
Abstract
Stem cells (SCs) hold great potential for regenerative medicine, tissue engineering, and cell therapy. The clinical applications of SCs require both high quality and quantity of transplantable cells. However, during conventional in vitro expansion, SCs tend to lose properties that make them amenable for cell therapies. Extracellular matrix (ECM) serves an essential regulatory part in the growth, differentiation, and homeostasis of all cells in vivo, and when signals are transmitted to cells, they do not respond passively. Many cell types can remodel pericellular matrix to meet their specific needs. This reciprocal cell–ECM interaction is crucial for the conservation of cell and tissue functions and homeostasis. In vitro ECM remodeling also plays a key role in regulating the lineage fate of SCs. A deeper understanding of in vitro ECM remodeling may provide new perspectives for the maintenance of SC function. In this review, we critically examined three ways that cells can be used to influence the pericellular matrix: (1) exerting tensile force on the ECM, (2) secreting a variety of ECM proteins, and (3) degrading the surrounding matrix, and its impact on SC lineage fate. Finally, we describe the deficiencies of current studies and what needs to be done next to further understand the role of ECM remodeling in ex vivo SC cultures.
Impact statement
The effect of extracellular matrix (ECM) remodeling on physiological activities and disease progression has been extensively studied, but its effect on in vitro stem cell (SC) culture has received insufficient attention. More and more research has shown that in vitro ECM remodeling is critical for maintaining SCs function. This review will highlight how cells remodel ECM and the significance of ECM remodeling in ex vivo SC culture, as well as summarize the shortcomings of current research and what needs to be done next to further our understanding of the role of ECM remodeling in ex vivo SC culture.
Introduction
Stem cells (SCs) have the potential for differentiation into various specialized cell types, as well as capacity for self-renewal, therefore holding great potential for cell-based therapy, such as regenerative medicine and tissue engineering.1–3 However, the clinical application of SCs requires both large numbers and high quality of transplantable cells. 4 This requires mass expansion of SCs, while at the same time, maintaining its stemness. Conventional methods for maintenance and expansion of SCs depend upon two-dimensional (2D) culture techniques utilizing xenogeneic media and plastic culture plates. 5 The use of xenogeneic media may potentially lead to transmission of pathogens and may influence reproducibility of experiments due to batch variation in the materials used. 6 Moreover, cells appear to lose their stemness upon prolonged ex vivo culture, and these approaches only provide limited expansion. 5
Extracellular matrix (ECM) is a three-dimensional (3D) noncellular macromolecular network, and plays important roles in regulating growth, differentiation, and homeostasis of all cells in vivo. ECM is mostly composed of proteoglycans, fibrous proteins, and water. The main fibrous proteins constituting the ECM are collagens, laminins, and fibronectin. 7 Due to differences in physical properties (stiffness, micropattern, viscoelasticity) and composition of ECM in different tissues, the effects of ECM on different cell types are different. For example, ECM mechanics have been shown to regulate cell metabolism, with recent studies showing that transferring human bronchial epithelial cells from stiff to soft substrates caused downregulation of glycolysis, and that the phenotype and function of human antigen-presenting dendritic cells are affected by substrate stiffness.8,9 On the other hand, modifying cell metabolism can also modulate ECM mechanics. For example, exogenous pyruvate is needed by breast cancer cells to support crosslinking of collagen at the metastatic niche, thus stiffening ECM. 10 Additionally, high glucose levels can induce increased secretion of ECM components by mesangial cells in the kidney, which has been linked to the uncontrolled build-up of ECM that occludes capillaries and affects renal function in diabetic nephropathy. 11
However, in both static and dynamic environments, when signals are transmitted to cells, they do not respond passively. Instead, cells in vivo can sense and remodel their local environment to meet their needs.12,13 This reciprocal cell–ECM interaction is crucial for maintaining functions and homeostasis of cells and tissues. Disrupting this balance may lead to pathological conditions and accelerate progression of disease.14,15 For instance, abnormal accumulation of ECM proteins can result in fibrosis, leading to distortion of normal layout and stiffness of tissues. 16 Myofibroblasts are main producers of ECM in many tissues and express high levels of tissue inhibitors of metalloproteinases (TIMPs) and fibrillar collagens. Excessive fibrosis in the ECM can activate fibroblasts to further increase the production of fibrotic ECM, thus generating a positive feedback loop that aggravates fibrosis. 17 Clearance of myofibroblasts or overexpression of fibrillar collagen degradation enzymes can significantly ameliorate established fibrosis in animal models,18,19 which means ECM remodeling can be a potential therapeutic target.
Since ECM remodeling is so essential in maintaining cellular and tissue homeostasis in vivo, some studies began to investigate the involvement of ECM remodeling in maintaining the function of SCs cultured in vitro. They found that regulating ECM remodeling exerted a significant impact on the fate of SCs.20,21 The purpose of this review is to provide a general overview of how cells remodel ECM, the effect of ECM remodeling on SCs within different culture systems (3D culture system or 2D culture system) and how to achieve a comprehensive understanding of ECM remodeling in vitro.
How Cells Remodel ECM
ECM, cell adhesion, and traction
Some studies have shown that cellular contractions and ECM–integrin interactions have a significant impact on SCs function. So, in this section, we will introduce how ECM and integrins interact with one another, as well as how cellular contractions remodel ECM.
To exert force on the ECM, the cells first need to form a tight junction with the ECM through receptors on its surface (① in Fig. 1). Integrins have long been known as the primary molecular connection linking cells to the ECM, and as bidirectional hubs for signal transmission between cells and their surroundings.

