from rabbit, 40 mouse, 41 human, 45 and porcine 46 ears have been reported. Furthermore, Kobayashi et al. 41 have tested CFC of the LRCs discovered in mice and stated that confinement of BrdU could be found in a portion of cells within the colonies derived from mouse perichondrocytes. The authors suggested this was an indication of asymmetric cell division, characteristic of stem cells. Retention of template DNA strands during asymmetric cell division (“immortal strand hypothesis”) is indeed one of the proposed mechanisms of label retention in some stem cell types. However, as with the dormancy, this phenomenon also cannot be generalized to define stem cells, because not all stem cells use this mechanism during self-renewal. 39
Screening for MSC markers led to the identification of two key candidates, CD44 and CD90, which were found to be expressed to a higher degree in perichondrocytes than in chondrocytes.41,45 In particular, in a study on human perichondrium derived from microtia patients, Kobayashi et al. found that perichondrocytes could be efficiently differentiated into chondrocytes in vitro when treated with basic fibroblast growth factor (bFGF) and insulin-like growth factor 1 (IGF1). When these differentiated cells were then expanded and transplanted into nonobese diabetic/severe combined immunodeficiency mice, a neocartilage with neoperichondrium layer expressing Col1 was formed (Fig. 5). In contrast to perichondrocytes, similarly cultured and expanded chondrocytes would only generate Col2-positive cartilage without the perichondrium layer. This signified the presence of bipotent cells in perichondrium but not in the cartilage. Furthermore, the expression of CD90 and CD44 was predominantly found in the perichondrium layer in both native tissues and in the reconstructed cartilage.

Chondrogenic potential of perichondrocytes.
Further experiments by Kobayashi et al. showed that cells coexpressing both CD44 and CD90 markers possessed a higher clonogenic capacity compared with those expressing only one marker. 45 More importantly, when plated at a density of a single-cell per well in a 96-well plate, double-positive cells were reported to form large colonies consisting of several hundred cells, which possessed chondrogenic, adipogenic, and osteogenic potential. Although other studies have also shown multipotency of perichondrocytes in vitro,40,46 these reports were mostly based on the differentiation of a pool of cultured cells, rather than a single cell-derived colony, and they were thus insufficient proof of the existence of multipotent progenitors. 47 The use of single cell-based assays by Kobayashi et al. has therefore by far provided the strongest evidence of the existence of multipotent progenitors in the perichondrium.
To assess their capacity to produce heterogeneous tissues of auricular cartilage in vivo, Kobayashi et al. further transplanted clonally propagated CD44+/CD90+ cells into mice and found that a tissue containing Col2-positive cartilage covered by a Col1-positive layer was produced 3 months after transplantation, indicating that multipotency of the clonally propagated cells was retained. Of note, this newly produced tissue also contained CD44+/CD90+ cells, which authors interpreted as an indication of self-renewal—the hallmark feature of stem cells next to the multipotency. Together, these results bring the study by Kobayashi et al. closest to proving that there is a population of stem and progenitor cells capable of multipotent differentiation and self-renewal within perichondrium. However, definitive proof of self-renewal would only be possible through further probing of the multipotency of secondary CD44+/CD90+ cells found in the neocartilage derived from an expanded single-cell colony.47,48 Such an experiment, to the best of our knowledge, is yet to be performed.
Overall, several studies have demonstrated that there are slow-cycling LRCs in perichondrium that express MSC markers and have a superior proliferation potential and CFC. Furthermore, a study by Kobayashi et al. provided strong evidence for the existence of multipotent progenitors in the perichondrium. However, the self-renewal potential of these progenitor cells remains to be proved.
Perichondrium in Cartilage Tissue Engineering
Considering the presumed functions of perichondrium, such as appositional growth, role in wound healing, regeneration, and metabolic homeostasis of the cartilage tissue, it is clear that the types of cartilage that are normally covered with a perichondrium layer (e.g., auricular and costal) would not be fully functional without it. Furthermore, in such cases, the avascular cartilage tissue relies on perichondrium for its supply of oxygen and nutrients. Preservation of the vascularized perichondrium in native cartilage grafts in reconstructive surgery of the head and neck is therefore highly desirable for graft survival and growth. 49 Tissue-engineered cartilage usually lacks the perichondrium layer. Maintenance of large engineered cartilage constructs (e.g., human auricle) can be associated with such limitations as poor cell viability and proliferation, which result in inhomogeneous tissue formation and/or partial necrosis of the transplants. 50 To promote the integration and growth of the cartilage transplants, placing vascularized flaps between the skin and the transplanted cartilage tissue is a common surgical approach.51–53 The use of flaps, however, comes at a cost to the esthetic outside look of the graft owing to the draping and thickness of the flap. A functional perichondrium would raise the possibility of using a full-thickness skin graft to cover the cartilage–perichondrium construct, thus saving the laborious surgical step of flap dissection and creation of a better esthetic result 54 (Fig. 6).

