
Abstract
Regenerative medicine based on stem cell research has the potential to provide advanced health care for human beings. Recent studies demonstrate that stem cells in human urine can serve as an excellent source of graft cells for regenerative therapy, mainly due to simple, low-cost, and noninvasive cell isolation. These cells, termed human urine-derived stem cells (USCs), are highly expandable and can differentiate into various cell lineages. They share many biological properties with mesenchymal stem cells, such as potent paracrine effects and immunomodulation ability. The advantage of USCs has motivated researchers to explore their applications in regenerative medicine, including genitourinary regeneration, musculoskeletal repair, skin wound healing, and disease treatment. Although USCs have showed many positive outcomes in preclinical studies, and although the possible applications of USCs for animal therapy have been reported, many issues need to be addressed before clinical translation. This article provides a comprehensive review of USC biology and recent advances in their application for tissue regeneration. Challenges in the clinical translation of USC-based therapy are also discussed.
Impact statement
Recently, stem cells isolated from urine, referred to as urine-derived stem cells (USCs), have gained much interest in the field of regenerative medicine. Many advantages of human USCs have been found for cell-based therapy: (i) the cell isolation procedure is simple and low cost; (ii) they have remarkable proliferation ability, multidifferentiation potential, and paracrine effects; and (iii) they facilitate tissue regeneration in many animal models. With the hope to facilitate the development of USC-based therapy, we describe the current understanding of USC biology, summarize recent advances in their applications, and discuss future challenges in clinical translation.
Introduction
Regenerative medicine has made tremendous progress in repairing diseased or injured organs. Stem cells represent a common choice of graft cells in the field of regenerative medicine because of the robust proliferation ability and multidifferentiation potency. Considering that each tissue has resident stem cells that are critical for tissue homeostasis, stem cells from adult tissues are widely regarded as a more readily available cell source for therapeutic applications than embryonic stem cells and induced pluripotent stem cells (iPSCs). For instance, mesenchymal stem/stromal cells (MSCs) from adult tissues have achieved some success in clinical applications such as cartilage repair and chronic wound healing.1,2 They can actively sense and respond to tissue injuries, acting as a paracrine secretor, immune modulator, and trophic player in facilitating tissue repair. 3
However, in most tissues, the abundance of MSCs is very low, and invasive procedures are usually required for cell isolation. 4 From a clinical point of view, an ideal cell source should be ethically acceptable, noninvasive, and easy-to-culture. In this context, human urine represents an excellent candidate for cell harvest since it is abundant in raw material availability, noninvasive in sample collection, and free of ethical problems.
Besides being composed of metabolic waste by-products, urine contains various types of viable cells. Recently, adult stem cells isolated from urine, termed urine-derived stem cells (USCs), have attracted growing interest as they possess many desirable properties for cell-based therapy, particularly high proliferative activity, multidifferentiation potential, potent immunomodulation ability, and strong paracrine effects.5–7 Different cell types in the urine, including USCs,8–10 renal epithelial cells,11–14 and renal tubular cells,15–17 have been used as the source of somatic cells to generate iPSCs. Indeed, iPSCs produced from urine cells, termed U-iPSCs, have great potential applications for personalized medicine, disease modeling, and drug screening (for detailed reviews, please refer to Refs.18–23 ).
There are several excellent review articles about stem cells derived from urine,24–28 most of which discussed both USCs and U-iPSCs. Unlike these reviews published previously, we just focus on USCs, but not U-iPSCs, in this article and aim to provide a comprehensive and up-to-date review of USCs and their broad applications for tissue repair and regeneration. To achieve this, we will summarize the current understanding of USC biology, review the state-of-art applications for tissue repair and regeneration, and discuss future challenges in USC-based therapy.
An Overview of USC Biology
Cell origin
The tissue origin of stem cells has significant impact on their biology, such as proliferation ability, differentiation potential, paracrine function, and molecular identity.29–32 Thus, the influences of tissue origin should be considered when designing the potential biomedical applications of stem cells or their derivates. 32 Because of the lack of specific markers, the exact origin of USCs remains unclear; however, some evidence indicates that they come from renal glomerulus.6,33
First, compared with voided urine-derived USCs, a similar biology was noted in the USCs cultured from urine obtained from the upper urinary tract, 33 which implies that voided USCs may originate from the upper urinary tract. Furthermore, there are compelling evidence to support the hypothesis that voided USCs originate from the kidneys. First, in a female recipient of a male kidney, Y chromosome was detected in the USCs, which indicated that the USCs came from the male donor's organs, namely the transplanted kidney, the renal pelvic, or the upper segment of grafted ureter. 6 Importantly, these USCs expressed markers (CD146 and podocin) being positive in parietal cells and podocytes, but negative in the mesothelial cells of the ureteral tract. Collectively, these observations suggest that USCs possibly originated from the kidney, especially the translational cells at the parietal cell/podocyte interface in the renal glomerulus. 6
Based on the expression of renal cell markers in vitro, we observed that different USC subpopulations may originate from different parts of the nephron. 34 However, more evidence is needed to confirm the exact origin of USCs in vivo. In view that USCs may come from different tissue origins, it is valuable to characterize the biology of different USC subpopulations by using the signal-cell clonal culture method. Obviously, understanding the differences between USC subpopulations will provide valuable information for the choice of suitable cell source for disease treatment, such as whether or not to use a specific USC subpopulation.
