
Abstract
Mesenchymal stem cells (MSCs) have been seen for years as great candidates for treating different diseases and an alternative to embryonic stem cells due to their differentiation capacity in vitro. More recent research has focused on their ability to modulate the immune response and regeneration at sites associated with inflammation, activities attributable to the release of trophic factors into the extracellular medium, a set of components known as the secretome. It has been possible to demonstrate the presence of these cells within the tumor microenvironment, which is associated with their tropism for sites of inflammation; however, their role here needs to be clarified. In different investigations, the feasibility of using MSCs or their secretome to treat cancer has been sought, with these results being ambiguous. It has been described that MSCs can be activated and present various phenotypes, which could explain the divergence in their action; however, these activation mechanisms and the different phenotypes still need to be well known. This review explores MSCs and their use in regenerative medicine with a targeted approach to cancer.
Impact Statement
This text addresses the diverging findings on the role of mesenchymal stem cells in the tumor microenvironment and discrepancies on the use of these cells as cancer treatment, separating the direct use of the cells from the use of the secretome. Multiple authors refer equally to the cells and their secretome to conclude on the positive or negative outcome, without taking into consideration how the cells are affected by their surroundings.
Introduction
Mesenchymal stem cells (MSCs) are a heterogeneous group that has attracted the interest of researchers in regeneration due to their ability to self-renew and differentiate into multiple cell lineages. Initially, they were isolated from bone marrow; however, nowadays they are obtained from different tissues; among the most common are bone marrow, adipose tissue, and Wharton's jelly from umbilical cord.1,2 Its origin is still under debate, as are some of its in vitro behaviors; however, its immunomodulatory role could be more relevant than its differentiation capacity. 3
The immunomodulatory capacity of MSCs has led to the investigation of the MSC's secretome, which corresponds to the material secreted by these cells and can induce a paracrine response in nearby cells.4–6 This may contain growth factors, cytokines, microvesicles, and exosomes. This characteristic of MSCs has opened the doors for diverse applications, from autoimmune diseases to cardiovascular diseases, in addition to exploring cell-free treatments.6–8
Another characteristic of MSCs is their marked tropism for inflammatory sites. Although they are expected to perform their immunomodulatory role here, including the microenvironment of the tumor, which presents a particular state of inflammation, their function has yet to be fully determined. There are both reports that describe how these cells facilitate tumor development or they inhibit it through different mechanisms.9,10 Considering its paracrine role, the MSC's secretome has been studied in the context of cancer, also giving ambiguous results,11–13 and this has led some researchers to hypothesize that MSCs can be activated toward different phenotypes, including one with a protumoral character and another antitumoral,14,15 and this activation can occur in the tumor microenvironment (TME) itself, generating niches. 16 The possibility remains open to study further the effect of exposing MSCs to the TME and see if these effects can be reversed, or if using conditioned media prepared under controlled conditions is the best option.
Mesenchymal Stem Cells
MSCs were initially isolated from bone marrow and identified as a subpopulation of the latter with the ability to adhere to plastic, with osteogenic potential, and with a fibroblastic appearance, which Friedenstein et al. identified as Colony-Forming Unit-Fibroblasts (CFU-F).17–19 Between the 1970s and 1980s, in their study on the culture of chicken embryonic mesodermal cells, the scientist Arnold Caplan and his collaborators observed that these cells responded to local stimuli or signals, causing lineage progression. 20 In principle, they identified that these could differentiate into osteoblasts, chondrocytes, and adipocytes under differentiated culture conditions21,22; in addition, they propose a hypothesis about the pathway of the mesengenic process (Fig. 1) that indicates the differentiation pathways of MSCs in a wide variety of cell types. 3 Moreover, they identified the periosteum, the medulla, and the connective muscle tissue as tissues that host these cells, which they called “Mesenchymal Stem Cells.”
