
Abstract
Craniofacial defects and dental tissue loss have significant negative impacts on the structure and function of jaws and face, often resulting in psychological issues in patients, emphasizing the urgent need for effective craniofacial tissue reconstruction. Unfortunately, natural regeneration of these tissues is limited. Dental-derived mesenchymal stem cells (MSCs) have emerged as a promising resource for tissue engineering-based therapeutic approaches. However, the clinical outcomes of MSC-based transplantation have not met expectations due to various complex reasons, and cellular senescence is recognized as one of the potential mechanisms contributing to the suboptimal results. The quality of MSC decreases during large-scale in vitro expansion, and it is also influenced by the age and the health status of donors. To address these challenges, extensive efforts have been made to developing strategies to combat senescence in tissue engineering, leveraging on current knowledge of underlying mechanisms. This review aims to elucidate the impact of cell senescence in craniofacial and dental regeneration and provides an overview of state-of-the-art antisenescence strategies. We first discuss the potential factors that trigger cell senescence in craniofacial tissue engineering. Then we describe senescence biomarkers, monitoring methods for senescent MSCs, and their underlying molecular mechanisms. The primary focus of this review is on current strategies to inhibit and alleviate cell senescence in tissue engineering. We summarize the strategies concerning the prevention of cell senescence, senolysis, modulation of the senescent associated secretory phenotype, and reversal of senescent MSCs, offering promising opportunities to overcome the challenges associated with cell senescence in craniofacial tissue engineering.
Impact statement
Cellular senescence presents a significant hurdle in harnessing the full potential of mesenchymal stem cells (MSCs) for therapeutic applications. This review focuses on exploring the impact of MSC senescence on craniofacial tissue regeneration. We include the latest advancements in senescence triggers, specific biomarkers, and detection methods. Furthermore, we summarize state-of-the-art antisenescence strategies, categorized as prevention interventions, senolysis, modulation of the senescent-associated secretory phenotype, and reversal of senescent MSCs. This review offers valuable insights into the role of cell senescence in craniofacial tissue regeneration and suggests potential directions for future research and therapeutic interventions.
Introduction
The craniofacial region is composed of variety of hard and soft tissues, including craniofacial differentiate bones, dental and periodontal tissues, cartilages, muscle, blood vessels, and nerves. The complex structure of these tissues and interactions among different cell types presents a significant challenge for craniofacial defect reconstruction. Mesenchymal stromal cell (MSC) based-regenerative medicine shows promise in addressing tissue and organ defects. MSCs possess multipotency and can differentiate into osteoblasts, chondrocytes, adipocytes, as well as specialized cells, such as myoblasts, neurons, and endothelial cells. 1
Dental tissue-derived MSCs have gained popularity as their typical MSC characteristics and presence in several oral tissues, including tooth connective tissue-derived dental pulp stem cells (DPSCs), stem cells from apical papilla (SCAPs), and stem cells from human exfoliated deciduous teeth (SHEDs); periodontal tissue-derived periodontal ligament stem cells (PDLSCs) and gingival mesenchymal stem cells (GMSCs); tooth germ-derived dental follicle stem cells (DFSCs); bone marrow mesenchymal stem cells (BMMCs) obtained from alveolar bone. These cells offer advantages such as ease of harvest, high yield, and ethical compliance, making them attractive for tissue regeneration, particularly in craniofacial tissue regeneration.2,3 However, the clinical outcomes of MSC-based transplantation have not met expectations. The negative impact of MSC senescence remains a major obstacle to achieve the safe and effective application of stem cells for tissue repair.
Cellular senescence is characterized by cell cycle arrest and morphological and functional changes, including reduced cell proliferation, increased cellular dysfunction, alternations in secretory features, and epigenetic modifications. 4 In 1968, Hayflick first proposed the phenomenon of cellular senescence, stating that most somatic cell types reach permanent cell cycle arrest after a specific number of population doublings. 5 MSCs typically undergo 30 to 40 population doublings in vitro before experiencing a progressive loss of stemness.6,7 MSCs derived from elderly or diseased donors may have decreased proliferative and differentiation capabilities compared to those from young or health donors. Uncontrolled cell senescence is a major cause of stem cell transplantation failure. Thus, combating MSC senescence is crucial to preserve their functional stability and maintain their effectiveness in tissue regeneration.8,9
In this review, we give a particular emphasis on the senescence of dental tissue-derived MSCs in craniofacial tissue engineering. We discuss the potential senescence triggers and summarize biomarkers with their detecting methods. Most importantly, we summarize the current progress in strategies to alleviate MSC senescence, offering promises for preventing and combating cell senescence in craniofacial tissue engineering.
Triggers of MSC Senescence
In vitro expansion
In vitro culture is essential for the expansion of MSCs as a large number of cells (10's to 100's millions) are required for a single clinical application. 10 However, MSCs do not proliferate indefinitely. Various conditions can induce cellular senescence as shown in Figure 1.