How cell remodel ECM. ① Adhesion to the ECM through focal adhesion proteins and integrins, followed by actomyosin contractility to arrange and stretch the ECM; breakdown of ECM proteins through ② the secretion of ECM degradation enzyme; or ③ membrane-binding ECM degradation enzyme; ④ secretion of new ECM proteins. ECM, extracellular matrix. Color images are available online.
Integrins are attached to actin cytoskeleton by means of binding proteins (Fig. 2), such as talin, vinculin, filamin A, and α-actin. 22 Talin is associated with force-mediated adhesion reinforcement, 23 and there is a competitive binding of integrins with α-actin and filamin with talin.24–27 α-actin is related to nascent focal adhesions (FAs) formation, which enable the transmission of high forces during adhesion maturation. 28 Filamin A is very similar to talin and appears to be a true mechanoreceptor, as stretching can adjust its connection with binding partners.29,30 A recent study showed that Filamin A regulates remodeling of the ECM through synthesis and degradation of collagens. 31
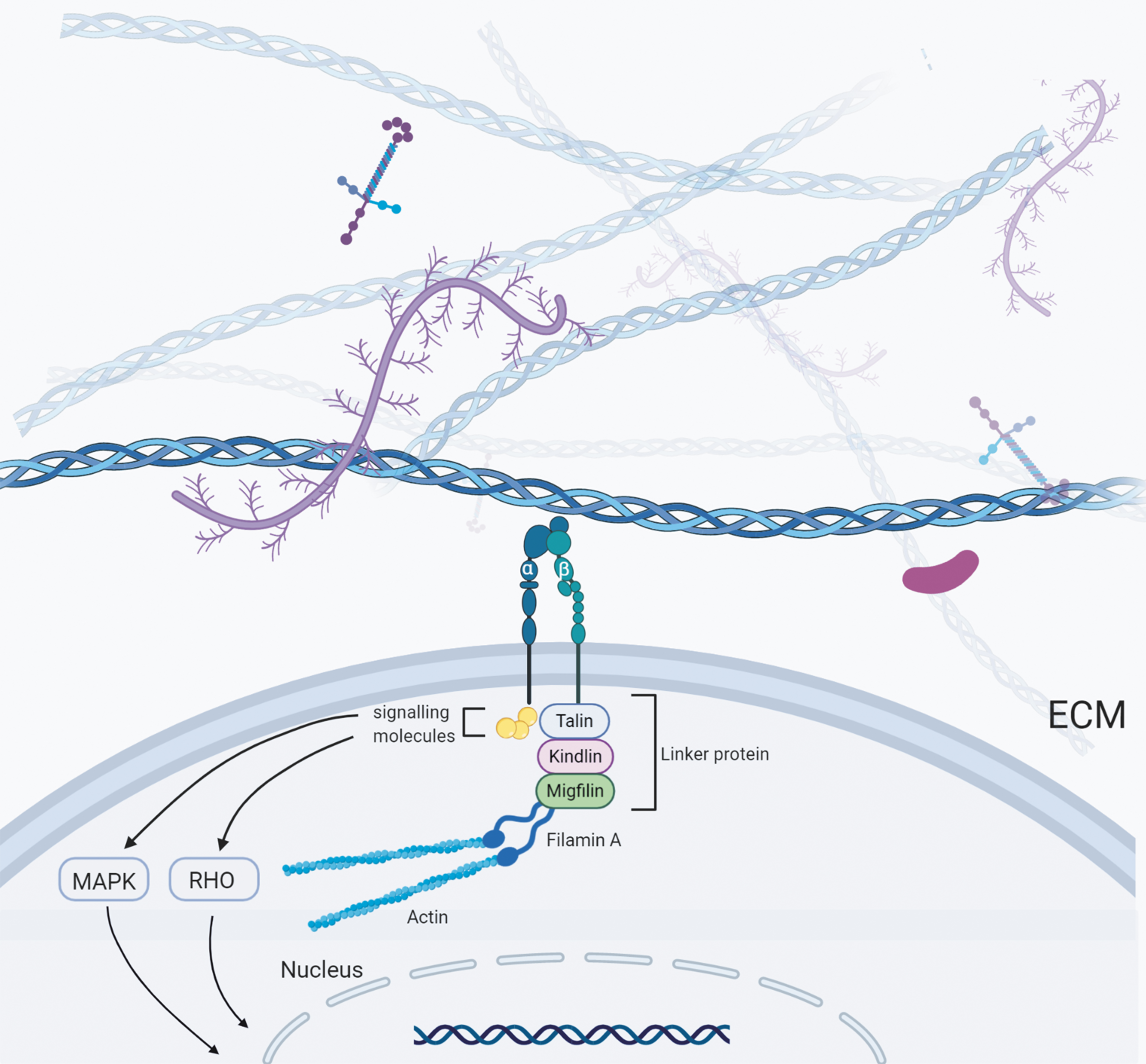
Mechanical interaction between cell and ECM. Cells interact mechanically with ECM through transmembrane receptors called integrins, which interact with intracellular signaling molecules (including FAK and SRC) through a series of association proteins (including α-actin, filamin, migfilin, kidnlin, vinculin, talin). ROCK and MAPK signaling pathways are essential in the activation of integrins. FAK, focal adhesion kinase; MAPK, mitogen-activated protein kinase; ROCK, RHO-associated protein kinase. Color images are available online.