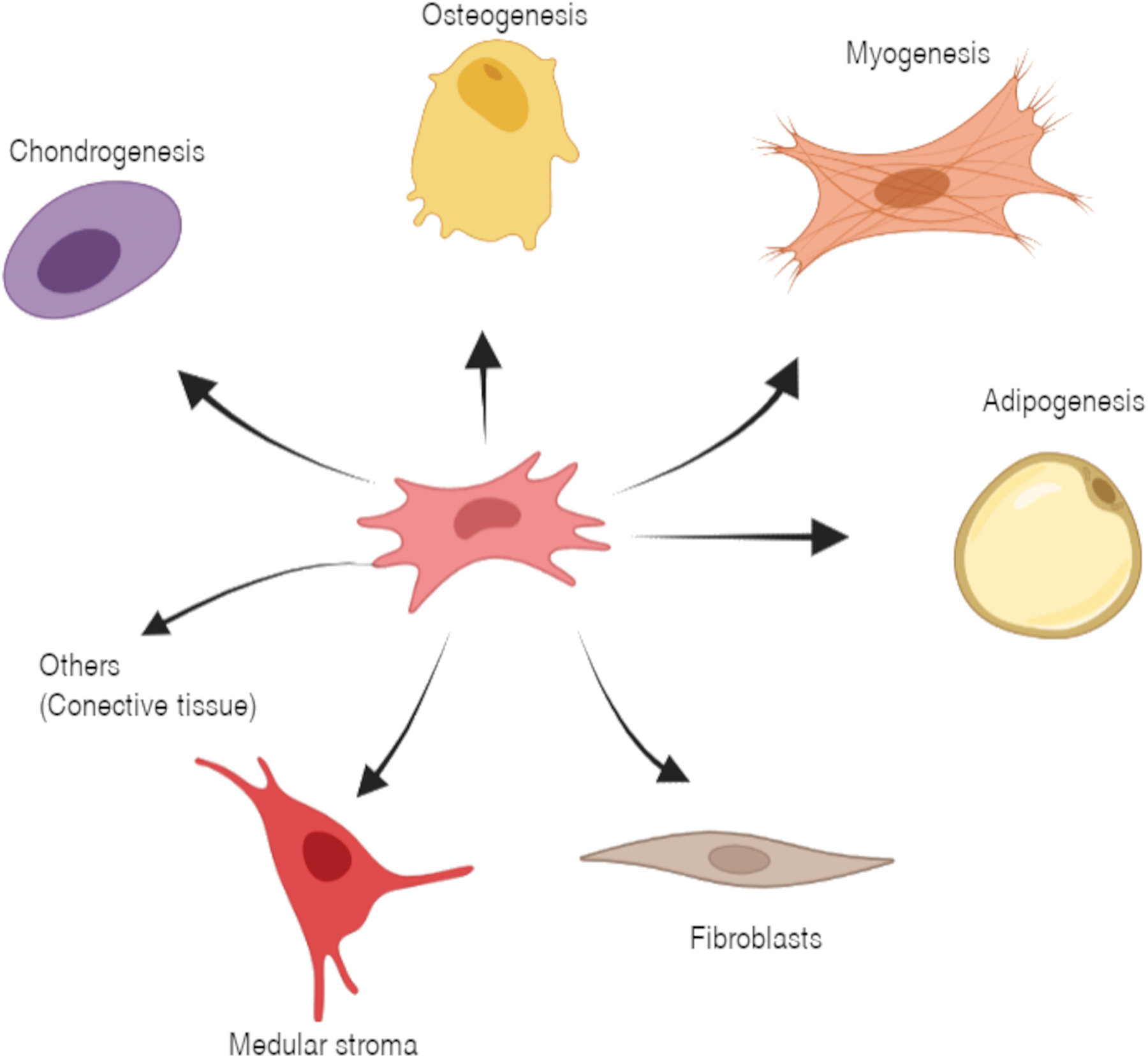
Hypothesis of the pathway of the mesengenic process tested by Caplan. (Information taken from Ref. 25 ). Figure created with BioRender.com Color images are available online.
In recent years, the sources of MSCs have been expanding, and although their embryonic origin has not been clarified, some discoveries suggest that they come from a supporting layer of the aorta–gonad–mesonephros.1,23 It has been possible to isolate populations of these cells from bone marrow (BM-MSC), adipose tissue (AMSC), umbilical cord (Wharton's jelly [WJ-MSC]), fetal liver, muscle, lungs, skin tissue among others.1,24
However, the hypothesis that MSCs derived from the differentiation of perivascular cells (pericytes) has been strengthened. It is established that these cells can be isolated from any vascularized tissue.3,25,26 Several studies on the microanatomical location of MSCs consistently reveal that they reside in the perivascular niche and share morphological characteristics and expression of surface proteins.26–29
Given these cells' great diversity, it is impossible to characterize them with a single marker. For this reason, in 2006, the International Society for Cellular Therapies (ISCT) proposed some minimum criteria for the identification and characterization of human MSCs: (1) These should be adherent to plastic when grown under standard culture conditions and form CFU-Fs; (2) must express CD105, CD73, and CD90, and lack expression of CD45, CD34, CD14 or CD11b, CD79α or CD19 and HLA-DR; and (3) must have the ability to differentiate into osteoblasts, adipocytes, and chondroblasts in vitro.1,30
However, this criterion is insufficient to explain the heterogeneity of the MSCs. Their phenotype and functionality have been found to differ between tissue source, gender, age, individuals, health status such as obesity, and even within the same MSC isolate, different profiles can be found as observed by single cell omics analyses (MSC heterogeneity is reviewed in Ref. 31 ). Zheng and collaborator analyzed >130,000 single-MSC transcriptomes from different tissues and individuals, finding that the extracellular matrix (ECM) contributes significantly to MSC heterogeneity. It is yet poorly understood how epigenetic differences and phenotypes in MSCs are linked to the local microenvironment, knowledge that can be used to engineered therapeutic MSCs or subproducts such as their secretome. Whether there is a naive MSC phenotype or not is still unclear, however, some authors have proposed that MSCs obtained from induced pluripotent stem cells are more homogeneous, also expression of miR-335 in MSCs was linked to a less differentiated state. 32
MSC Secretome
Although the in vitro differentiation capacity of MSCs has been tested, 33 the evidence that this can occur in noncultured cells is limited. The ISCT had already recommended in 2005 to use the term “Multipotent Mesenchymal Stromal Cells,” since a general criterion of their activity as stem cells was not achieved. 34 Arnold Caplan himself, who proposed the name “Mesenchymal Stem Cells,” has more recently suggested replacing it with “Medicinal Signaling Cells” 3 due to their tropism and immunomodulatory activity in response to inflammation and vascular damage.