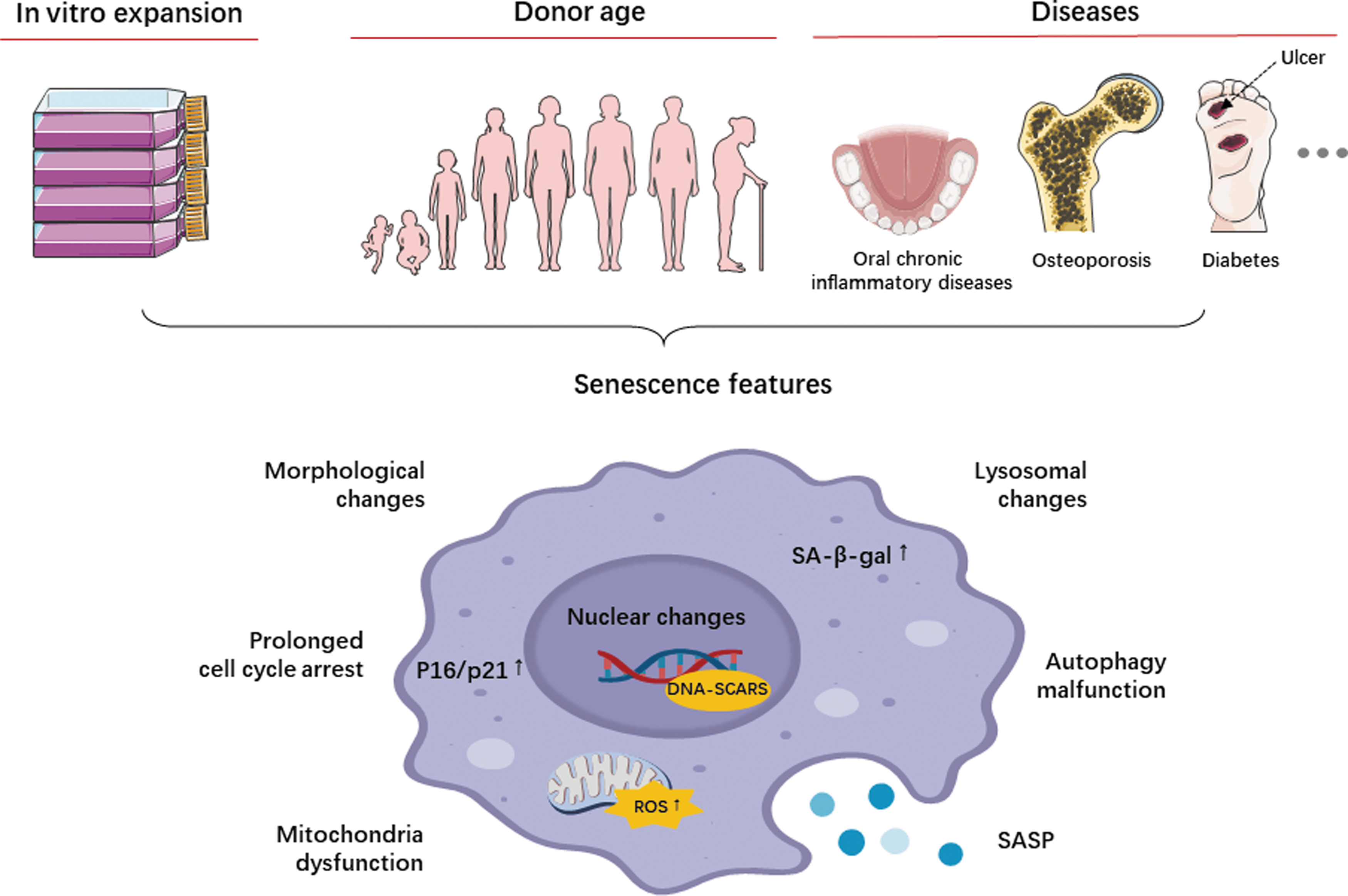
The potential inducers of dental derived-mesenchymal stromal cell senescence and the main features of cellular senescence.
Extensive expansion leads to replicative senescence.11,12 Prolonged in vitro culture of DPSCs was observed to cause telomere attrition and replicative senescence. 13 Interestingly, SHEDs exhibited a milder senescence phenotype even at late passages, displaying higher proliferation rates and osteogenic differentiation capabilities. 14 It may be attributed to the fact that SHEDs are obtained from younger donors, typically aged between 5 and 7 years. Nevertheless, it is recommended to limit MSC passages to 4–6 for therapeutic use.