Genetic stability
Due to the low frequency of USCs in human urine, in vitro expansion is necessary to obtain a sufficient number of cells for therapy, but this procedure increases the risk of genetic alterations and cell transformation. Genetic stability is a critical safety concern for stem cell-based therapy, because genomic instability, such as chromosomal alterations, is associated with increased tumorigenicity. 35 Conventional karyotyping has been utilized to analyze the chromosomal stability of USCs.5–7,36–39 It was reported that USCs maintained a normal karyotype after serial propagation.5,7,37 For instance, Kang et al. demonstrated that USCs at passage 3, 5, and 7 possessed normal diploid complement of autosomes and sex chromosomes, and no chromosomal aberrations were recorded at these passages. 7
Telomeres are specialized nucleoprotein complexes at the end of eukaryotic chromosomes, which protect chromosomes from deletion, fusion, and aberrant rearrangements. Gradual attrition of telomeres during cell replication causes cellular senescence and apoptosis. In cells with self-renewing ability, telomere length is important for their sustained or unlimited proliferation. Telomerase is a ribonucleoprotein enzyme that catalyzes the addition of telomere to the ends of chromosomes. To exclude potential tumorigenicity of cells, assessment of their telomerase activity (low or undetectable) is generally recommended. In USCs, the robust cell proliferation ability is associated with their long telomeres.6,38,39 Interestingly, some USC clones at early passages expressed detectable levels of telomerase activity, and their proliferation capability was higher than those without telomerase activity. 39 Nevertheless, the telomerase activity decreased gradually during cell subculture. Therefore, despite the existence of detectable telomerase activity in some USC clones, USCs did not acquire tumorigenicity after expansion, which is evidenced by in vivo studies. 36
For future clinical applications, the risk of tumorigenicity of USCs should be rigorously assessed. Although several in vitro tests (e.g., karyotyping, telomere length analysis, and telomerase activity assay) have been performed, respectively, a combination of in vitro tests and in vivo studies is proposed for tumorigenicity analysis, since in vitro assessments are indirect tests, which cannot guarantee the lack of ability to form tumors in vivo. Transplantation of stem cells into athymic mice, either subcutaneously or ectopically, remains a classic method for the tumorigenicity analysis of cells in vivo. According to previous reports, USCs did not form teratomas when subcutaneously implanted in nude mice or grafted into the subcapsular space of kidneys,6,36 which provides a convenient proof-of-principle about their tumorigenicity.
Cell isolation and expansion
Human urine sediments contain live cells, which were first successfully cultured in 1972. 40 First reported by Zhang et al. in 2008, USCs were characterized as a subpopulation of urine cells that possessed progenitor cell characteristics (Fig. 1). 5 These cells were cultured by a simple and cost-effective method, including urine centrifugation and subsequent cell culture in a mixed medium composed of keratinocyte serum-free medium, progenitor cell medium, and other supplements (growth factors, bovine pituitary extract, etc.).5,33,36,41,42 To avoid microorganism contamination, sterile urine samples obtained from the mid-stream and last stream of spontaneously voided urine, or collected by ureteral catheter under sterile conditions are commonly employed for cell isolation. Furthermore, penicillin and streptomycin can be added to the collected urine to minimize contamination.

A brief history of the discovery of USCs and their applications in regenerative medicine. The left timeline depicts the discovery of USCs, and the right timeline describes the applications of USCs in tissue repair and regeneration. Due to space limitations, only a selection of important events was presented, such as the first usage of USCs for the repair of different tissues. SUI, stress urinary incontinence; USC, urine-derived stem cell. Color images are available online.
Besides the mixed cell culture medium, other growth media, such as Dulbecco's modified Eagle's medium, have been used in some reports.8,34,37,43–45 Using Matrigel as the coating material for cell culture flasks, and through the addition of a rho-associated protein kinase inhibitor Y-27632, Kim et al. have developed an improved method for the isolation and expansion of USCs, which showed significant improvement in the migration, colony forming ability, and osteogenic or chondrogenic potential of cells. 8 Although different cell culture conditions (e.g., cell culture medium, surface coating of cell culture flasks, etc.) have been reported, whether these differences have a significant impact on the therapeutic potential of USCs remains largely unknown. Particularly, the lack of standards for the characterization of USCs hinders a direct comparison among different studies, which should be addressed in the future.
In addition to freshly voided urine, samples for USC isolation can be collected by ureteral catheterization and stored in different conditions.5,38 Apart from healthy donors, USCs from patients with different diseases (e.g., bladder cancer) have been characterized.36,46,47 In the primary culture, USCs showed a rice grain-like or spindle-shaped morphology.5,34 Because of long telomeres, USCs can proliferate extensively, even with 60–70 population doublings. 6 From a clinical perspective, it is necessary to determine the alterations of USC biology during serial subculture, which is prerequisite for the determination of suitable cell passages for therapy.
Donor age, gender, and sample harvest methods also influence the isolation and growth of USCs.5,46,48 Compared to older donors, young individuals showed higher viability, better proliferation, and lower senescence in their USCs. 48 In male donors, the efficiency of USC isolation was higher than that in female donors. 46 Compared with spontaneously voided urine, urine collected by catheterization contained more abundant USCs as the catheterization procedure may have caused the scraping of cells from the inner bladder wall. 5
Surface markers
Concerning cell surface marker expression, previous researches have reported that USCs and MSCs expressed a very similar repertoire of surface markers. Typically, they are positive for CD29, CD44, CD73, CD90, CD105, CD106, CD144, and CD166, but negative for hematopoietic stem cell markers (CD11b, CD14, CD19, CD34, and CD45), endothelial cell markers (CD31), and human leukocyte antigen DR.5–7,33,36,37,41,42 Furthermore, USCs express some markers for pericytes, such as CD146 and neural/glial antigen 2.6,38 In particular, several pluripotent stem cell markers are variably expressed in USCs, such as Oct 3/4, c-Myc, SSEA-1/4, and Klf-4.5,6,33,36 During the expansion of USCs, the expression of some markers changes. Zhang et al. reported that the proportion of CD44-positive USCs decreased with increased cell passage, while the expression of CD133 increased during the subculture of USCs. 5 As the specific markers for USCs remain unknown, a combination of cell surface markers is widely used for USC characterization.
Differentiation potential
As shown in Figure 2, USCs can give rise to many cells of three germinal lineages. Following induction, they express specific markers of target cells. Differentiated USCs can generate functional tissues in vivo, such as bone, urothelium, and smooth muscle.49–51 Unfortunately, the molecular mechanisms underlying the lineage-specific differentiation of USCs, such as the signaling pathways involved, are not well understood.