Based on the hypothesis that MSCs are pericytes, Caplan suggests that once an inflammation domain is generated or a blood vessel is damaged, the pericytes of this tissue are released from their adluminal site, entering a state of activated MSCs (Fig. 2), which thus helps prevent the initiation of an autoimmune response, inhibits apoptosis, prevents scar formation, and stimulates angiogenesis by secreting bioactive molecules, which favors the recovery of the affected tissue.2,35
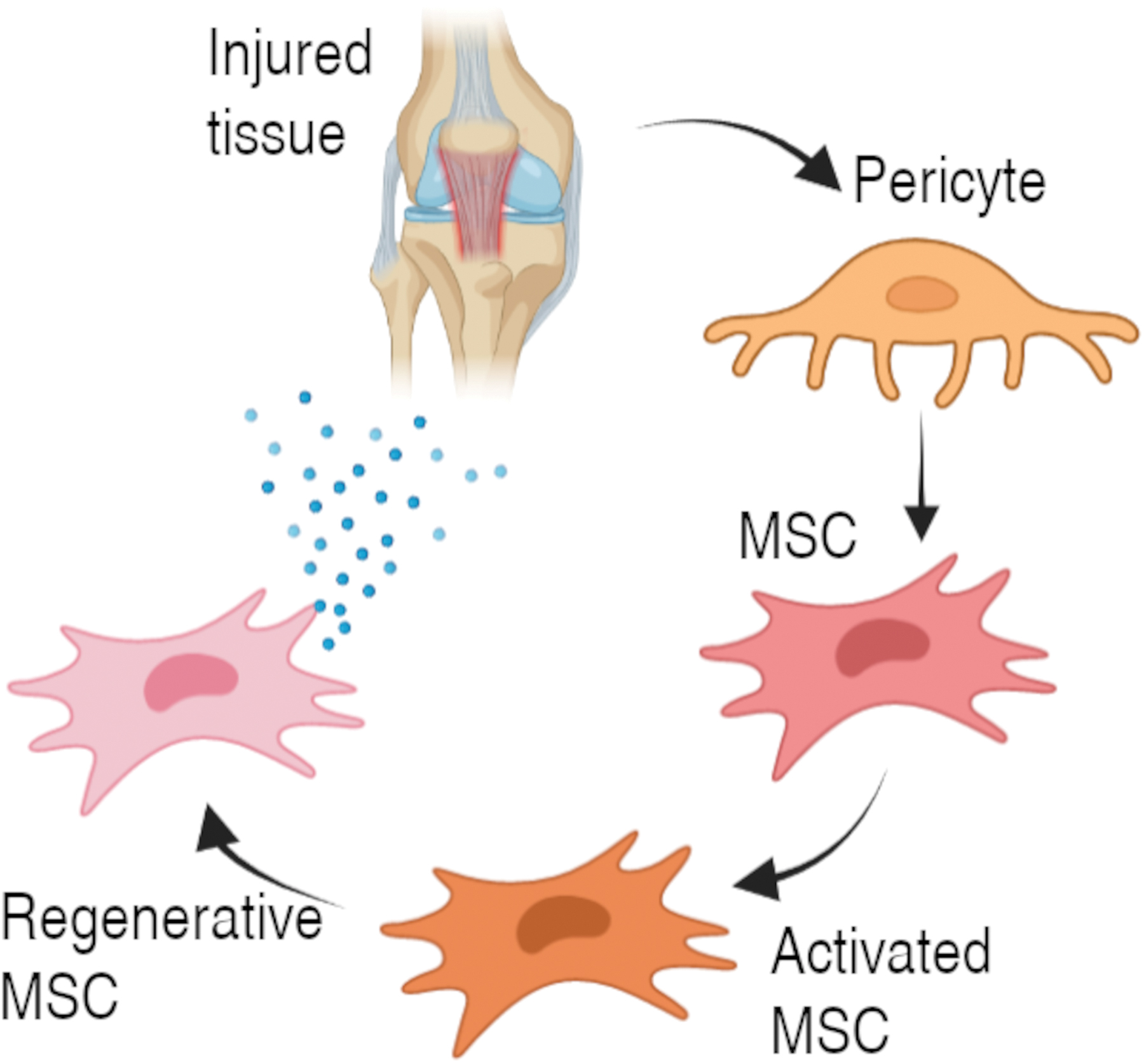
MSC activation pathway from pericytes. Information taken from Caplan 2 . Figure created with BioRender.com Color images are available online.