Two-dimensional (2D) or three-dimensional (3D) culture model also affects stem cell properties. MSCs cultured in 3D systems display suppressed senescence signatures. 15 DPSC spheroids demonstrated superior multilineage differentiation capabilities compared to 2D monolayer cultures. Genes associated with stemness, osteogenesis, angiogenesis, and adipogenesis were significantly upregulated in 3D cultured DPSCs.16–18 Similar results were also found in other dental tissue-derived MSCs.19–21
Substrate stiffness serves as a biophysical cue that regulates stem cell properties through mechanotransduction.22,23 Soft culture substrates favor stem-like cellular phenotype and maintain the stemness of MSCs during long-term expansion. 24 Conversely, increasing the stiffness of the culture substrate enhanced the osteogenic/odontogenic differentiation of DPSCs, as well as promote bone defect repair and dentin formation. 25
In addition, excessive external stress contributes to premature senescence of stem cells. Stressors primarily include altered levels of growth factors or nutrients and increased oxygen levels. The latter leads to oxidative stress, an imbalanced reactive oxygen species (ROS) production, and antioxidant protection, resulting in senescence accumulation in cells. 26
Donor age
Donor age is another important factor influencing the senescence of MSCs.27,28 DPSCs from permanent teeth showed decreased stemness marker OCT3/4 and CXCR4 expression, prolonged cell doubling time, and higher apoptotic rates. 29 Lezzi et al. investigated the influence of donor age on DPSC quality during in vitro expansion, comparing cells obtained from juvenile, middle age, and aged donors. The results indicated a decline with age in osteogenic/odontogenic and neurogenic differentiation potential, although no significant morphological changes were observed. 30 Alzer et al. further demonstrated that DPSCs from aged teeth complete or partial loss the ability to differentiate into both neuronal and osteogenic lineages, but exhibited a preference for adipogenic differentiation. 31 DPSCs displayed enhanced proliferation and mineralization ability in response to mild inflammatory stimulation, but this effect decreased with increasing donor age. 32
In addition, several studies have reported a decline in PDLSCs' proliferation, migration, osteogenic differentiation ability, expression of pluripotency-associated transcription factors, and immunosuppressive ability with increasing donor age.33,34 Aged PDLSCs also exhibited impaired ability to form functional cell sheets. 35
Interestingly, a recent study found that the properties of GMSCs were not evenly affected by donor age. Although aged GMSCs showed decreased adipogenesis and osteogenesis, their differentiation potential toward neuronal lineage and immunosuppressive behavior in lipopolysaccharide (LPS)-induced acute lung injury was not affected by donor age. 36 Researchers explained that aged gingival tissues underwent apoptosis or autophagy to maintain tissue homeostasis, but more detailed studies are still required to further investigation.
In conclusion, donor age negatively impacts on the quantity and quality of MSCs, which could limit the availability of sufficient MSCs for autologous transplantation, especially among the elderly patients. Thus, reliable strategies are needed to monitor the MSC quality in the different treatment regimens.
Diseases
The quality of MSCs can be influenced by various diseases, including local chronic inflammatory diseases and age-related diseases. These diseases can lead to dysfunction in MSCs and even premature senescence.37,38
Periodontitis is one of the most prevalent inflammatory diseases in oral health. The microenvironment in periodontitis contributes to the accumulation of senescent cells. The release of ROS in periodontitis serves to defend against invading bacteria but can also cause damage to healthy host cells, resulting in long-term DNA damage and leading to cellular senescence. The accumulation of senescent alveolar bone osteocytes exacerbates chronic inflammation and generates aging-related secretory phenotypes, thereby deteriorating the periodontal microenvironment. 39
Osteoporosis is a systemic chronic inflammatory disease that affects the craniofacial region. The pro-inflammatory bone marrow microenvironment, with increased levels of cytokines such as IL-6 and IL-1, influences bone metabolism and age-related bone loss. In particular, bacteria-activated T lymphocytes stimulating RANKL signaling play a role in promoting osteoclastogenesis and local alveolar bone loss. 40
Diabetes is closely associated with periodontal disease. Hyperglycemia in diabetes increases oxidative stress, which accelerates DNA damage and cell senescence.41,42 Tang et al. demonstrated that diabetes-induced oxidative stress contributed to telomere dysfunction and senescence of PDLSCs, further exacerbating periodontal bone loss in periodontitis. 43 In addition, the formation of advanced glycation end products in diabetes inhibited the osteogenic differentiation of PDLSCs and induced apoptosis of PDLSCs through the JNK-mitochondrial pathway.44,45
Understanding the effects of diseases on MSCs is crucial for personalized treatment approaches for specific disease conditions. Thorough assessments of MSC activities in disease states can guide the selection and optimization of MSC-based therapies to improve clinical outcomes.
Detection of Senescence Markers
The identification of senescence cannot rely on a single feature; instead, it requires the presence of multiple features in combination. In this study, we provide an overview of the primary senescence markers, along with their corresponding detection methods and underlying mechanisms, as summarized in Table 1.
List of Biomarkers Commonly Used for the Assessment of Cellular Senescence
IF, immunofluorescence; FC, flow cytometry; IHC, immunohistochemistry; WB, western blotting; ELISA, enzyme-linked immunosorbent assay; Q-FISH, quantitative fluorescence in situ hybridization; ROS, reactive oxygen species; SAHF, senescence-associated heterochromatin foci; SASP, senescent associated secretory phenotype; SA-β-gal, senescence-associated beta-galactosidase; TRF, telomere restriction fragment analysis; TEM, transmission electron microscopy.