Under pulling force from myosin II activity or pushing force from actin polymerization, F-actin moves backward and drives the linker proteins to move backward. 22 This is the mechanism by which forces are transferred to integrins that drag the ECM. Rigid matrix can resist this force, and the binding integrin remains immobile. Compliant ECM deforms under this force and the bound integrin molecules also move. The movement of integrin reduces the net loading rate on all force-bearing components, which leads to changes in cell response. 32 The RHO–ROCK (RHO-associated protein kinase, ROCK) signaling pathway is crucial in this process, as the RHO effector ROCK is crucial for the maintenance of myosin II activity, which, together with complementary actin polymerization, enables cells to forcefully act on the ECM. 22
Cells use their own contractile force to regulate various physiological processes, including intracellular transport, cell division, and migration, as well as ECM organization and homeostasis.22,33–36 There are various different modes of cell contraction, including isotropic contraction and anisotropic contraction. Such anisotropic contraction which fibroblasts or endothelial cells generate on the surrounding ECM or adjacent cells in vivo is used in matrix remodeling and tissue morphogenesis. 32
Degradation of original ECM
The degradation of ECM proteins by cells is a significant component of ECM remodeling. 14 It is important for repairing damaged tissues and release of biologically active molecules. 37 Abnormal ECM degradation plays an important role in cancer metastasis. For example, high MMP (matrix metalloproteinase) activity at the primary tumor site leads to a disrupted vasculature that allows tumor cells to intravasate and enter the circulation. 38 Additionally, neutrophils can induce laminin degradation in an inflammatory environment by elastase and MMP-9. A specific laminin epitope is exposed as a result of this proteolytic action, which activates an integrin-mediated signaling cascade, resulting in the reawakening of dormant cancer cells at metastasis sites and their subsequent proliferation. 39 ECM degradation is also significant in ex vivo SC culture. In 3D culture systems, substrate degradability is an important target for regulating SC fate.21,40 In this section, we will discuss the degradation of ECM. Extracellular degradation and lysosomal degradation are two main ECM degradation pathways. 41
Extracellular or pericellular degradation (② and ③ in Fig. 1) is regulated by different families of proteases, like ADAMs (a disintegrin and metalloproteinases), MMPs, ADAMTS (ADAMs with a thrombospondin motif). 14 The most commonly involved enzymes in ECM degradation are MMPs. In general, their activity is low but increases under some specific conditions, including wound healing, embryonic development, or some disease condition.37,42 Cancer cells, for example, induce surrounding host cells to generate MMPs in a paracrine manner by secreting extracellular MMP inducers, growth factors, interferon, and interleukin. 43
For now, researchers have found 23 human MMPs. They are produced in two ways, secretion into extracellular space (② in Fig. 1) or anchored on cell membranes (③ in Fig. 1). Most MMPs are secreted as zymogens and then activated through proteolytic cleavage or modified by oxidation in the extracellular space. 44
The key function of ADAMs is to cut the ectodomains of transmembrane proteins close to the cell surface, thereby freeing the full ectodomain of adhesion molecules, receptors, growth factors, and cytokines. 45 At the same time, some ADAMs can also degrade ECM proteins such as collagens. The ADAMTS protein family are secreted proteinases with different functions, including aggrecanases and procollagen N-propeptidases. The latter is important for tissue-specific deposition of collagen fibrils onto ECM. 46 So far, the roles of many ADAMTS remain uncertain.
Lysosomal degradation after receptor-mediated internalization is another ECM degradation pathway. Internalization of different ECM proteins is controlled by different mechanisms. For example, α2β1 integrin and Endo180 (also known as uPARAP) are two receptors that control collagen internalization. 41 The nonmuscular myosin II A and actin-binding protein gelsolin aggregate at the adhesion sites after collagen I binds to α2β1 integrin in fibroblasts, which is necessary for phagocytosis and collagen internalization into early endosomes. 47 Then collagen is delivered into lysosomes for degradation. 48 Endo180 has been shown to be related to the internalization of several collagens, including collagen I, IV, and V, through clathrin-dependent endocytosis.49,50 Moreover, Endo180, not β1 integrin, is required for the internalization of soluble collagen, which indicates that Endo180 mediates a different mechanism from fibrillar collagen internalization. 51 It is still unclear what determines the internalization of collagen, α2β1 integrin or Endo180, and further studies are required to explain this point. The α5β1 integrin can regulate the absorption of fibronectin through a caveolin-1-based pathway.52,53 This integrin is necessary for myofibroblast internalization of both soluble and matrix fibronectin. 53 Furthermore, a recent study showed that internalized ECM can be resecreted. 54 However, it is still unclear which mechanism governs ECM protein distribution to various endosomal compartments, leading to degradation or secretion. 41
Extracellular proteolysis and matrix internalization are not independent processes. For example, a precondition for receptor-dependent ECM endocytosis has been considered to be the proteolytic cleavage of ECM fibers. 55 While both intact and cleaved collagens can be internalized, the binding of the cleaved collagen to Endo180 is increased and endocytosis can progress more efficiently. 56
The degradation of ECM can release a variety of bioactive fragments that regulate various biological processes, such as angiogenesis, adipogenesis, and fibrosis. These fragments are known as matricryptins or matrikines. 57 For instance, MMP-2 and MMP-9 release the C-1158/59 peptide (14 residues) from collagen I, which increases angiogenesis and promotes wound healing after myocardial infarction. 58 This means that ECM fragments produced by ECM remodeling may also be essential for ex vivo SC culture.
Nascent ECM protein deposition
Some studies have attempted coating ECM proteins before seeding SCs on the dish, which has a major effect on the lineage fate of SCs.59,60 Additionally, SCs cultured in vitro can also deposit ECM. In this section, we will introduce and describe the secretion of ECM proteins.