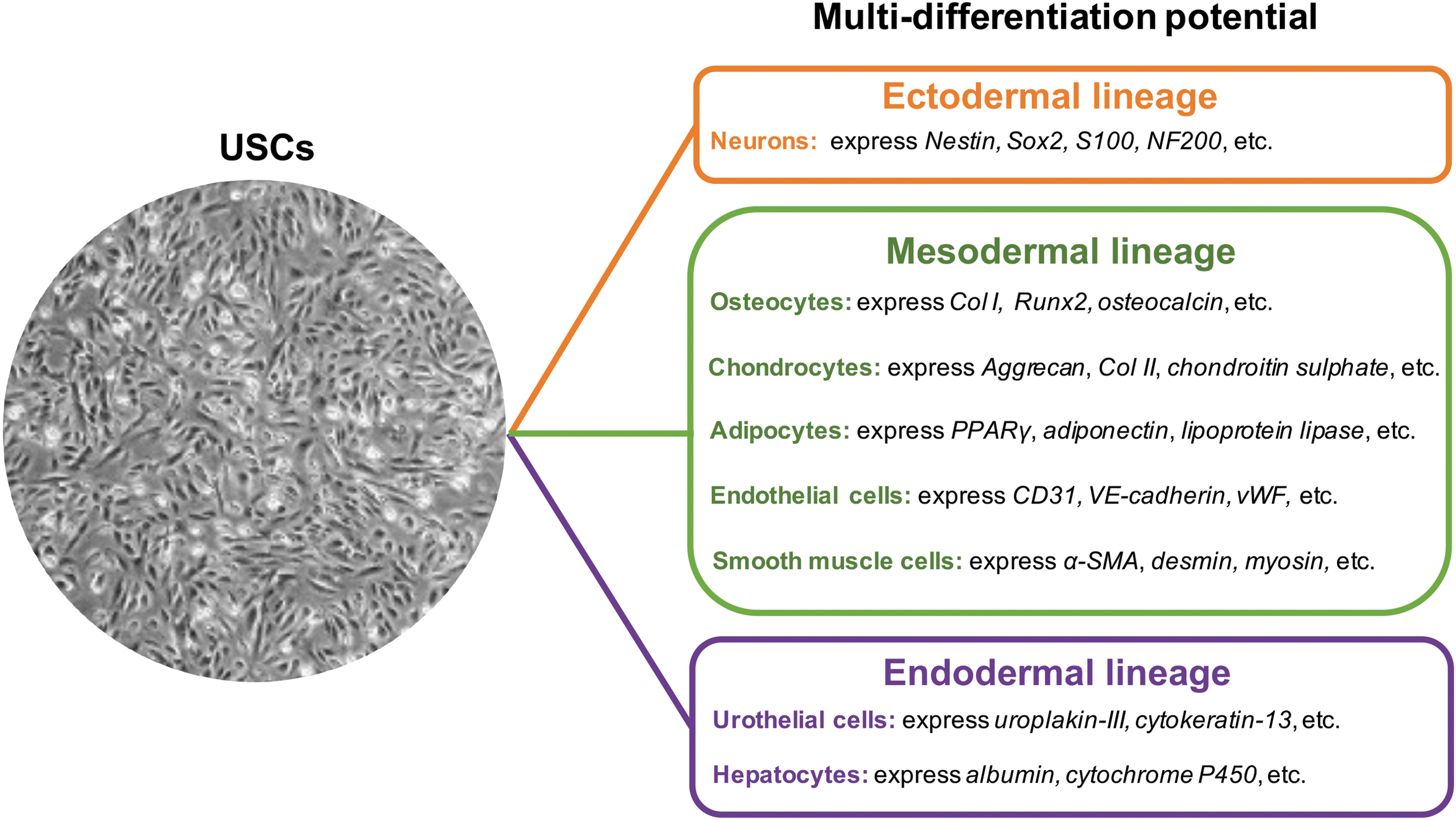
The differentiation potential of USCs. After induction, USCs can differentiate into various cell types of three germinal layers. α-SMA, alpha-smooth muscle actin; Col I, collagen type I, Col II, collagen type II; PPARγ, peroxisome proliferator-activated receptor-gamma; Runx2, runt-related transcription factor-2; vWF, von Willebrand factor. Color images are available online.
Urothelial differentiation
When induced with urothelial differentiation medium, USCs express urothelial-specific markers, such as uroplakin-III and cytokeratin-13.6,33,38 An obvious change in cell morphology was observed during the differentiation process, which typically results in a cuboidal shape. After induction, urothelial-induced USCs express proteins associated with tight junctions between the cells (e.g., E-cadherin and cingulin), show tight junction ultra-structures, and possess a barrier function similar to that of native bladder cells.6,38 When grafted with scaffolds, urothelial-differentiated USCs can generate stratified layers positive for urothelial markers in vivo. 6
Myogenic differentiation
When cultured in a myogenic medium, USCs express smooth muscle-specific markers such as alpha-smooth muscle actin, desmin, and myosin.6,33,38 The expression of these markers increases during the differentiation process. In a cell-populated collagen lattice contractility assay, myogenic-differentiated USCs showed contractile properties similar to that of healthy smooth muscle cells. 6 When subcutaneously implanted in nude mice, myogenically differentiated USCs can generate multiple layers of smooth muscle cells. 6
Endothelial differentiation
Incubated in an endothelial induction medium, USCs can efficiently differentiate into functional endothelial cells, which show a cobblestone-like morphology, express endothelial-specific markers (CD31, von Willebrand factor, and VE-cadherin), form tubular networks on Matrigel,6,52 and develop tight junctions similar to that of native endothelial cells. 52 In particular, the permeability barrier function of USCs increases significantly after differentiation. 52 When subcutaneously implanted in nude mice, endothelial differentiated USCs can form new vessels in vivo. 52
Osteogenic differentiation
After osteogenic differentiation, USCs can produce a mineralized extracellular matrix.6,48,53 The activity of alkaline phosphatase and the expression of osteogenic genes (collagen type I, runt-related transcription factor-2, osteocalcin, etc.) will increase during the osteogenic commitment of cells. Interestingly, the osteogenic potential of USCs is significantly influenced by donor age, with USCs from young children possessing stronger osteogenic capacity than those from older donors. 48 In addition, the activation of osteogenic signaling pathways, such as the Wnt/β-catenin pathway, can promote the osteogenesis of USCs.6,53,54
Chondrogenic differentiation
Using a high-density cell plate method or a cell pellet culture system, the chondrogenic potential of USCs was usually evaluated by induction in a chondrogenic medium.6,7,37,43,55,56 Successful differentiation was determined by chondrogenic gene expression and a specific extracellular matrix formation. Some studies reported that USC pellets can undergo chondrogenic differentiation,37,43,57 while others found that the chondrogenic potential of USCs was poor.6,55,58 Compared with adipose stem cells (ASCs) and bone marrow MSCs, USCs showed lower chondrogenic potential.7,56,57 However, the extracellular matrix deposited by USCs can enhance the chondrogenic potential of bone marrow MSCs, which might be due to the trophic factors released within the extracellular matrix. 55
Adipogenic differentiation
The adipogenic potential of USCs is usually determined by culturing the cells in an adipogenic medium for 2–4 weeks. Oil red O staining is typically used to detect the formation of lipid droplets in the cytoplasm. During adipogenic differentiation, USCs transform their morphologies into a cuboid shape and upregulate the expression of adipogenic genes, including peroxisome proliferator-activated receptor-gamma, adiponectin, and lipoprotein lipase.6,37 Induced USCs will form tiny lipid droplets in the cytoplasm, and some of the droplets merge to form larger ones. During in vitro expansion, USCs can maintain their adipogenic potential for seven passages. 59 However, some variations are noted in different USC subpopulations. For instance, USCs with a rice-shaped morphology possessed inferior adipogenic potential compared to those with a spindle shape. 34
Neurogenic differentiation
Cultured in a neural induction medium, USCs can change their shape into a neuron-specific morphology that contains distinguished neurogenic extensions.6,7 Furthermore, the induced USCs express neuronal markers, such as Nestin, Sox2, S100, MAP2, NF200, and GFAP.6,37,60
Other differentiation potentials