After the observations of the immunomodulatory effect exerted by MSCs, their secretome has gained significant interest.4,36 A wide variety of cytokines, chemokines, and growth factors considered protagonists in tissue regeneration and repair processes have been identified in it. 4 Proinflammatory cytokines, such as IL-7, IL-8, IL-9, INFγ, IL-1β, and TNF-α, have been identified in the secretome of AMSC, also anti-inflammatory cytokines, such as IL-1Ra, IL-10 and IL-13, and some bifunctional ones such as IL-2 and IL-6. 5 Proangiogenic factors, such as platelet-derived growth factor (PDGF), fibroblast growth factor (FGF), vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), and angiopoietin, have also been identified. 37 Just as the therapeutic potential of AMSC has been reported, proteins in their secretome have also been identified with possible applications for the treatment of neurodegenerative, cardiovascular, metabolic and respiratory diseases, bone regeneration, and antiaging and skin care purposes. 5
Giannasi et al. studied the conditioned medium of three donors. Among the samples, 26 analytes were found with a high degree of uniformity; 7 lipid molecules were also uniformly identified in the 3 samples by ultra-high performance liquid chromatography-mass spectrometry: arachidonic acid, eicosapentaenoic acid, docosahexaenoic acid, prostaglandin E2, prostaglandin F2α, N-palmitoylethanolamide, and N-stearoyl ethanolamide. 38
Shin et al. 39 studied the secretome of MSCs obtained from the placenta (PL-MSC-CM), Wharton's jelly (WJ-MSC-CM), bone marrow (BM-MSC-CM), and adipose tissue (AMSC-CM) by mass spectroscopy followed by a functional analysis identifying significant differences in composition; however, the functional analysis was similar. They identified 181 proteins in common; of these, 78% were predicted to be secreted. Factors related to tissue development and remodeling were found in a higher proportion in PL-MSC-CM and BM-MSC-CM while proteins related to antiapoptotic activity were found in AMSC-CM, WJ-MSC-CM, and PL-MSC-CM. In PL-MSC-CM, WJ-MSC-CM, and AMSC-CM but not BM-MSC-CM, an abundant number of proteins related to cell migration and apoptosis reduction were identified. Kehl et al. 7 also compared different MSC sources finding that WJ-MSC secretes a more complete network of proangiogenic factors and that AMSC lacks crucial angiogenic proteins, including FGF-2, AKT1, NOTCH3, and angiopoietin 1.
MSCs in Cancer Treatments
It is estimated that 9.6 million people died in 2018 from cancer, considered the second leading cause of death worldwide. About a third of deaths are due to one of the five dietary factors and habits that increase the risk of suffering from it.40,41 It is manifested by the rapid proliferation of abnormal cells, which affects the normal development of healthy tissue. 41 These cells resist cell death, allow replicative immortality, promote angiogenesis, and activate invasion and metastasis mechanisms (Fig. 3), all due to multiple genetic mutations.42,43

Tumor microenvironment and cancer hallmarks (Hanahan et al. 42 ). Figure created with BioRender.com Color images are available online.
One property of MSCs makes them of particular interest for treating different diseases: their ability to migrate to inflamed tissues,44–46 in addition to the immunomodulatory properties opens up a range of possibilities to study the effect of MSCs over the TME. It has been possible to demonstrate in different studies that MSCs are recruited by different tumors such as breast cancer, colon cancer, ovarian cancer, melanomas among others.47–50
The tumor microenvironment
In addition to the genetic and epigenetic alterations in tumor cells, tumorigenesis depends on changes in the surrounding microenvironment, both at the cellular and noncellular levels, mediated by reciprocal interactions. 51 The first observations that allowed moving from investigations focused on tumor cells to a vision of TME took place in the 80s, thanks to the investigations of Paget 52 in breast cancer.