Morphologic changes
Senescent cells exhibit enlarged and vacuolized appearance, condensed nuclei, cytoplasmic granularity, and the presence of intracellular debris. Recent studies have highlighted the connection between cell size and function, with senescent cells often displaying increased size due to dysregulated cell growth and division. As cells grow beyond a certain size, protein synthesis and DNA content fail to scale in proportion to cell volume, leading to cytoplasm dilution and limiting DNA availability for cell function. 46 Moreover, the enlarged morphology is associated with elevated stress fiber formation mediated by upregulated cofilin1. 47
Cell morphology and cytoplasmic granularity can be easily assessed using light microscopy and flow cytometry-based analysis.12,48 Confocal imaging following DAPI staining is typically used to detect multinucleation. 49 However, these measurements are qualitative and challenging to quantify or detect in vivo.
Cell cycle arrest
Two types of cyclin-dependent kinase (CDK) inhibitors, p21CIP (hereafter p21) and p16INK4a (hereafter p16), are the main drivers of cell cycle arrest in senescence. P21 has a dual role in cell cycle progression, acting as both an inhibitor and a promoter. It can induce cell cycle arrest in response to stress-induced p53 activation, while transient DNA damage-induced expression of p21 facilitates DNA repair and cell survival. 50 P21 is important for initiation of senescence in some settings, but its expression does not persist in senescent cells. Therefore, p21 alone cannot be a definitive marker of the senescence phenotype. In contrast, p16-RB pathway plays a more significant role in maintaining the senescent state. 51 P16 directly interacts and inhibits CDK4/6, leading to cell cycle arrest through the accumulation of phosphorylated pRb and inhibition of E2F transcription factors. 52 While measuring p16 protein levels can be challenging and hindered in mice due to the lack of specific antibodies, the use of p16 reporter mouse models helps overcome this limitation.53,54 Therefore, P16 is widely used as a decisive marker for senescent cells.
Cell cycle arrest can also be assessed using proliferation or DNA replication assays, such as Ki-67, EdU, or BrdU staining. However, lack of proliferation is not a specific marker of senescent cells, so it is impractical for measuring senescence in postmitotic cells and inadequate for in vivo experiments.
Nuclear changes
Telomere shortening is a well-established biomarker of cellular senescence. 55 Telomeres are specialized nuclear protein caps at the end of chromosomes that play a crucial role in maintaining genome stability during DNA replication. With each cell division, telomeres progressively shorten until they reach a critical length, triggering cellular senescence.56,57 Telomerase, an enzyme that restores the missing 5′-TTAGGG DNA repeats, can prevent telomere shortening. However, most human somatic cells, including MSCs, do not express telomerase, leading to progressive telomere erosion and replicative senescence. 58 Telomere length can be measured by qPCR and quantitative fluorescence in situ hybridization. 59
Senescent cells often exhibit senescence-associated heterochromatin foci (SAHFs), which are condensed DNA regions associated with transcriptional silencing of proliferation-related genes. 60 Human senescent cells in culture display punctate SAHFs stained by DAPI and marked by H3K9me3 accumulation and incorporation of heterochromatin protein 1.49,61 SAHFs are commonly used to identify oncogene-induced senescence in human cells.
Senescent cells also accumulate persistent nuclear DNA damage lesions known as DNA segments with chromatin alterations reinforcing senescence (DNA-SCARS). DNA-SCARS associate with promyelocytic leukemia nuclear bodies and accumulate DNA damage response (DDR) proteins such as ATM and ATR and CHK2. Phosphorylated H2A histone family member X (γ-H2AX) foci can serve as a marker for DNA- SCARS and can be detected by immunoblotting or imaging techniques. 62 However, not all senescence-inducing stimuli lead to persistent DDR.
Loss of lamin B1 from the nuclear envelope is another common marker of senescent cells. Lamin B1 is a structural protein of the nuclear lamina and involved in maintaining nuclear integrity. 63 Its destabilization results in the formation and release of cytoplasmic chromatin fragments (CCFs). CCFs are positive for γ-H2AX and enriched for the histone H3K27me3. 64 Loss of lamin B1 can be detected through immunoassays and visualized using imaging or immunoblotting techniques.
Lysosomal changes
The senescence state is characterized by increased lysosomal activities and upregulation of lysosomal proteins. Senescence-associated beta-galactosidase (SA-β-gal) is a widely used marker to assess cellular senescence. SA-β-gal is a lysosomal enzyme whose activity can be detected using cytochemistry, histochemistry, or fluorescence-based methods. The assay measures the increased expression and activity of SA-β-gal, indicating elevated lysosomal content in senescent cells. 65 However, the SA-β-gal assay may have limitations, yielding false-positive or -negative results in certain cases. It is not suitable for paraffin-embedded tissue sections and has limitations when applied to live cells.