In healthy tissues, the degradation of ECM is balanced by the deposition of ECM proteins (④ in Fig. 1).7,14 Disruption of this delicate balance can lead to pathological conditions. 61 For example, abnormal osteopontin accumulation causes adipose tissue inflammation, which leads to fibrosis and insulin resistance (a hallmark of type 2 diabetes). 62 In vivo, the deposition of ECM proteins is performed by a number of cell types.63,64 One of the main groups of cells that contribute to ECM protein deposition is considered to be fibroblasts.65,66 Fibroblasts produce ECM molecules that regulate tissue strength and elasticity. Recently, Shook et al. found that there are multiple subsets of ECM-producing myofibroblasts at the wound bed. They observed that macrophages and fibrotic environments can affect the composition of myofibroblasts, with different myofibroblast subpopulations having different abilities to secrete ECM proteins. 67 ECM production of fibroblasts can also be regulated by myogenic progenitor cells (MPCs). Fry et al. found that the exosomes secreted by MPCs contain miR-206, which can inhibit Rrbp1, the major regulator of collagen biosynthesis in fibroblasts, which avoids excessive deposition of ECM. 68
Effects of ECM Remodeling on In Vitro SC Culture
ECM remodeling in 2D cell culture systems
Due to recognition of the key roles of ECM in regulating the fate of SCs, current 2D cultures began to use different methodologies, including sandwich culture (placing cells between two layers comprising of ECM), varying micropatterning (engineered 2D surface patterning), and altering substrate stiffness. 69 Moreover, SCs in 2D culture can remodel their pericellular ECM (Fig. 3A). The effects of ECM remodeling should draw more attention. However, evaluating the effects of ECM remodeling on SCs directly in 2D cell culture systems is challenging. The presence of decellularized ECM (dECM) makes it much easier to evaluate the role of ECM remodeling in ex vivo SC culture. For the processing of dECM from cultured SCs, two main methods are currently used: exposure to chelating agents or nonionic detergents. 70 Before SCs are seeded, dishes are coated with dECM. In the subsequent subsections, the influences of these dECM on SC fate will be discussed (Table 1).
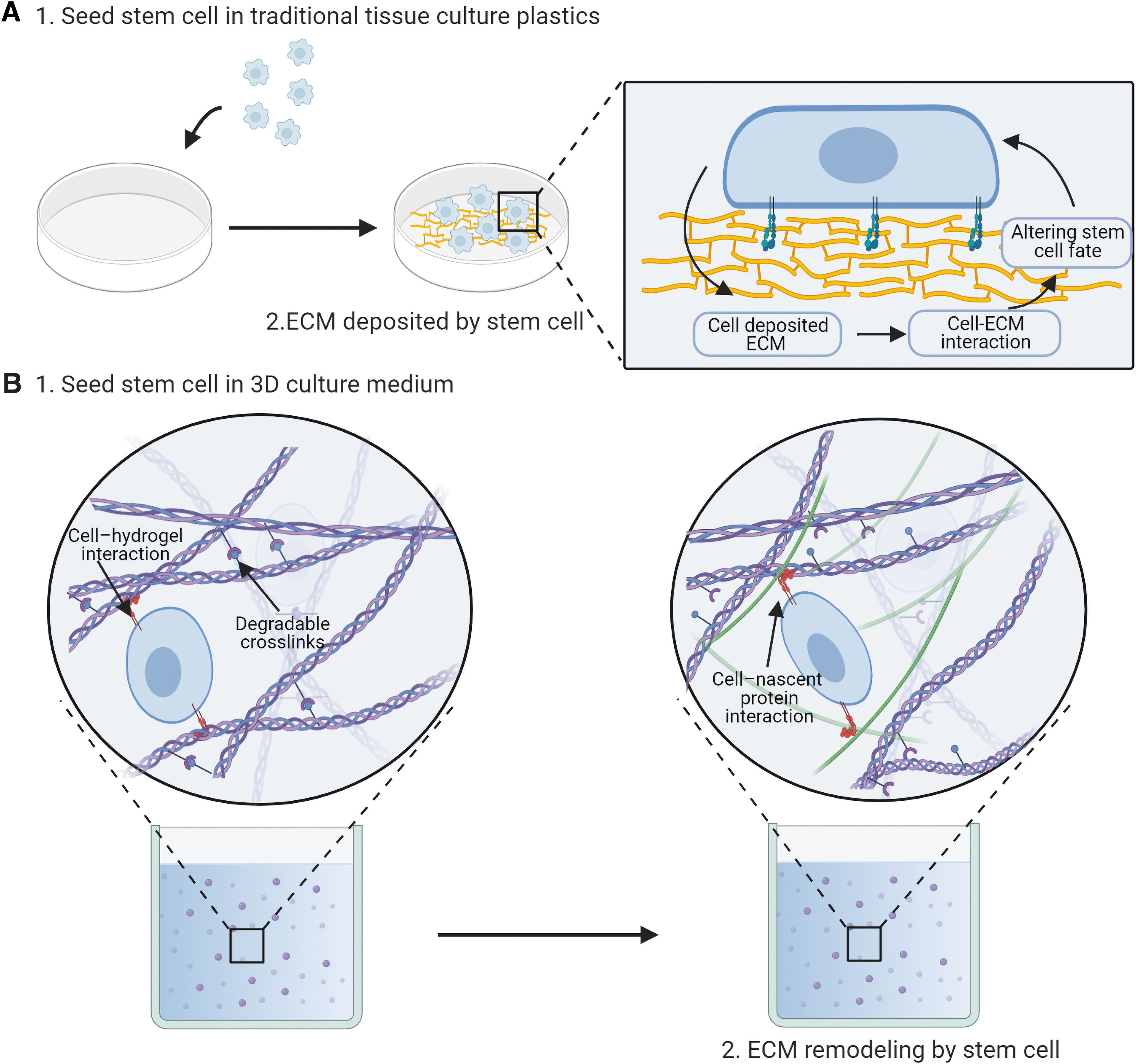
Schematic diagram of ECM remodeling in 2D/3D culture system.