Interactions within the TME range from cell to cell/ECM contacts to soluble molecules. 53 TME is characterized by presenting a state of chronic inflammation to which cells with different characteristics respond; among them are cells of the innate and adaptive immune system, fibroblasts, endothelial cells, pericytes, and MSCs, all supported by an active ECM. 54 The accelerated growth of tumor cells generates a hypoxic environment that causes the expression of genes such as hypoxia-inducible factor 1 (HIF-1), which in turn induces the expression of multiple genes that regulate tumor progression. 55 It also promotes angiogenesis through the recruitment of endothelial progenitor cells and pericytes. However, the tumor vasculature is usually disordered, with discontinuities in the basement membrane, causing leaks 56 ; sometimes even tubular formations of tumor epithelial cells are created (vascular mimicry). 57
Additionally, tumor cells undergo a metabolic change increasing glucose uptake and lactate production while having functional mitochondria (known as Warburg effect) facilitating biosynthesis and creating an acidic environment, that in turn, can give rise to inflammation, increased interstitial pressure, and the emergence of tumor-associated cells (fibroblast, MSC, macrophages, etc.), mainly through the TGF-β/Smad pathway (TME reviewed in Ref. 54 ). The ECM plays an important role supporting tumor growth and creating a barrier for cytotoxic drugs, but also provides biochemical and mechanical cues favoring cancer stem cell homeostasis 58 ; furthermore, ECM turnover differs from healthy tissue resulting in stiffer and fibrotic ECM and increasing hypoxia; the expression of matrix metalloproteinases (MMPs) favors the release of growth factors allocated in the ECM and also facilitates migration and metastasis.
The immune infiltrate plays a critical role in tumor development. A tumor with the presence of proinflammatory cytokines and CD8 T cells, CD4 TH1 or NK cells, considered a “hot tumor,” presents a better prognosis than one with immunosuppressive cells such as tumor-associated macrophages, myeloid suppressor cells, and regulatory T cells, “cold tumor”; however, the TME is generally considered immunosuppressive. 59 Tumor cells can develop the ability to evade the recognition and effect of T cells by expressing the PD1 ligand (PD-L1), which by binding to PD-1 on T cells, decreases their proliferation, inhibits cytokine secretion, and induces apoptosis. 60 Current cancer therapies try to target not only tumor cells but also the stroma.
MSCs in the TME
It has been proposed that MSCs can account for 0.01–5% of the TME population. 61 As mentioned, MSCs have a marked tropism for inflamed sites, a characteristic of TME, 62 possibly through mechanisms involving C-X-C chemokine receptor type 4 (CXCR4), MMP2, urokinase-type plasminogen activator along with IL-6 and IL-863,64 and CCL2, CCL5, CXCL12, and CXCL16 produced by TME cells. 