Lipofuscin accumulation is another notable lysosomal change observed in senescent cells. Lipofuscin is a membrane-bound waste product that cannot be degraded or expelled from the cell but can only be diluted through cell division and growth. It is associated with protein and lipid damage, as well as mitochondrial dysfunction. 66 Sudan-Black-B histochemical stain is commonly used to specifically detect lipofuscin accumulation. 67
Mitochondrial dysfunction
Mitochondrial dysfunction plays a crucial role in the maintenance of the pro-inflammatory phenotype observed in senescent cells, despite not being commonly utilized for senescent cell detection. Senescent cells display distinct alterations in mitochondrial characteristics, including mass, membrane potential, and morphology. 68 These structural changes in mitochondria lead to the release of mitochondrial enzymes and enhanced production of ROS. Several fluorescent dyes, such as Dihydroethidium, Dihydrorhodamine 123 (DHR123), and MitoSOX™ Red, can be used to detect mitochondrial-derived ROS or overall intracellular ROS levels.
Senescent associated secretory phenotype
Senescent cells secrete a diverse array of factors, including pro-inflammatory cytokines, chemokines, growth factors, angiogenic factors, matrix metalloproteinases (MMPs), and extracellular vesicles (EVs), collectively referred to as the senescent associated secretory phenotype (SASP). 69 The SASP exhibits high heterogeneity, with some SASP factors involved in tissue remodeling, embryonic structure reorganization, and immunosurveillance, while others, such as interleukins and MMPs, have been associated with aging and disease phenotypes. Certain SASP components are packaged in small EVs that can exert distal effects, such as promoting cancer cell proliferation.70,71
Various techniques, including RT-qPCR, proteomic techniques (e.g., LC MS/MS), cytokine arrays, and ELISA, can be used to identify most SASP components.72,73 Evaluating the composition of the SASP can help distinguish different senescence programs. For instance, senescent cells associated with tissue repair exhibit increased expression of MMPs and growth factors like PDGF-A and VEGF, while age-related senescent cells are predominantly characterized by pro-inflammatory factors. 74
Autophagy malfunction
Autophagy has emerged as a new hallmark of aging and stem cell senescence.75,76 It is a process that facilitates degradation of intracellular components through lysosomal activity, known as autophagic flux. The main form of autophagy is macroautophagy, which is closely associated with cell aging. Autophagic activity tends to decline in multiple tissues during aging. Possible mechanisms of how autophagy delays aging involve the removal of protein aggregates or damaged mitochondria, prevention of stem cell attrition and oncogenic transformation, the reduction of inflammatory responses, and nuclear autophagy. 77
Monitoring autophagic flux commonly involves the detection of LC3 protein processing. LC3 proteins undergo specific cleavage by Atg4 at the C terminus, generating LC3-I, which then conjugates to phosphatidylethanolamine to form LC3-II. Since LC3-II is degraded in autolysosomes, its levels are widely used as a marker to monitor autophagy. In addition, the expression of autophagy-related genes (Atg) can indicate the autophagic flux. Studies on cell senescence have demonstrated a decrease in autophagy activity, as indicated by reduced LC3 protein expression and decreased expression of autophagy genes, such as Atg1, Atg5, Atg6, Atg7, Atg8, and Atg12, in several animal models and human tissues.75,78,79
Novel potential markers and detection methods
Senescent cells exhibit heterogeneity in different tissues, indicating the importance of exploring cellular senescence markers beyond the commonly studied, such as p19, uPAR, and GPNMB. 80 This highlights the need for a deeper understanding of senescent cell heterogeneity. The advent of single cell RNA-sequencing (scRNA-seq) has revolutionized the study of senescence heterogeneity by enabling the characterization of transcriptomic profiles at the single-cell level. Several studies have successfully utilized single-cell technologies to identify senescence-associated signatures in the context of aging.81,82 More recently, spatial transcriptomic technology has been introduced in the field of senescence, enabling the mapping gene expression patterns within intact tissue and providing spatial context to transcriptomic data. 83 However, it is important to note that current studies represent only the initial progress, and we anticipate that continued advancements will likely revolutionize our understanding of senescence.
The application of Artificial Intelligence (AI) in image translation has demonstrated great potential in effectively identifying senescent cells within a large cell population. This technology enables the identification of visual features such as cell morphology features84,85 and nuclear morphology. 86 These AI techniques can be integrated with cell culture routines, making them easily accessible tools for MSC manufacturing in the future.
Current Strategies to Combat Cell Senescence in Craniofacial Tissue Engineering
Prevention of cellular senescence
Inhibition of ROS
Modulating redox balance through the addition of antioxidants or adjusting oxygen tension in culture holds promise for maintaining stemness and reducing senescence as summarized in Figure 2.