The Effect of Extracellular Matrix Remodeling in Two-Dimensional Culture
ASCs, adipose stem cells; dECM, decellularized extracellular matrix; DPSCs, dental pulp stem cells; ESCs, embryonic stem cells; LIF, leukemia inhibitory factor; MSCs, mesenchymal stem cells; NR, not reported; SDSCs, synovium-derived stem cells.
Stemness can be maintained by ECM remodeling in growth media
In growth media, stemness can be maintained by ECM remodeling. The maintenance of stemness is manifested by the enhancement of self-renewal capacity and ability to stay undifferentiated in growth medium, followed by enhancement of differentiation potential in differentiation media.
dECM, derived from adipose SCs (ASCs), can enhance the proliferative and multidifferentiation (adipogenic and chondrogenic capacity) potential of ASCs. He and Pei observed that cell proliferation on dECM-expanded ASCs was 6–10 times that of noncoated flasks. When cultured in serum-free chondrogenic/adipogenic medium, dECM-expanded ASCs expressed more typical chondrogenic/adipogenic markers. 71
Dental pulp SCs (DPSCs)-derived ECM can balance replication and mineralization of DPSCs. Zhang et al. found that compared with DPSCs grown on conventional plastic dishes, ECM-expanded DPSCs showed enhanced proliferative capacity and fewer cells involved in the process of occasional mineralization. 72
SCs culture on plastic also generated ECM, however, the stemness of ECM-expanded SCs was better maintained. This means that SCs may lose some key properties when in direct contact with conventional tissue culture wares over prolonged periods of culture.
The stemness of mesenchymal SCs (MSCs) have been shown to decline with donor age. Ng et al. found that the key characteristics of human adult MSCs (haMSCs) can be restored by expanding cells on the dECM generated by human fetal MSCs (hfMSCs). Compared with haMSC-derived ECM or traditional tissue culture wares (TCPS), haMSCs cultured on hfMSC-derived ECM were more proliferative. These haMSCs were smaller, more similar in shape, and maintained multipotency during ex vivo culture. 73
Fetal synovium-derived SC (SDSC)-deposited ECM can also revitalize the stemness of adult SDSCs. Li et al. observed that fetal SDSC-derived dECM (FE) was better at encouraging cell growth and chondrogenic potential of adult SDSCs relative to adult SDSC-derived dECM (AE). From proteomics analyses, they found that both ECM have some unique proteins. They discovered that FE had several distinct matrix proteins, such as tenascin, fibrillin-2, and versican core protein, while AE had some unique matrix proteins, such as thrombospondin-1, periostin, fibulin-6, and elastin. They also analyzed the mechanical properties of these two ECM, with the results showing that adult ECM exhibited higher elasticity than fetal ECM, which might be responsible for the rejuvenative effects of fetal ECM. 74
The SC characteristics of MSCs often decrease on normal culture surfaces during extended ex vivo expansion. Pei et al. discovered that dECM created by early-passage MSCs can rejuvenate properties of late-passage MSCs. Compared with those culture on TCPS, the late-passage MSCs (passage 5) cultured on the dECM derived from early-passage MSCs (passage 1 or 2) were smaller in size, had stronger chondrogenic and osteogenic differentiation potential, and showed a threefold rise in proliferation over a 7-day culture duration. 75 These results thus showed that the loss of stemness of adult or late-passage SCs may be due to changes in matrix remodeling ability.
Since ECM deposited by SCs in 2D culture system plays such an important role in regulating SC fate, there are some studies attempting to coat specific ECM proteins (artificially regulating ECM remodeling) before SCs are seeded, so as to maintain the stemness of SCs. For example, Wilkinson et al. found that type I collagen, fibronectin, and laminin can affect early cell lineage fate decisions of hematopoietic stem and progenitor cells (HSPCs). They discovered that fibronectin-coated substrates can maintain HSC populations, whereas collagen-coated substrates enhanced proliferation, and that laminin-coated substrates improved myeloid differentiation, after 24 h culture in serum-free medium. They also found that suppressing myosin II-mediated contraction abolished the effects of fibronectin or collagen coating on the maintenance of primitive HSCs. In addition, when primary HSCs cultured on fibronectin-coated substrates were treated with either integrins α5β1 or ανβ3 antibody, HSCs differentiation toward multiple myeloid lineages was observed. These findings thus suggest that intracellular tension and integrin activation play a key role in regulating the lineage fate of HSCs. 60 Furthermore, Rodin et al. revealed that laminins 511 and 521 can promote the proliferation of human induced pluripotent SCs through the activation of integrin a6b1 and the PI3K/Akt pathway. 76 These results thus suggest that ECM remodeling may regulate the activation of integrin and cell contraction to maintain stemness.