The relevance of perichondrium in cartilage tissue engineering for head and neck reconstructive surgery. A vascularized perichondrium layer could simplify transplantation of the engineered cartilage constructs and facilitate the use of a full-thickness skin graft without the need for vascularized flaps. Direct draping of the skin graft onto the construct, without an intermediate flap between the graft and the engineered cartilage tissue, could lead to a better esthetic outcome. Furthermore, perichondrium can enhance the flexibility of the cartilage tissue constructs, which might lack the elasticity comparable with the native elastic cartilage. The perichondrium is important for the functioning of the cartilage tissue because of its role in wound healing and regeneration, and the hypothesized function of metabolic regulation, which prevents cartilage calcification. Finally, the perichondrium might support the maintenance of stem and progenitor cells (arrows), which are valuable for tissue engineering and regenerative medicine.
Yanaga et al. have designed a two-step surgical method of engineering an auricular construct with a perichondrium that can be directly transplanted subcutaneously.55,56 The authors harvested cells from remnants of auricular cartilage from microtia patients and cultured them in vitro for 1 month in layered culture in the presence of fibroblast growth factor-2 (FGF-2). The expanded autologous cells were then injected into a subcutaneous pocket in the fascia of the lower abdomen and allowed to form a neocartilage for 6 months. Of interest, upon explanation, it was found that the neocartilage also possessed a layer of perichondrium-like tissue. This explanted block of cartilage tissue was then sculpted to obtain a framework of the auricle and implanted at the defect site into the temporal region, followed by skin grafting after the ear elevation. The authors reported no resorption of the transplants after 2- and 5-year follow-up inspection. Furthermore, authors hypothesized that the successful inosculation of the skin graft after ear elevation indicated the formation of functional neoperichondrium with the capacity to generate vascular supply around the neocartilage. 55 The formation of the perichondrium-like layer has also been observed in vitro when cells obtained from rabbit ears were cultured under continuous flow in a bioreactor.57,58 In these experiments, perichondrocytes and chondrocytes were purposefully cocultured to induce the formation of the perichondrium layer. However, the functionality of these constructs in vivo has not been assessed.
Besides the vascular supply, perichondrium can play an important role in determining the mechanical characteristics of the specific cartilage type. In the comparative study discussed in the previous sections, Bairati et al. observed structural differences in perichondria derived from different tissues (Fig. 3A) and speculated that this variation might be important to achieving the demands of the particular anatomical site. 21 The authors proposed that cartilage and perichondrium should be regarded as a single anatomical unit in the skeletal system. An illustration of this concept was also given in a histological study on the perichondrium of nasal septum and nasal lobular cartilage. 22 In this example, the perichondrium of lobular cartilage was found to have a higher content of elastic fibers and a significantly thicker outside portion compared with that of the septal cartilage. These differences were suggestively attributed to the function of the cartilage; namely, lobular cartilage is located in the nasal tip and requires mobility, whereas the nasal septum has a mere supportive function.
Viewed from the perspective of tissue engineering, this structural role of perichondrium is relevant. The presence of the perichondrium layer is crucial to the flexibility of human auricular cartilage, which is essential to proper anatomical functioning and maintenance of the integrity of an engineered auricular graft. 59 This was illustrated in a study where the addition of layers of lyophilized swine perichondrium was found to increase the flexibility of in vitro engineered cartilage. 60 This layer of pseudoperichondrium, as the authors call it, efficiently protected the cartilage constructs from fracture when subjected to gross mechanical testing by bending and torsion. Furthermore, the constructs with a perichondrium layer were able to return to their initial shape after the deformation.
Overall, a functional layer of perichondrium can simplify transplantation, survival, and maintenance of a cartilage graft by providing vascular supply and nutrients. Furthermore, mechanical characteristics of perichondrium should correspond to the specific anatomical function of the cartilage type, and thus should not be overlooked in the development of tissue-engineered cartilage constructs.
Concluding Remarks
Perichondrium is indispensable for the nutritional and vascular supply of the underlying cartilage tissue and can become invaluable to the maintenance of transplanted auricular tissue-engineered constructs (Fig. 6). Perichondrium might also be implicated in metabolic regulation of cartilage, determining the proper functioning of chondrocytes, and inhibiting calcification. For cartilage tissue engineering, the structural role of perichondrium is relevant and should be taken into account and should correspond to the anatomical function of the specific cartilage type.
Particularly noteworthy is the strong evidence that perichondrium might house multipotent progenitors and therefore have regenerative potential that can be harnessed for tissue engineering. However, definitive proof of the role of perichondrium as a stem cell microenvironment still needs to be achieved. Furthermore, translation of the existing findings into tissue engineering applications is still hampered by the insufficient phenotypic characterization of the putative progenitors, which is required to reproducibly isolate these cells based on a defined set of markers while excluding any ambiguity and contamination with fibroblasts or chondrocytes. The studies on the characterization of cellular components of human perichondrium are in turn limited by the anatomical scarcity of the tissue and the low harvesting capacity caused by the need for preservation of perichondrium during surgical interventions. Even so, with the advancement of tissue engineering, the ambitious goal of fully reproducing functional anatomical structures with complex architecture such as the human auricle is within reach, making the efforts of overcoming these limitations indubitably worthwhile.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Swiss National Science Foundation (CRSII5_173868 to N.R.).