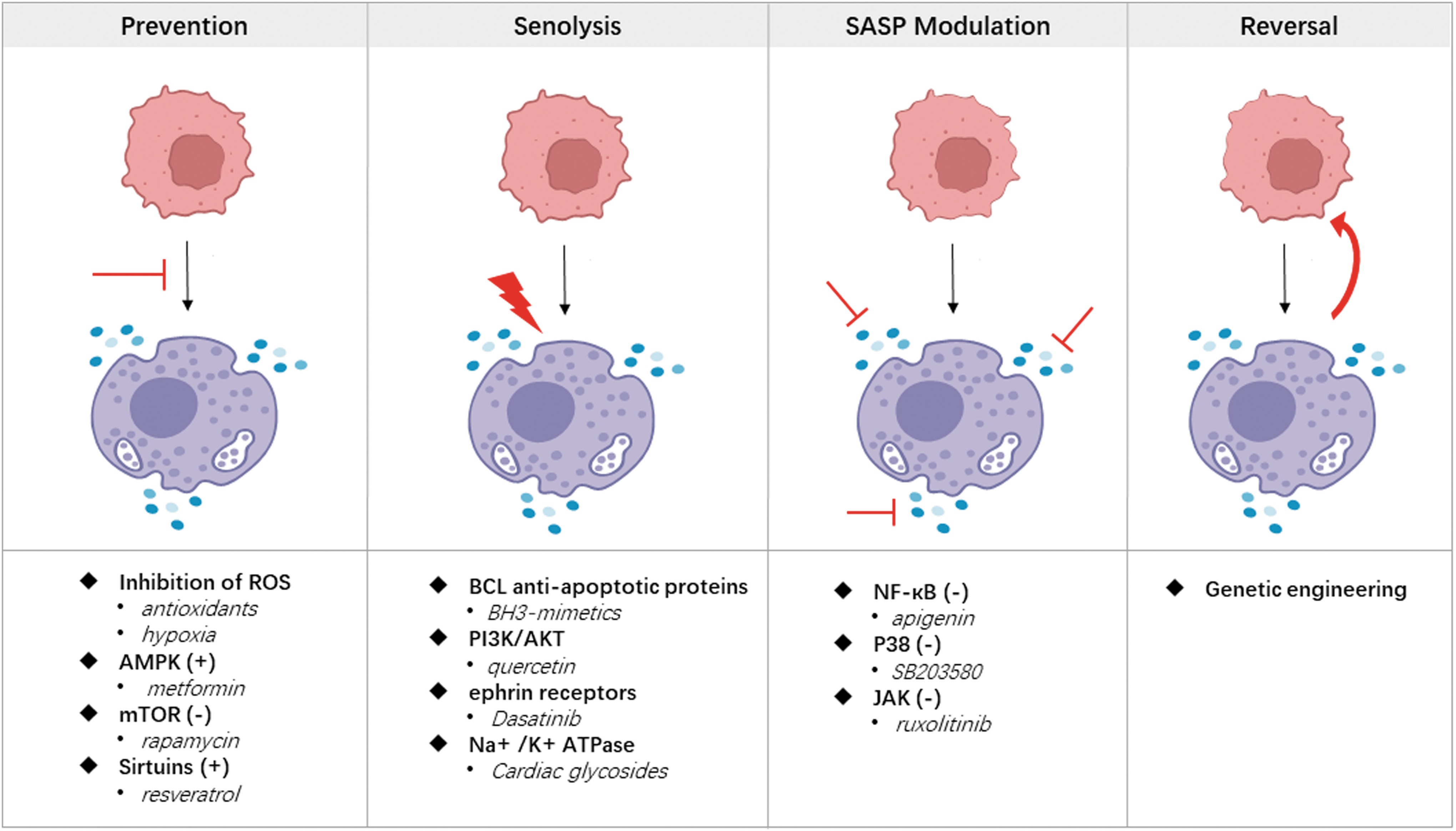
The four main strategies to combat cell senescence.
Antioxidants, including N-acetyl-l-cysteine (NAC), ascorbic acid, melatonin, or tannic acid (TA), have been found to inhibit ROS production or scavenge ROS directly in dental-derived MSCs. For example, ascorbic acid enhanced the pluripotency of PDLSCs by reducing the expression of Notch3, thus inhibiting cell aging. 87 Melatonin attenuated long-term expansion-induced PDLSC senescence and restored autophagy through PI3K/AKT/mTOR signaling. 88 TA coated PDLSC spheroids promoted cell viability and stemness related signaling pathways such as MAPK, Hippo, and Wnt. 89 Meng et al. identified 803 differentially expressed genes by RNA-sequencing between DFSCs with and without 5 mM NAC. They found that PI3K/AKT/ROS signaling was significantly enriched in NAC treated DFSCs. 90 However, the optimal dosage of antioxidants remains unclear, and caution should be paid to avoid DNA damage resulting from excessive antioxidant use.