Besides the differentiation potential mentioned above, USCs can differentiate into renal cells,61–63 interstitial cells of Cajal-like cells, 64 insulin-producing cells, 65 and hepatocyte-like cells. 66 When cultured with a kidney extracellular matrix, USCs generate organoids that are similar to real renal organoids.62,63 After cultured in a kidney differentiation medium, USCs form cell aggregates that positively express renal genes (PAX2, WT1, and NPHS1). 61 Using a gene transfection method, Sun et al. successfully produced USC-derived interstitial cells of Cajal-like cells. 64 Hwang et al. reported that USCs can differentiate into insulin-producing cells that secrete insulin upon glucose stimulation. 65 When co-cultured with hepatocytes, USCs can differentiate into hepatocyte-like cells. 66
Paracrine effects
With potent paracrine effects, USCs can secrete a wide array of paracrine factors, such as growth factors, cytokines, and extracellular vesicles (EVs).61,67 The USC secretome has attracted much interest in tissue repair. In a rat model of age-related kidney insufficiency, human USCs can inhibit the inflammation and fibrosis of kidney through their paracrine effects. 68 In vitro, the trophic factors of USCs can promote the chondrogenic potential of bone marrow MSCs. 55 When cultured with periodontal ligament stem cells (PDLSCs), USCs can enhance the osteogenic differentiation of PDLSCs through paracrine effects. 69
Gene transfection has been applied to modify USC secretion. After subcutaneous implantation, USCs that overexpressed the vascular endothelial growth factor (VEGF) gene resulted in more vascularization than those without genetic modification. 70 In type 2 diabetic rats, the transplantation of USCs transfected with the fibroblast growth factor 2 (FGF2) gene obviously improved the erectile dysfunction of rats through the paracrine effects. 67 Similarly, in a rat model of cavernous nerve injury-induced erectile dysfunction, USCs transfected with the pigment epithelium-derived factor gene significantly improved the erectile function through their paracrine effects. 71
As one of the key paracrine mediators, EVs contain diverse bioactive cargos. 72 The promising prospect of USC-derived EVs (USC-EVs) has been demonstrated in animal studies.44,73–75 For instance, in a murine model of severe hind-limb ischemia, USC-EVs markedly enhanced the angiogenesis and muscle regeneration of ischemic limbs. 44 Also, USC-EVs can ameliorate diabetic erectile dysfunction (DED), which may be due to the microRNAs enriched in them. 73 Through the transference of proteins with proangiogenic and antiapoptotic effects, USC-EVs can efficiently attenuate glucocorticoid-induced osteonecrosis of the femoral head. 74 In osteoporotic mice, USC-EVs can effectively alleviate bone loss by transferring function proteins. 75
The exosomes secreted by USCs (USC-Exos) are also effective at protecting tissues against injuries such as diabetic wounds, 76 muscle injury, 77 diabetic nephropathy,78,79 renal injury,80,81 ischemic stroke, 82 intervertebral disc degeneration,83,84 and spinal cord injury. 85 For instance, in the skin wounds of diabetic mice, USC-Exos enhanced the activities of repair cells. 76 In a rat model of diabetic nephropathy, USC-Exos can prevent kidney injury through the inhibition of cell apoptosis in podocytes and tubular epithelial cells. 78
Immunoregulation
In vitro, USCs have been observed to possess strong immunomodulatory effects.7,59,61,86–88 When cultured with phytohemagglutinin-stimulated peripheral blood mononuclear cells (PBMNCs), USCs effectively inhibited the proliferation of PBMNCs. 59 When stimulated by allogeneic PBMNCs, USCs can elevate the secretion of immunoregulatory cytokines to inhibit the proliferation of PBMNCs. The immunomodulatory effect of USCs was similar to or better compared with bone marrow MSCs.86,87 Compared with ASCs, USCs are more efficient at inhibiting immune cell activation. 7 Besides the function of suppressing T cell response to activation, USC-EVs induced the proliferation of B cells and the secretion of IgM in vitro, which provide a potential tool to promote the response of B cells in immunodeficiency or cancer. 88
In animal studies, USCs have been employed to treat inflammatory diseases, such as inflammatory bowel disease, 87 interstitial cystitis, 89 and renal ischemia/reperfusion injury.80,81 In a murine colitis model, systemic administration of USCs ameliorated the severity of colitis and reduced inflammation. 87 In a rodent model of interstitial cystitis, USCs were shown to restore bladder function by inhibiting inflammation. 89 Furthermore, in acute kidney injury (AKI) models,80,81,90 chronic kidney disease models,91,92 and aging-related renal insufficiency models, 68 USCs have been shown to protect renal function, alleviate tissue damage, and promote kidney repair partially through the inhibition of inflammation in the kidneys.