ECM remodeling in differentiation media enable SC differentiation
In differentiation media, ECM remodeling plays a different role. Zhang et al. assessed the effect of dECM derived from stepwise adipogenic induction of ASCs on ASCs themselves. ASCs were cultured in proliferation and adipogenic media, and dECM deposited by ASCs was characterized and decellularized. They found that during adipogenesis, ECM secreted by cells was remodeled from fibronectin dominant to laminin dominant. ASCs cultured on growth dECM showed higher migration potential, whereas ASCs cultured on adipogenic dECM were more likely to undergo adipogenic differentiation. They also found that integrin subunit α7, α6, and αv expression were substantially higher in both growth and adipogenic dECM-cultivated ASCs, indicating that integrins are essential for ASC migration and adipogenesis. 77
Guneta et al. cultivated ASCs in proliferation, osteogenic, and adipogenic media, and characterized and decellularized the secreted ECM. They observed that more collagen and glycosaminoglycans (GAG) were produced by ASCs in osteogenic media relative to ASCs in growth and adipogenic media. These two differentiated dECM have subsequently been shown to trigger the initial differentiation of ASCs into the osteogenic or adipogenic lineages, respectively, as shown by the increased expression of lineage-specific markers. Additionally, the expression of some integrin subunits (α6 and β1) was observed to be upregulated in ECM-expanded ASCs. Once again, this shows that integrins may be essential for cell–ECM interactions. 78
Yan et al. evaluated the effects of three separate ECMs derived from embryonic SCs (ESCs). These ECMs are derived from decellularized ESC-derived neural progenitor cell (NPC) aggregates spontaneously differentiated embryoid bodies (EB), and undifferentiated ESC aggregates (AGG). They observed that the stemness of ESCs could be best sustained by the AGG and EB ECM, whereas NPC-derived ECM downregulated their stemness. They also tested the neural induction ability of these ECMs and observed that cells in the NPC group showed higher expression level of neural marker. Proteomic analysis of ECMs showed significant differences in ECM composition and neurotrophins in the NPC group compared with the AGG and EB groups. There are significantly different β-catenin expression levels in cells cultivated on different ECM, indicating that the Wnt/β-catenin signaling pathway is required for cell–matrix interactions, which is consistent with previous research.79,80
The remodeled ECM will facilitate the differentiation of SCs into specific lineages when SCs were cultivated in differentiation media. This means that different culture media may play a role in part by influencing how cells remodel ECM, and the underlying mechanisms still need further investigation.
ECM remodeling in 3D cell culture systems
Due to its capacity to mimic tissue-like structures, 3D culture systems are becoming more and more common compared with 2D culture. At present, there are several distinct 3D methods of cell culture, including: liquid overlay cultures, hanging drops, hydrogels, bioreactors, scaffolds, nichoid and 3D bioprinting. Those 3D culture systems that replicate the in vivo SC niche have been shown to be capable of preserving SC stemness.81,82 Hydrogels are widely used as synthetic ECM for 3D cell culture and have been frequently used to develop tissue engineering. In the following subsection, we will discuss the effect of ECM remodeling on the fate of SCs in hydrogels (Fig. 3B).
ECM degradation has a major impact on SC fate in 3D hydrogels
The impact of degradability and matrix stiffness on the preservation of NPC stemness in 3D hydrogels were evaluated by Madl et al. They encapsulated NPC in proteolytically degradable polyethylene glycol (PEG) hydrogels or physically remodelable alginate hydrogels, and found that with increasing hydrogel degradability, the expression of Nestin and Sox2 (NPC stemness markers) increased dramatically, but not with increasing stiffness (elastic modulus ranging from ∼0.5 to 50 kPa). Moreover, NPC proliferation and differentiation capacity also increased with increasing hydrogel degradability.
To explore whether cellular traction is involved in the maintenance of NPCs stemness, cytoskeletal tension inhibitors against myosin II, Cdc42, RhoA/B/C, PAK1, and ROCK were used. The results showed that the inhibition of cellular traction had no effect on NPCs stemness, indicating that the preservation of stemness in high-degradability gels was not due to the production of tensile force. 20
In a later study, it was found that upon suppressing the YAP/TAZ signaling pathway, NPCs in degradable hydrogels showed a significant downregulation of stemness marker Nestin and Sox2. Inhibiting NPC-mediated degradation of hydrogels by ADAM9 knockdown led to a major reduction in YAP expression, which means that matrix degradation in 3D hydrogels maintained the stemness of NPCs by promoting the expression of YAP/TAZ. 83
Ferreira et al. found that MSC-mediated degradation in 3D hydrogels is related to osteogenesis. The hydrogel they used to culture MSCs were covalently crosslinked to poly ethylene glycol diacrylate (PEGDA) and thiol-modified hyaluronic acid (S-HA). They fabricated a range of hydrogels with varying compositions of S-HA and PEGDA, by changing the concentrations of PEGDA, but retaining the same concentration of S-HA. They found that MSCs encapsulated in efficiently crosslinked hydrogels (1:0.75, with a ∼10 kPa initial elastic modulus) favored osteogenesis, which in turn mediated a significant decrease in elastic modulus, in the absence of chemical stimulation. Inhibition of hyaluronidase activity by Vcpal can inhibit osteogenesis of MSCs in 1:0.75 hydrogels, which means that MSC-mediated degradation in S-HA-PEGDA hydrogels is related to osteogenesis. 21 However, they did not investigate the underlying mechanisms, which was revealed by subsequent later studies.