65 Once in the tumor, interactions between the components of the TME, can cause clonal selection and stimulate MSC adaptation; in addition, the differentiation of MSCs into cancer-associated fibroblasts (CAFs) has been demonstrated.66,67 TGF-β has been identified as a potent inducer of a protumoral phenotype in MSCs and CAFs in several tumor types 68,69; this in turn promotes the expression and secretion of more TGF-β, which may favor the migration of more MSCs to the TME, 70 in addition to favoring the metastasis of tumor cells through the induction of the epithelial/mesenchymal transition (EMT) program. 71
MSCs also have an essential role in tumor angiogenesis by secreting proangiogenic molecules such as VEGF or angiopoietin that aid in the recruitment of endothelial progenitor cells,72,73 in addition to its ability to acquire phenotypes similar to that of endothelial cells, 74 collaborating in the formation of capillaries. 7 MSCs can also favor the presence of immune cells with an anti-inflammatory profile of both innate and adaptive immune systems, thanks to the secretion of nitric oxide, prostaglandin E2, IL- 6, IL-10, or indolamine 2,3-dioxygenase 72 in response to stimuli by proinflammatory cytokines such as IFNγ, TNF-α, and IL-1.65,75 The presence of these cells in the TME decreases the activity of cytotoxic T cells and NK cells. It has also been shown that MSCs can promote immunosuppression through the secretion of PD-L1 and PD-L2.72,76
Regarding tumor cells, different types of interactions have been reported, 77 either by direct contact or indirectly through their secretome. Among the direct interactions, gap junctions, nanotubes, and even cell fusion have been found. In a breast cancer model, the exchange of mitochondria (MitoCeption) through nanotubes was evidenced, favoring proliferation and migration due to altered metabolic activity. 78
It has also been possible to appreciate an antitumor effect exerted by the MSCs, giving rise to a controversy that is still unresolved.10,72 The ability of MSCs to polarize toward a pro- or anti-inflammatory phenotype in response to some signals has been demonstrated in a series of studies; Waterman et al. suggest the terms MSC1 and MSC2, respectively, like what occurs with macrophages. Polarization was demonstrated by activation of toll-like receptors (TLRs) using lipopolysaccharides (LPS) as TLR4 ligand and MSC1 inducer, or polynosinic acid-polycytidylic acid (Poly I:C) to stimulate TLR3 and induce MSC2. 14 In addition, they demonstrated that MSC1 could inhibit tumor growth in different tumor cell lines through the overexpression and secretion of TNF-related apoptosis-inducing ligand, leukemia inhibitory factor, granulocyte-macrophage colony-stimulating factor, and a decrease in Chemokine (C-C motif) ligand 5 (CCL5 or RANTES) and HGF, among others, while MSC2 had the opposite effect. 15 These observations have given rise to the hypothesis that MSCs located in different compartments within the same tumor with different local characteristics may favor one MSC phenotype (Fig. 4).16,72
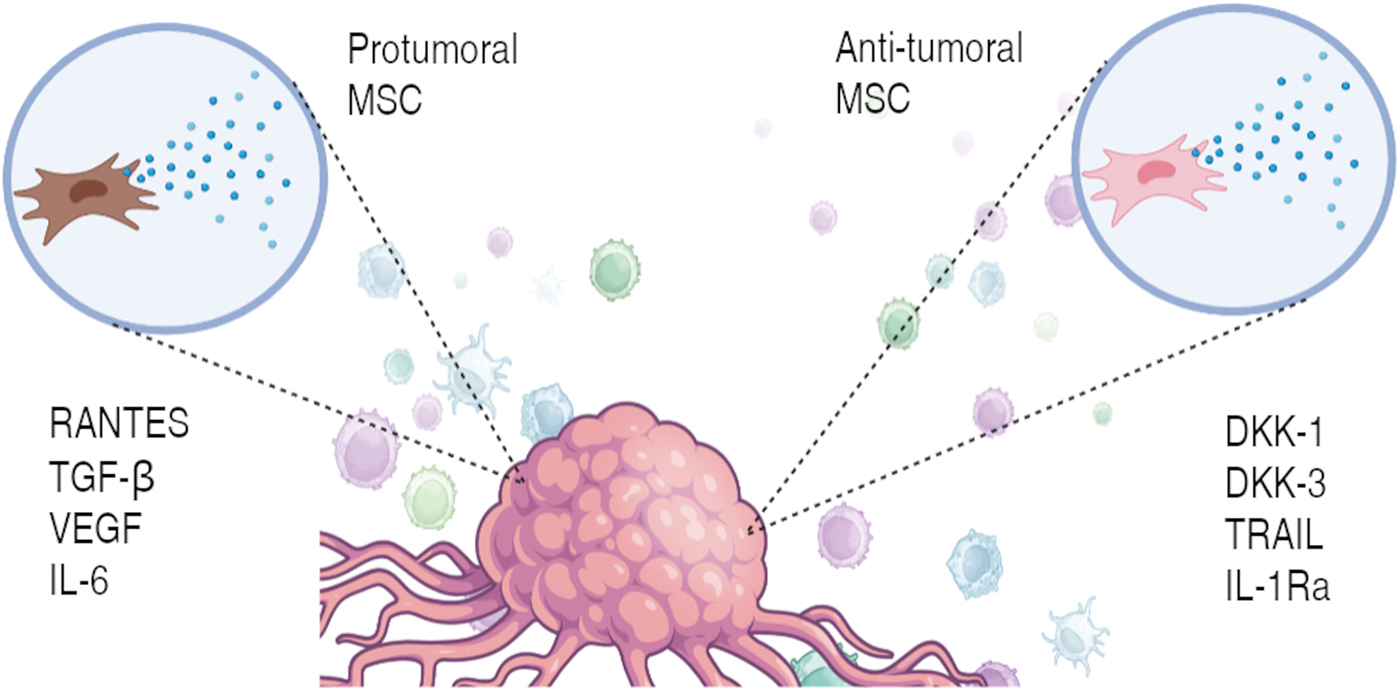
Possible MSC phenotypes in the tumor microenvironment. Figure created with BioRender.com Color images are available online.