USCs Versus Other Stem Cells
To provide a useful guide for seed cell selection, several studies have compared USCs with other adult stem cells.7,37,56,61 According to the International Society for Cellular Therapy, a minimal set of standard criteria has been proposed to define human MSCs, which includes (i) the cells are plastic adherent in vitro; (ii) they positively express CD105, CD73, and CD90, and negatively express CD45, CD34, CD14, CD19, and HLA-DR; and (iii) they have the ability to differentiate to osteoblasts, adipocytes, and chondroblasts after induction in vitro. 93 Although many biological properties of USCs are similar to those of MSCs (e.g., plastic adherent, negative expression of CD45, CD34, and HLA-DR, and the osteogenic, chondrogenic, and adipogenic potential of cells), 6 some differences have been identified.7,56,86 For instance, when compared with bone marrow MSCs, USCs possess higher expandability.7,56,57 The immune tolerance and immunomodulatory effect of USCs were better than those of bone marrow MSCs and ASCs.7,86 Compared with ASCs, USCs from the same donor had higher myogenic, neurogenic, and endogenic differentiation potential, but lower osteogenic and chondrogenic potential. 7 Compared with bone marrow- and placenta decidua basalis-derived MSCs, USCs showed better endothelial and vascularization potential, 56 but their osteogenic and chondrogenic potential was inferior to that of bone marrow MSCs.56,57
There are some inconsistencies regarding the adipogenic potential of USCs. Guan et al. reported that USCs had a similar adipogenic potential as ASCs, 37 while Kang et al. observed that the adipogenic potential of USCs was inferior to that of ASCs. 7 In terms of renal differentiation, USCs possessed a similar ability to that of ASCs; however, they secreted more growth factors during the renal differentiation. 61 Taken together, when selecting seed cells, subtle differences among USCs and other stem cells should be taken into consideration.
USCs in Tissue Repair and Regeneration and Their Roles
Roles of USCs in tissue repair and regeneration
Cell engraftment and differentiation have been regarded as the main repair mechanism in stem cell-based therapy. However, many studies have revealed that the rates of survival and differentiation of exogenous stem cells in target tissues were too low to explain the therapeutic effects.67,80,94,95 It was suggested that the paracrine effects of stem cells, rather than their differentiation and replacement of injured cells, play a major role in the healing mechanism.80,95,96 This hypothesis has been supported by an increasing number of studies, in which stem cell-conditioned medium or exosomes possessed a similar repair ability to their cell counterpart.80,97,98
After transplantation, USCs have multifaceted roles in tissue regeneration (Fig. 3). First, they can modulate the inflammatory responses. When implanted in an inflammatory environment, USCs can secrete anti-inflammatory cytokines and downregulate the local levels of proinflammatory factors to enhance cell survival and improve their repair ability.68,80,99,100 Treatments with USCs or their secretion protect the survival of tissue-specific cells in vitro, mainly through the reduction of cell apoptosis in an unfavored environment.78,79,90,101 Notably, USCs or their secretion effectively inhibits cell apoptosis in injured tissues.71,89,95,99

The roles of USCs in tissue repair and regeneration. USCs enhance tissue regeneration through multiple mechanisms. In most cases, only a small portion of USCs can transdifferentiate into functioning cells in injured organs to replace damaged or dead cells. Besides cell replacement, USCs promote tissue regeneration mainly by their paracrine effects to modulate local inflammation, recruit repair cells, enhance angiogenesis, inhibit cell apoptosis, promote the survival of resident cells, and regulate the remodeling of extracellular matrix. However, the roles of USCs involved in tissue repair are not equivalent, and importantly, the USCs can adapt their therapeutic effects based on the local microenvironment. Color images are available online.
Angiogenesis is critical for tissue repair because it brings essential nutrients and oxygen to cells. Many studies have demonstrated that USCs improved new blood vessel formation in ischemic tissues, either by direct differentiation or the release of angiogenic factors.43,44,67,102,103 Besides promoting angiogenesis, the secretion of USCs can stimulate the repair function of resident cells in injured tissues. 103 It has been observed that USCs or their conditioned medium can enhance the remodeling of extracellular matrix, which is beneficial for achieving an improved repair outcome.94,103
USCs for Tissue Repair and Regeneration
Due to the ability to facilitate tissue repair and regeneration, USCs have been investigated for tissue engineering and disease treatments, such as bladder regeneration, 104 skin wound healing, 43 and neural disorder treatment. 105 Although the full potential of USCs in tissue regeneration remains to be explored, the outcomes of animal studies are encouraging (Fig. 1).
Genitourinary repair and regeneration
Originating from the urinary tract system, USCs provide a good option for the repair of genitourinary organs. They can differentiate into urothelial,6,33,38 endothelial,6,52 and smooth muscle cells.6,33,38 Particularly, they can secrete bioactive factors to facilitate tissue repair.61,67–69 Therefore, USCs have been utilized to regenerate genitourinary organs in animal studies (Table 1), such as bladder regeneration in a rat bladder reconstruction model 104 and urethra regeneration in a rabbit urethral defect model. 106 Furthermore, USCs have been used to treat urogenital diseases in rodent models (Table 1), such as rat AKI model,90,99 mouse diabetic nephropathy model, 92 rat interstitial cystitis model, 89 rat stress urinary incontinence (SUI) model, 73 and rat DED model.67,101
The Application of Urine-Derived Stem Cells in Genitourinary Repair and Regeneration
AKI, acute kidney injury; bFGF, basic fibroblast growth factor; SIS, small intestinal submucosa; USC, urine-derived stem cell.