Lee et al. observed that enhanced osteogenesis of MSCs is associated with matrix remodeling-mediated cell volume expansion. Physically remodelable alginate hydrogels (with a ∼20 kPa initial elastic modulus, as this modulus has been shown to facilitate osteogenesis) were used to cultivate MSCs. They found that inhibiting volume expansion by slow stress relief hydrogels or increasing osmotic pressure suppressed MSC osteogenesis. On the other hand, increasing cell volume expansion by hypoosmotic pressure promoted osteogenesis. Subsequently, they found that volume expansion of MSCs is related to the triggering of TRPV4 ion channels, and these two factors together regulate nuclear localization of RUNX2 to facilitate osteogenesis. 84
Moreover, the differentiation of MSCs in 3D hydrogels might be related to degradation-mediated cellular traction. Khetan et al. found that compared with human MSCs (hMSCs) within methacrylated HA (MeHA), hydrogels that restricted cell-mediated degradation, hMSCs within degradable MeHA hydrogels displayed higher tractions and high levels of cell spreading, with a bias toward osteogenesis in adipogenic/osteogenic media. However, when treated with Y-27632 (an inhibitor of ROCK), MSCs cultured in degradable MeHA hydrogel (with a ∼20 kPa initial elastic modulus) showed reduced cellular tractions and decreased osteogenesis, but increased adipogenesis instead. 40
Nascent protein deposition can alter SC fate in 3D hydrogels
Loebel et al. found that local nascent protein deposition and remodeling can alter hMSC's fate in 3D hydrogels. They use a bio-orthogonal labeling technique to visualize nascent protein deposition by MSCs within hydrogels and found nascent protein deposition within a day of culture across a variety of different hydrogels (physically crosslinked hydrogels [agarose and alginate] or covalently crosslinked hydrogels [NorHA, MeHA, and PEG-MAL], all with ∼9 kPa elastic modulus). 85
By using Paxillin to visualize FAs, they found there are a direct interaction between MSCs and nascent ECM during adhesion and spreading in degradable NorHA hydrogels, with this binding having a significant influence on cell behavior. Inhibiting this binding by anti-integrin alpha2 or HFN7.1 (function-perturbing monoclonal antibodies to selectively block the interaction of MSCs with secreted collagen or human fibronectin) can reduce cell spreading, nuclear translocation of YAP/TAZ, and resulted in a shift toward adipogenic differentiation in bipotential adipogenic/osteogenic differentiation media. Similar trends in cell behavior were also observed while limiting ECM protein degradation with TIMP-3 or perturbing the transport and secretion of extracellular proteins with EXO-1. 85
Ferreira et al. found that pericellular ECM secreted by MSCs is related to adipogenesis. S-HA-PEGDA, the hydrogels they use, offer insight into the role of cell-secreted ECM in directing fate, because they provide no sites for integrin-mediated interactions, limiting integrin-mediated interactions to those with the cells' own secreted ECM. They found that MSCs encapsulated in inefficiently crosslinked hydrogels (1:0.375/1:3, with a ∼1 kPa initial elastic modulus) favored adipogenesis and can cause a significant increase in elastic modulus, and inhibiting protein secretion with EXO-1 abrogated adipogenesis of MSCs in 1:3 or 1:0.375 hydrogels. 21
The influence of artificial ECM remodeling on SC fate
Since ECM remodeling is so critical in ex vivo SC culture, several studies have attempted to artificially control the physical properties of synthetic ECM, such as hydrogel, to regulate the fate of SC in vitro.86–88 In the following section, we will discuss the effects of physical properties, such as mechanical stiffness, stress relaxation, and topography, of synthetic ECM on SC fate.
Mechanical stiffness
Substrate stiffness has been shown to influence cell lineage commitment, morphology, and proliferation in both 2D and 3D in vitro cell cultures, as well as under in vivo conditions.89,90 This makes substrate stiffness an excellent target for regulating the fate of SCs. Hogrebe et al. cultured hMSC in KFE-RGD (Arg-Gly-Asp) hydrogels and the stiffness of the hydrogels was controlled by altering total peptide concentration. They found that upon culture in adipogenic differentiation medium, all soft gels (with a ∼0.5 kPa initial elastic modulus) induced higher PPARγ-2 (adipogenesis marker) expression than stiffer gels (with a ∼3 kPa initial elastic modulus), which means that soft substrate favored adipogenesis of MSCs. 91
Darnell et al. utilized RNA-seq to investigate the impact of substrate stiffness on mouse MSCs. MSCs were cultured in alginate hydrogels with different substrate stiffness (ranging from 3 to 30 kPa) for 40 h before performing RNA-seq. A rise in the expression of alkaline phosphatase (an early marker of osteogenesis) was observed in rigid gels. Enrichment analysis showed a significant enrichment for processes, such as bone remodeling, cell–matrix adhesion and proteolysis between the 3 and 30 kPa conditions. These findings thus shed light on previously unknown aspects of MSC stiffness response. 92
Stress relaxation
Natural ECMs exhibit stress relaxation and viscoelasticity. Modulating the stress relaxation of synthetic ECM is another effective way to regulate the fate of SCs. Chaudhuri et al. found that modifying stress relaxation of 3D hydrogels had a significant effect on SCs lineage fate, independent of cell-adhesion-ligand density, initial elastic modulus, and degradability of hydrogel. They found that osteogenic differentiation, cell proliferation, and spreading of MSCs are all enhanced in cells cultured in rapidly relaxing hydrogels. Interestingly, MSCs form a mineralized, collagen-1-rich matrix similar to bone in gels with faster relaxation (with a ∼17 kPa initial elastic modulus). Additionally, they also showed that adhesion-ligand binding, actomyosin contractility, and mechanical clustering of adhesion ligands mediate the effects of stress relaxation. 93
Topography
While stress relaxation and substrate stiffness can influence SC fate, substrate topography also plays an important role in regulating SC function. Li et al. found that topography is a key factor in influencing rat MSC morphology, spreading, and differentiation. They discovered that MSCs cultured in pillar polyacrylamide (PA) hydrogel were larger in shape and spread faster than MSCs cultured within groove hydrogel. They also found that MSCs in stiff pillar hydrogel favored osteogenic differentiation, whereas MSCs in soft groove hydrogel favored neuronal differentiation. 94
Ferlin et al. investigated the effects of two 3D-printed pore geometries (cubic or cylindrical pore geometries) of synthetic polymer poly(propylene fumarate) (PPF) on hMSC differentiation. They found that under adipogenic or chondrogenic induction, hMSCs in the cubic hydrogel demonstrated a substantial upregulation of the expression of key cell phenotype markers for each respective lineage, as compared with cylindrical hydrogel. 95
Perspectives on Ex Vivo ECM Remodeling
Current problems
As we previously discussed, ECM remodeling plays a vital part in ex vivo SC's culture, which means it can be a potential regulatory target for the maintenance of stemness. To achieve this, we need to have a comprehensive understanding of in vitro ECM remodeling. However, there were still some problems in previous studies. For instance, the physical characteristics of ECM, like substrate rigidity and topography, also exert a profound influence on SC fate in vitro. 86 However, several of these characteristics are altered during the production of transferable dECM, 70 which means that current evaluation of the effects of ECM reconstruction in 2D culture systems does not take its physical properties into account. Meanwhile, when evaluating the role of ECM remodeling in 3D hydrogels, current studies only attribute these effects to the degradation of matrix or the deposition of nascent proteins, while modifications in physical properties of hydrogels caused by these changes are neglected, which need further investigations.