A study carried out in mice showed that the injection of MSCs derived from the umbilical cord and adipose tissue can inhibit tumor growth in a breast cancer model, thanks to the migration and incorporation of the cells into the tumor. 79 Another study conducted in this same animal model, with MSCs derived from bone marrow in coculture with endothelial cells, demonstrated the in vitro incorporation of the MSCs in the capillaries formed by the endothelial cells, causing their destruction in proportion to the MSC concentration. In addition, it showed the existence of a direct interaction between them through the formation of communication channels through which MSCs incorporate reactive oxygen species into endothelial cells; whereas the in vivo evaluation showed a significant decrease in the tumor size, thanks to the decrease in angiogenesis. 80
However, adverse effects have been found when injecting MSCs derived from human adipose tissue in a model of prostate cancer in mice, where tumor volume was increased. 81 Another study was also carried out in a murine model of prostate cancer, but this time with MSCs derived from human bone marrow, also demonstrating a negative effect on tumor suppression. In this study, they found that the proliferative effect induced in tumor cells was mediated by soluble factors, specifically TGF-β. 82
As well as in regenerative medicine, research is being directed toward the use of MSC's secretome in the treatment of tumors. Using this approach, a study of the effect of conditioned medium obtained from perivascular MSCs from the Wharton's jelly on a glioblastoma model, showed that it promotes the proliferation of tumor cells in vitro and an increase the tumor size in vivo. Some proteins related to deregulated signaling pathways in cancer were identified, such as the Wnt related to tumor growth, migration, and invasion; the PDGF associated with proliferation, differentiation, and apoptosis; and the VEGF pathway, considered one of the most important in the regulation of angiogenesis. 12 Other works have shown to have a positive effect in the attenuation of tumor development, such as in a study carried out with MSC-CM derived from mouse adipose tissue, where it was shown to significantly inhibit tumor growth and induce apoptosis, last property highly related to the release of TRAIL and DKK-1 and DKK-3, inhibiting proliferation and differentiation by blocking the Wnt/β-Catenin pathway.83,84
Thus, there is a discrepancy regarding the role that MSCs may play in the TME.85–87 These events can occur, in the case of promoting tumor development, through different mechanisms such as the transition to CAFs and stimulation of the EMT, as well as the secretion of immunosuppressive and proangiogenic factors together with inhibitors of apoptosis, and in the case of the opposite effect through mechanisms that include the regulation of the cell cycle causing temporary arrest in the G1 phase of cancer cells, the regulation of signaling pathways associated with cell survival such as the PI3K/Akt pathway and the aforementioned Wnt/β-catenin pathway as well as the induction of apoptosis in endothelial cells by decreasing angiogenesis. 87
Secretome of MSCs in cancer
Although there are discrepancies about the effect that MSCs have on TME, which may be due to the use of MSCs of various origins, the used model or the culture conditions, 10 several studies have identified that MSCs that come into contact with TME acquire a protumoral phenotype. In a study on breast cancer conducted by Karnoub et al., 88 it was identified that the MSCs of the tumor stroma could increase the metastatic potential of different cell lines and also accelerate the growth of MCF7 cells in a murine model; they identified that this effect was mainly due to the MSC-secreted RANTES (CCL5), which was induced only with direct contact with tumor cells.
A similar result was obtained by Muehlberg et al. in a mouse model using 4T1 cells and MDA-MB-231 in combination with AMSC; the effect was reversed by knocking out CXCR4 expression in 4T1 cells. 89 A study carried out by Jazedje et al., after confirming the protumoral effect of coinjecting tumor cells and MSCs, was able to show that the intraperitoneal administration of the same MSCs in mice with tumors already established seven days before, decreased the metastasis rate, in addition, a second injection at 14 days significantly increased the survival of the mice. 90
MSCs are highly influenced by TME signals, whereas the secretome can be harvested from cells grown and expanded in vitro under controlled conditions. Under this approach, some studies have focused on microvesicles or exosomes, where an inhibitory effect on tumor growth has been identified in lines of breast cancer and melanoma, such as a study by He et al. on the effect of WJ-MSC-CM on MDA-MB-231, where it was found that WJ-MSC-CM inhibited the proliferation and migration of tumor cells by inhibiting EMT, in addition, the effect was potentiated in combination with radiotherapy; the authors suggest that the effect was due to the downregulation of Stat3. 91 Similar reports have identified the same inhibitory effect on these cells through TIMP1/2 secretion. 92 The AMSC-CM also decreased A375 viability, caused cell cycle arrest in G0/G1, and inhibited migration. 93 Similar effects have been found with the use of BM-MSC-CM 13 and fetal MSC-CM. 94
Engineering the MSC secretome
As discussed throughout the text, MSC heterogeneity can drive into different therapeutic outcomes; nonetheless, the effect of preconditioned MSCs has also been investigated with the aim of favoring the secretion of some cytokines or growth factors for different applications. 95 The use of LPS, for example, has been shown to induce a secretion profile with antitumor potential through TLR4 activation.15,96
Various parameters can affect the composition of the secretome.97–99 A three-dimensional (3D) culture, for example, provides an environment closer to the physiological one, and can trigger the expression of different phenotypes and increase or decrease the secretion of certain factors, as evidenced in the study carried out by Redondo-Castro et al., they found that MSC spheroids stimulated with IL-1, increased the number of secreted key factors for regeneration and immune response modulation when compared with two-dimensional (2D) cultures. 97
In the context of the TME, 3D cultures apart from having an effect on the secretome profile of MSCs and providing a physiological-like environment, 100 prevent communication between them and cancer cells. In a study carried out with WJ-MSC grown in drops of calcium alginate, it was possible to prove that 3D culture significantly improves the antitumor effect of these cells compared with 2D in a breast cancer model. In addition, it increased the viability and proliferation of the MSCs. In this study, an appreciable increase in the secretion of anti-inflammatory cytokines, VEGF, TGF-β, TNF-α, IFN-γ, IL-10, IL-6, and IL-3β, was noted in comparison to 2D. 101 Not only 3D but also hypoxic conditions, inflammatory stimuli (such as IFF-γ, TNF-α, LPS), the source and donor of MSCs present a differentiated secretion profile,102–104 which leaves a broad way to explore.