Kidney regeneration
The transplantation of USCs or their paracrine factors has showed great promise in treating AKI.80,81,90,99 For instance, in an ischemia/reperfusion injury-induced AKI model, USCs or USC-Exos can suppress cell apoptosis, inhibit inflammation, and improve renal function.80,81,99 By transferring functional microRNAs, USC-Exos can protect the function of kidney cells that are exposed to hypoxia/reoxygenation in vitro.80,81
In a rat model of diabetic nephropathy, USCs have been found to reduce renal fibrosis, alleviate inflammation, and protect renal function through the paracrine effects. 92 In diabetic rats, USC-Exos can inhibit the apoptosis of podocytes, increase the proliferation of glomerular endothelial cells, and promote vascular regeneration. 78 In vitro, microRNAs enriched in USC-Exos, such as miR-16-5p, have a protective effect on podocytes. 79 In chronic kidney disease models, USCs significantly improved renal functions through the inhibition of inflammation, oxidative stress, and fibrosis in the renal tissues.68,91
Bladder regeneration
Bladder tissue engineering has been regarded as a promising strategy to restore bladder function in patients with severe bladder diseases, such as congenital anomalies, cancers, and end-stage bladder diseases. Urothelial, endothelial, and smooth muscle cells are required for the reconstruction of bladder mucosa, vessels, and bladder wall, respectively. In the literature, USCs have been applied for bladder tissue engineering,24,104,107 as they can differentiate into bladder-associated cells, including urothelial,6,33,108 endothelial, 6 and smooth muscle cells.6,33 In a rat bladder reconstruction model, surface-modified composite scaffolds seeded with human USCs were shown to significantly increase the bladder capacity, compliance, and regeneration of urothelium, submucosa tissue, and bladder smooth muscle.104,107 In patients with bladder cancer, USCs from upper urinary tract urine are normal and, thus, suitable for bladder regeneration. 36
When cultured on an intestinal submucosa matrix, urothelial-differentiated USCs can form multilayered urothelial-like tissues. 108 In addition, when urothelial-differentiated USCs and smooth muscle-differentiated USCs were seeded on a small intestinal submucosa (SIS) scaffold and subcutaneously implanted into nude mice, the cell-scaffold constructs could generate multilayered tissue-like structures consisting of urothelium and smooth muscle. 6 All these studies point to the promising applicability of USCs in bladder regeneration.
Besides, USCs have been used to treat bladder dysfunctions.89,100 In a rodent interstitial cystitis model, intravenous administration of USCs significantly suppressed oxidative stress, inflammation, and cell apoptosis in the bladder tissues, consequently improving the micturition function and histological construction of the bladder. 89
Urethral reconstruction
The urethra is a hollow tubular organ at the distal part of urinary tract. Urethral defects usually require reconstructive surgery, but it remains a significant challenge because of various complications post-surgery. Owing to the insufficiency of autologous tissue for urethral reconstruction, there has been enormous interest in bioengineered urethra.
Because of the ability to differentiate into urothelial and smooth muscle cells, USCs hold great promise for urethral regeneration.49,50,108 When both urothelial and smooth muscle cells differentiated from USCs were seeded on a microporous bacterial cellulose scaffold or a porous SIS scaffold, the induced USCs not only penetrated the porous matrix but also formed a multilayered urothelium on the surface of the scaffolds. When the cell-scaffold constructs were implanted into athymic mice, the differentiated USCs retained their phenotypes.49,50 These observations indicate that USCs can generate urethral tissue in vivo.
Importantly, in a rabbit urethral defect model, autologous USCs seeded on SIS scaffolds were shown to improve urethral reconstruction, which was evidenced by significant improvements in the urethral caliber, the urothelial regeneration speed, the smooth muscle content, and the vessel density. Noteworthy, grafted USCs were found to differentiate into urothelial cells and smooth muscle cells in vivo. 106
Stress urinary incontinence
The sphincter muscle controls bladder tightness, and its degeneration or the impairment of its neural or vascular support can lead to SUI, which affects millions of people, especially senior citizens. USCs are useful for SUI treatment because of the myogenic potential and the ability to promote vascularization and innervation.70,109–111 For instance, USCs overexpressing VEGF gene can improve the neovascularization and nerve regeneration of implants in vivo.70,110,111
Promising outcomes have been reported in the correction of SUI through the transplantation of USCs or their secretion products.77,112 Systemic administration of USCs or their concentrated conditioned medium can facilitate the functional recovery of SUI, which has been confirmed by increased leak point pressure and an improvement in the structure of the external urethral sphincter. 112 In addition, USC exosomes can promote the activation, proliferation, and differentiation of muscle satellite cells. 77 In a rat SUI model, USC-Exos significantly improved the urodynamic parameters and repaired the injured muscles. 77 These studies provided a sound basis for further development of USC-based therapy for SUI correction.