Long-term normoxic culture of hDPSCs leads to loss of stemness and senescence, while hypoxic culture can alleviate these effects. Hypoxic conditions increase the expression of stemness markers STRO-1 and OCT4, reduce the number of senescent cells, and downregulate senescence-related genes P53 and TGF-β. 91 Noncoding RNA expression, such as the upregulation of miR-140-3p, was involved in hypoxia-mediated effects on DPSCs, promoting their differentiation into bone/dentin by targeting KMT5B. 92 However, in the case of PDLSC, hypoxia-inducing factors and TGF-β1 synergically inhibited osteogenic differentiation. 93 Further investigations are required to determine the optimal hypoxia conditions on each specific type of MSC behavior.
Pharmacological interventions
Pharmacological targeting of AMPK/mTOR signaling is a prominent therapeutic strategy to prevent senescence. Signaling pathways involved in cellular energy status and nutrient sensing directly influence senescence by regulating cell growth arrest, prolonged cell proliferation, or sustained self-renewal of stem cells in culture. AMPK and mTOR are key enzymes in this intricate signaling network. Manipulating AMPK or mTOR has been shown to inhibit senescence in MSCs. For instance, metformin can prevent MSC senescence by activating AMPK, sirtuins, autophagy, and mitochondrial biogenesis. Metformin inhibits DPSC senescence and exerts antiaging effects through the AMPK/mTOR signaling pathway by downregulating miR-34a-3p. 94 Inactivation of mTOR by rapamycin is also promising for antiaging therapy, as it can prevent cellular senescence and age-related disease in animal models. Rapamycin was found to modulate the age-associated oral microbiome into a “younger” composition and reduced periodontal inflammation. 95
Pharmacological activation of Sirtuins also prevent cellular senescence. Sirtuins, a family of NAD-dependent histone deacetylases, are closely involved in glucose metabolism, playing a crucial role in halting cellular senescence and age-related diseases. 96 Several sirtuin members, including SIRT1, SIRT3, SIRT6, and SIRT7, have been identified as potential antiaging factors. Activation of SIRT1 protects senescent cells from p53-dependent senescence and induces autophagy to prevent cellular senescence or apoptosis. 97 SIRT3 expression decreases during in vitro expansion of MSCs, and its depletion accelerates aging and inhibits MSC differentiation into osteoblasts and adipocytes. 98 Similarly, Sirt6 overexpression prevents replicative senescence in human cells. 99
Sirtuin-activating compounds, such as resveratrol, fisetin, or NAD+ precursors, can reduce cellular senescence by activating AMPK. 100 Resveratrol, for example, exhibits antioxidant and anti-inflammatory properties, enhances tissue resistance to oxidative stress, reduces pro-inflammatory factors, and increases superoxide dismutase levels.101,102 Resveratrol enhanced bone differentiation in DPSC through promotion of SIRT1 expression and activation of Wnt/β-catenin signaling pathway. 103 SIRT1 also regulated SCAP proliferation and odontogenic differentiation by increased p53 deacetylation. 104 These findings highlight the importance of the SIRT family as targets for regulating cellular senescence in dental-derived MSCs.
Growth factors
Growth factors, such as FGF and IGF, have been extensively investigated due to their potential in promoting cell proliferation, maintaining stemness, and inhibiting cellular senescence. FGF is mostly studied for its role in maintaining stem cell stemness.105,106 The treatment with basic FGF (bFGF) enhanced the proliferation and clonal potential of SHEDs and adult DPSCs, while not affecting chondrogenic differentiation. In addition, IL-17 has been found to increase the proliferation potential of SHEDs and stimulate early osteogenesis in DPSCs and SHEDs, contributing to the preservation of stemness and prevention of cellular senescence. 107
The insulin-like growth factor binding protein (IGFBP) family, a subset of the IGF family, plays a crucial role in growth, development, senescence, and apoptosis in various tissue cells. 108 Among them, IGFBP7 has been identified as a regulator of apoptosis, proliferation, and bone metabolism. 109 IGFBP7 prevented senescence in DPSC through metabolically induced deacetylation of H3K36ac and decreased transcription of p21. The effects of IGFBP7 on senescence may vary with tissue source and concentration. 110 Moreover, IGF-1 has been recognized as a multifunctional peptide that promoted osteogenic differentiation of BMMSCs.111,112 Similarly, Feng et al. investigated the effects of IGF-1 on DPSCs and observed that it promoted proliferation and osteogenic differentiation through the mTOR pathway. 112 Therefore, the application of growth factor FGF or IGF can enhance regenerative potential and provide a strategy for preventing cell senescence.