Effect of natural products on MSCs
Several studies have reported dose-dependent effects on the proliferation and differentiation of MSCs in vitro when they are treated with different natural products, primarily rich in polyphenols or flavonoids, 105 which are being studied as alternative treatments against different types of cancer, but little is known about the effect on the secretome. In the case of Naringin, rich in flavonoids and with antitumor properties, the induction of osteogenic differentiation was evidenced, accompanied by an increase in the proliferation of BM-MSC, similar to that observed with other flavonoids.105,106 Resveratrol, a polyphenolic phytoestrogen, promotes osteogenic differentiation and proliferation at low concentrations, but increasing concentrations also increase senescence, and cell cycle arrest.107,108 It has also been observed that natural products can alter the secretion of chemokines and cytokines in MSCs. Naringin itself can stimulate the secretion of chemokines with a C-X-C motif, favoring their migration. 109
In the context of the TME Di Pompo et al. showed that using nanoparticles loaded with curcumin, from the polyphenol family, can attenuate the protumoral phenotype of MSCs induced by acidosis, inhibiting the release of proinflammatory cytokines such as IL-6 and IL-8. 110 Prakoeswa et al. studied the effect of resveratrol on MSCs from different sources in the context of chronic wounds, also showing that at low concentrations, there is an increase in the proliferation of these cells accompanied by an increase in secretion of growth factors such as EGF, HGF, PDGF, and TGF-β1, being the AMSC the ones that showed the most significant response. 111 These results can be understood as beneficial for wounds; however, it could be the opposite in the context of cancer.
Conclusions
MSCs have always attracted attention as a heterogeneous group of cells with significant therapeutic potential, which has been partially exploited, as demonstrated by the multiple clinical trials carried out with the administration of these cells for different purposes. We are aware of its low immunogenicity, multipotency, and immunomodulatory capacity; however, their role within these tumors is not well known.
Many of the properties of MSCs can be attributable to their secretome; however, it has been identified that this can differ among MSC populations, which in turn will have different outcomes. Some culture methods and treatments have been studied to favor specific factor's secretion (or favor a particular phenotype), even in the context of cancer; however, there is a possibility that the effect is attenuated once the cells reach the TME. Among these, the use of natural products has been little explored in the context of cancer; it has been shown that several flavonoids and polyphenols can favor the proliferation and differentiation of MSCs, even altering the secretion of cytokines and chemokines; however, studies related to tumor development are needed. For now, it is necessary to better understand the possible transformation mechanisms of MSCs, either to use the cells themselves or if it is safer to obtain conditioned media with the desired characteristics under controlled conditions.
Footnotes
Acknowledgments
The authors would like to thank Ricardo Ballesteros from the GAT program (PUJ) for translating the article, and Prof. Alfonso Barreto (PUJ) for reading the article.
Authors' Contributions
JARI did the literature search and wrote the article; LMRM contributed to the literature search, reviewed, and edited the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
Funding was provided by the “Departamento Administrativo de Ciencia, Tecnología e Innovación COLCIENCIAS (792-2017 2a Convocatoria Ecosistema Científico “Generación de alternativas terapéuticas en cáncer a partir de plantas a través de la investigación y el desarrollo traslacional, articuladas en sistemas de valor ambiental y económicamente sostenibles”), Banco Mundial and Vicerrectoría de Investigaciones, Pontificia Universidad Javeriana, Bogotá, Colombia (contract no. FP44842-221-2018), MINCIENCIAS, Ministerio de Educación Nacional, Ministerio de Industria, Comercio y Turismo, and ICETEX.”