Erectile dysfunction
As a common male sexual disorder, erectile dysfunction affects the life quality of patients. Several studies have determined whether the transplantation of USCs provides an effective approach to improving erectile dysfunction.67,71,73,101,113 In a rat DED model, intracavernous injection of human USCs or USCs overexpressing the FGF2 gene significantly improved erectile dysfunction, resulting in increased intracavernosal pressure (ICP), a higher ratio of ICP to mean arterial pressure, and an increased expression of endothelial and smooth muscle markers in the cavernous tissue.67,101 Furthermore, USCs have been shown to ameliorate cavernosal endothelial dysfunction in DED rats. 101
Interestingly, several weeks post cell transplantation, few USCs could be detected at the implant sites. 67 Inspired by this observation, it was noted that the paracrine effect of USCs, rather than their differentiation, accounted for the improvement in erectile function. This was further confirmed by the observation that USC-EVs can ameliorate DED. 73 Furthermore, in a cavernous nerve injury-induced erectile dysfunction model, USCs were shown to protect erectile function in, and the cavernous structure of, rats through their paracrine effects. 71 Similarly, in a rat Peyronie's disease model, human USC-Exos were shown to ameliorate tunica albuginea fibrosis and improve erectile function. 113
Musculoskeletal regeneration
Skeletal muscle regeneration
In the field of skeletal muscle regeneration, USCs have been regarded as a potent and attractive source of seed cells (Table 2). Following myogenic differentiation, USCs expressed high levels of skeletal muscle markers.114,115 The phenotype of myogenically differentiated USCs was stable after implantation in vivo. 114 Overexpression of the VEGF gene can effectively enhance the survival and myogenic differentiation of USCs. 70 Besides gene transfection, local delivery of growth factors provides an alternative method to improve the survival and myogenic differentiation of USCs in vivo.109,116 This indicates that a combination of USCs, growth factors, and biomaterials is beneficial for skeletal muscle regeneration.
The Application of Urine-Derived Stem Cells in Musculoskeletal Regeneration
β-TCP, beta-tricalcium phosphate; BCP, biphasic calcium phosphate; BMP2, bone morphogenetic protein 2; CS, chitosan sponges; VEGF, vascular endothelial growth factor.
Cartilage regeneration
Because of the poor intrinsic repair ability of cartilage, stem cells have attracted considerable interest in articular cartilage repair. In 2018, Chen et al. reported, for the first time, the utilization of USCs for cartilage repair (Table 2). 117 Mixed with hyaluronic acid, USCs were injected into rabbit knee joints with cartilage defects. Twelve weeks after implantation, the transplantation of USCs combined with hyaluronic acid induced more neocartilage formation than the cell-only group and the scaffold-only group. This result indicates the possibility of USCs in cartilage regeneration. However, the underlying mechanism remains unknown and warrants further investigation.
Recently, Sun et al. compared the chondrogenic potential and cartilage repair ability of human USCs and bone marrow MSCs derived from the same individual (Table 2). 57 USCs showed inferior chondrogenic potential to that of bone marrow MSCs in vitro. However, after seeding on acellular cartilage extracellular matrix scaffolds, both USCs and bone marrow MSCs significantly promoted the cartilage repair in a rabbit critical-size cartilage defect model. In both cell types, the repair tissue was mainly hyaline cartilage, and no significant difference was noted in the repair effects. The similar cartilage repair ability of USCs indicates that they would be a new promising candidate cell source for cartilage regeneration.
Bone regeneration
Large bone defects require a large number of osteogenic cells to support bone regeneration. Because of the osteogenic potential and paracrine effects, USCs have been utilized as seed cells to construct bone grafts (Table 2). Several applicable methods have been reported to enhance the osteogenic differentiation of USCs, such as the supplementation of osteogenic material and the employment of gene transfection.51,118 Transduced with the bone morphogenetic protein 2 (BMP2) gene, USCs can undergo osteogenic differentiation even without other osteogenic supplements. 51 In the BMP2-induced osteogenic differentiation of USCs, focal adhesion kinase played a critical role. 54 Interestingly, donor age of USCs influences their osteogenic potential. Comparing with USCs derived from elderly donors, those isolated from child donors have showed a stronger osteogenic potential. 48
In a rat critical-sized bone defect model, USCs in combination with beta-tricalcium phosphate (β-TCP) scaffolds induced more bone formation than the exclusive β-TCP scaffold group. 119 Guan et al. reported that calcium silicate ion extracts could stimulate the osteogenic differentiation of USCs. 53 Similarly, in a rabbit critical-sized segmental bone defect model, surface mineralized biphasic calcium phosphate ceramics loaded with USCs yielded more new bone tissue than the scaffold-only group. 120 Besides the direct application of USCs, their paracrine factors are also promising for bone regeneration.74,75 At the early stage of glucocorticoid exposure, USC-EVs could obviously attenuate the glucocorticoid-induced osteonecrosis of the femoral head. 74 In an osteoporotic mouse model, systemic injection of USC-EVs alleviated bone loss and maintained bone strength. 75
Tendon repair
A recent study by Chen et al. has explored the usage of USCs for tendon repair (Table 2). They demonstrated that transplantation of an autogenous USC sheet could enhance the healing of rotator cuff injury in dogs. 121 The cell sheets were implanted at the defect sites of rotator cuff. Cell tracking revealed that the cell sheets were viable 6 weeks post implantation. Importantly, more enthesis-like tissue was noted in the USC sheet group, which achieved a higher score of failure load than the non-USC sheet transplantation group. This pioneer study suggested that USCs have good potential for tendon repair.
Skin wound healing
To promote skin wound healing, USCs have been utilized to fabricate tissue-engineered skin grafts (Table 3).43,94,102 In a rabbit full-thickness skin defect model, USCs in combination with a nanofibrous membrane significantly promoted the angiogenesis and healing of wounds. 43 Cao et al. evaluated the effect of a surface-structured bacterial cellulose biomaterial and human USCs, either alone or in combination, on the healing of full-thickness skin defects. They observed that a combination of the biomaterial and USCs resulted in better angiogenesis and wound healing compared to those treated with biomaterial or USCs alone. 102
The Application of Urine-Derived Stem Cells in Skin Wound Healing, Neurological Deficits, and Other Chronic Diseases
PCL/GT, polycaprolactone/gelatine.