Future directions
ECM remodeling is a dynamic process and is essential for maintaining SC function ex vivo. However, no system has yet been established to evaluate and regulate ECM remodeling in vitro. The following technologies need to be applied to provide a more thorough understanding and regulation of in vitro ECM remodeling.
Evaluation the effect of ECM remodeling in coculture
Coculture is the direct or indirect culture of several different cell types within the same culture conditions, with the goal of controlling cell function and behavior by facilitating the interaction of various cell types. Target cells and assisting cells are the two types of cells within a coculture system. Generally, target cells are those that will ultimately make up the engineered tissue and regulate the tissue's function. Assisting cells on the other hand, can induce target cells to exhibit desired functions, such as differentiation or proliferation and matrix organization or production. Tissue engineering coculture systems are often used to drive differentiation of SCs toward specific cell lineages by cocultures of terminally differentiated cells with SCs. 96 For instance, chondrocytes can induce differentiation of multipotent MSCs to chondrocytes without an inducer. 97 These studies usually attribute the mechanisms of coculture-induced differentiation to paracrine-signaling molecules or direct cell–cell contact. However, despite the importance of ECM remodeling in preserving SC function, the impact of ECM remodeling in coculture systems has largely been overlooked. The evaluation of ECM remodeling within coculture systems to identify key factors controlling SC function may be beneficial to improving SC culture in vitro.
Decellularization of 3D culture systems
Decellularization technology is to remove all cellular components from organs or tissues while preserving the microscopic and macroscopic anatomy of the ECM. 98 The latest advances in decellularizing tissues provide an extra research tool to assess the effect of pathological ECM remodeling on cell function and disease progression. For example, decellularized lungs from patients with chronic obstructive pulmonary disease did not support the long-term survival of lung fibroblasts, MSCs, endothelial progenitor cells, or bronchial epithelial cells, according to Wagner et al. 99 But when evaluating the role of ECM remodeling in 3D culture systems, decellularization technology has largely been ignored. We are of the opinion that it is necessary to develop decellularization technology for 3D culture systems. Decellularization and recellularization of matrix at different culture time points combined with scRNA-seq and lineage tracing can provide a more in-depth analysis of the effect of ECM remodeling on SC fate and the dynamic ECM remodeling process of SCs cultured in vitro.
Four-dimensional bioprinting
Four-dimensional (4D) bioprinting is a technology that has emerged in recent years. It combines time with 3D bioprinting as the fourth dimension. When external stimuli are applied, or when cell fusion or postprinting self-assembly occurs, the printed object can change its shape or function. 100 The 4D bioprinting can achieve dynamic changes instead of maintaining a static state with 3D printing, which makes it attractive in regenerative medicine and tissue engineering, for it can better fit the body's physiological features. The emergence of 4D bioprinting may provide a way to artificially modulate in vitro ECM remodeling. Different types of 4D self-morphing culture substrates can be used to maintain the function of SCs according to different requirements. Recently, a novel 4D substrate was developed and manufactured by Miao et al. to reproduce ECM remodeling of neural stem cell (NSC) and neural cells at various differentiation stages. 101 They demonstrated that the time-dependent self-morphing 4D culture substrate significantly improved neuronal differentiation of NSCs along with achieving significant axonal alignment.
4D culture systems can create a unique and dynamic 3D environment for SCs' biology research and can also generate a dynamic physiological environment to induce SC-based tissue regeneration.
Conclusions
Cells remodel their pericellular matrix by traction, matrix degradation, and ECM protein deposition. For SCs cultured in vitro, remodeling of the matrix is needed both to preserve its stemness and to allow differentiation. Many factors such as different culture media or physical properties of hydrogels may not directly regulate SCs, but rather affect ECM remodeling, which in turn changes the fate of SCs. These results thus suggest that ECM remodeling can be a potential regulatory target for ex vivo SCs culture. Much more effort should be focused on exploring the role of ECM remodeling in ex vivo SCs culture.
Footnotes
Acknowledgment
The figure used in this review was created with BioRender.com.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by National key R&D program of China (2017YFA0104900), NSFC grants (81874019, 81572115, 81572157), 325 Health High Level Talent project of Zhejiang province, Clinical top young talents cultivation project of Zhejiang university, the Fundamental Research Funds for the Central Universities.