Modulating the paracrine effects of USCs can change their wound healing potential.76,94,103 We observed that hypoxia preconditioning increased the growth factor secretion of USCs, consequently improving the wound healing ability of a tissue-engineered composite seeded with USCs. 94 When stimulated with the ionic products of bioglass, USCs can upregulate the expression of growth factors and stimulate the paracrine effects with recipient cells; when grafted in vivo, activated USCs resulted in better wound healing outcomes, such as improved angiogenesis and collagen deposition. 103 As a key mediator of paracrine actions, USC-Exos can augment the function of wound healing-related cells, promote wound angiogenesis, and enhance wound healing in diabetic mice. 76
Neural disorders
With the potential to differentiate into neuronal lineages,37,105,122 USCs have been investigated to treat neurogenic lesions (Table 3).37,82,105 When induced with specific factors, USCs are capable of neurogenic differentiation. 37 Both laminin and platelet-derived growth factor BB can promote the neuronal differentiation efficiency of USCs. 122 When mixed with a hydrogel and injected into rat brain lesions, USCs can survive, migrate, and differentiate into neuron-like cells. 37 In a rat stroke model, intravenous injection of USC-Exos enhanced neurogenesis and further promoted the functional recovery of post-ischemic stroke. 82 Similarly, in a rat model of cardiopulmonary resuscitation after cardiac arrest, USCs markedly facilitated the recovery of neurological function, probably through the secretion of neuroprotective factors and the inhibition of brain edema. 105 In a mouse Rett syndrome model, it was observed that human USC-Exos improved the behavior, cognitive ability, and motor coordination of mice. Through targeting the Eph receptor A4/TEK axis, the exosomal miR-21-5p of human USC-Exos promoted early neural formation in the mouse model. 123
Other chronic diseases
Apart from the applications described above, USCs also serve as a versatile therapeutic tool for several chronic diseases (Table 3), such as diabetes mellitus,95,124 chronic liver injury,66,125 and inflammatory bowel disease. 87
In a rat model of type 2 diabetes, administration of USCs was shown to significantly alleviate the histological destruction and functional decline of the heart, kidney, and bladder. 95 In another study, intrapancreatic transplantation of USCs prolonged the median survival time of diabetic mice and improved their glucose homeostasis through the maintenance of pancreatic beta-cell mass and the enhancement of vascular regeneration. 124 When co-cultured with human hepatocytes, USCs were shown to differentiate into hepatocyte-like cells. 66 Notably, in a mouse model of chronic liver injury, the transplantation of USCs improved abnormal liver function and repaired damaged liver tissue. 125 In addition, hypoxia preconditioning further improved the liver recovery efficiency of USCs, which was relevant to the upregulation of the C-X-C motif chemokine receptor 4 signaling. 126 In murine colitis models, systemic administration of USCs not only reduced the clinical severity of colitis but also improved the survival rate of mice. 87
Future Directions
The therapeutic potential of USCs has not been fully investigated. Before progressing into clinical applications, many issues need to be addressed. First, there are no donor selection criteria for USCs. It is important to consider the effects of diseases and medical treatments on the biology of USCs. Detailed donor criteria should be established to avoid disease transmission. Patients with severe infectious diseases should be excluded from the donor list.
Second, to increase the effectiveness and repeatability of cell isolation, standard procedures for USC isolation are recommended. Because of the toxic environment of urine, it is necessary to optimize the method of urine sample preservation to maintain the viability of USCs during prolonged sample collection and transportation. Furthermore, with growing understanding of USC biology, it is necessary to propose a minimal set of standard criteria for defining USCs, which is critical for the quality control of USCs in cell-based therapy and lays a solid foundation for direct comparison of different studies.
To establish applicable protocols for large-scale production of USCs, much work should be carried out to optimize the culture conditions, such as the choice of culture media and the establishment of a favorable microenvironment. To avoid disease transmission, xeno-free cell culture systems should be established to produce clinical-grade USCs. Before cell delivery in vivo, the quality control of USCs, including a careful check of microorganism contamination, should be analyzed.
Third, the poor survival and few engraftments of USCs after implantation should be solved to improve therapeutic efficiency. This requires applicable strategies to facilitate the short-term retention and long-term engraftment of cells, such as the use of porous biomaterials to protect cells. Extending the lifespan of grafted USCs can result in better repair outcomes because of the prolonged release of paracrine factors and the enhanced recruitment of resident cells. Further research, especially large animal studies, are required to answer important questions related to the efficiency and repair mechanisms of USCs. Advanced methods, such as omics techniques, should be used to uncover the underlying mechanisms.
So far, no clinical trial has been registered on the website of the National Institutes of Health (www.clinicaltrials.gov) to determine the safety and efficiency of USCs in disease treatment. This could be attributable to the following reasons: (i) the lack of a thorough understanding of USC biology; (ii) the lack of large animal studies about the safety and efficiency of USCs; (iii) the lack of standard protocols for USC isolation and expansion under good manufacturing practice (GMP) guidance; and (iv) the lack of suitable standards for the quality control of USCs in cell-based therapy. Furthermore, before clinical attempts, future preclinical research should focus on the suitable indications for USC transplantation, the cell doses, and the optimal delivery methods.
Although current researches mainly focus on human USCs, a few studies have isolated and characterized USCs from animals, such as those from rabbits106,127 and dogs.45,121 These animal-derived USCs have shown good repair potential for some diseases.106,121 However, unlike the simple and noninvasive urine collection from human donors, the harvest of urine from animals is complicated and usually involves invasive process, such as urethral catheterization under general anesthesia.45,121,127 Based on these reports, there is no doubt that USCs have possible applications for animal therapy, but the sample harvest method needs further improvement to minimize the damage to animals.
Conclusions
As a relatively new source of stem cells, USCs share many biological characteristics with MSCs. They are highly expandable and can maintain a normal karyotype after extensive expansion. Besides broad differentiation potential, USCs can secrete various kinds of bioactive factors to benefit tissue regeneration. In preclinical studies, USCs have showed many positive outcomes in tissue engineering and disease treatments. However, to achieve successful clinical translation, many critical issues need to be addressed, especially the large-scale production of clinical-grade USCs. With further optimization and investigation, USCs may change regenerative medicine.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by National Natural Science Foundation of China (Grant Nos. 32071331 and 31600792), and Post-Doctor Research Project, West China Hospital, Sichuan University (No. 2018HXBH053), and the 1.3.5 Project for Disciplines of Excellence, West China Hospital, Sichuan University (Grant No. ZYJC18002).