Senolysis
The targeted elimination of senescent cells using senolytics is a promising approach for addressing age-related diseases. Senolytics specifically aim to induce death in senescent cells without affecting proliferating or quiescent cells by targeting vulnerabilities existing unique to senescent cells. These vulnerabilities mainly include the increased expression of BCL antiapoptotic proteins (navitoclax and other BH3-mimetics), activation of the PI3K/AKT pathway (quercetin), ephrin receptors (dasatinib), lysosomal V-ATPase (bafilomycin A1, concanamycinA), Na +/K + ATPase (cardiac glycosides), and dysregulation of the mTOR pathway (sertraline). 113 In addition, general cytotoxic drugs like the antibiotic duocarmycin can be transformed into senolytics by incorporating a galactose motif, enabling selective cleavage under high β-galactosidase expression and targeted delivery to senescent cells. 114 Some antibiotics, including azithromycin and roxithromycin, exhibit preferential cytotoxicity against senescent fibroblasts, although the underlying mechanism remains unclear. 115
Nevertheless, the efficacy of senolysis treatment is limited, as different types of senescence display varying sensitivities to senolytics. Further research is needed to fully understand the potential of senolytics in combating senescent cells.
SASP modulation
Suppressing the SASP offers an alternative therapeutic strategy to alleviate cellular senescence-related phenotypes. SASP inhibitors, also known as senomorphics, target various signaling pathways, including NF-κB, mTOR, IL-1α, and p38 MAPK, to suppress SASP expression. In dental-derived MSCs, NF-κB inhibitors such as apigenin or glucocorticoids have been shown to reduce pro-inflammatory SASP phenotype.116–118 Activation of p38 MAPK by various senescence-inducing stimuli leads to irreversible cell cycle arrest, senescence, and SASP development. Inhibition of p38MAPK using SB203580 markedly reduces the secretion of SASP factors. 119 JAK inhibitor ruxolitinib reduces senescence and attenuates SASP cytokines and chemokines in senescent cells. 120 Furthermore, the mTOR inhibitor rapamycin was showed to modulate the composition of SASP toward a less damaging secretome and increased life span in mice. 121
Senomorphic therapy aims to alleviate the detrimental effects of a persistent SASP without eliminating senescent cells. However, it is important to note that some SASP inhibitors may have off-target effects. For example, rapamycin at higher doses in mice can cause nephrotoxicity, metabolic impairment, and susceptibility to infections. 122 Careful consideration of agent-specific effects and dosages is crucial for the safe and effective use of SASP inhibitors.
Reversal of senescent MSCs
Genetic engineering is commonly used to reverse the senescent state of MSCs. Senescence-related genes are of great interest in senescence study and the development of antisenescence strategies. Genetic engineering techniques have proven effective in reversing MSC senescence. For example, activation of Yes-associated protein (YAP) in the Hippo-YAP signaling pathway promotes proliferation, inhibits cell apoptosis, and delays senescence in PDLSCs. 123 Deletion of FAM96B leads to decreased telomerase reverse transcriptase activity and increased levels of senescence-related markers and proteins, while overexpression of FAM96B enhances the proliferation potential of DPSC and inhibits senescence. 124 Adrenomedullin (ADM), which is highly expressed in dental tissues during critical tooth development stages, promotes the proliferation of DPSCs through the JNK/C-Jun pathway and inhibits apoptosis through the Src/SK-3b pathway. 125 Dou et al. further demonstrated that ADM promoted CCNA2 expression by inhibiting miR-152, thereby enhancing cell cycle progression and resisting cell senescence. 126
Notably, knockdown or overexpression of specific genes can significantly restore the proliferation and differentiation potential of MSCs. However, it is crucial to consider the risk of cancer development associated with dysfunctional genes that may promote tumorigenesis. Careful evaluation and monitoring are necessary when employing genetic engineering techniques for reversing senescence in MSCs.
Clinical trials
At present, clinical trials to inhibit or alleviate cell senescence have shown promising results. Senolytic drugs, such as Dasatinib and Quercetin, have demonstrated improved physical function in patients with idiopathic pulmonary fibrosis 127 and reduced senescent cell in diabetic kidney disease patients. 128 Multiple other clinical trials, including Alzheimer's disease, osteoporosis, chronic kidney disease, and osteoarthritis, are underway. 129 However, a clinical trial using senolytic compound UBX0101 for treatment of osteoarthritis (NCT03513016) was recently halted in phase 2, as it did not achieve its primary pain improvement goal. 129 Considering unknown side effects in humans, current clinical trials are limited to patients with serious health conditions. Further research is necessary to fully comprehend the potential implications of these treatments in effectively combating cellular senescence.
Conclusion
Studies of cellular senescence have experienced exponential growth in the past decade due to its widespread implications. However, there are still substantial gaps and key questions that need to be addressed in this field.
First, we anticipate significant advancements in technology that will enable multilevel analyses of senescent cells at tissue-specific, cellular, subcellular, and single-molecule resolution. These analyses will encompass the proteome, transcriptome, epigenome, and metabolome, providing a more comprehensive understanding of cellular senescence.
Second, the development of novel senolytic drugs and SASP modulators specific to dental tissue-derived MSCs could offer more precise and efficient strategies to target senescent cells and mitigate their detrimental effects. Preclinical studies should be conducted to assess the efficacy and potential side effects of these agents before advancing to human clinical trials.
Third, clinical trials focusing on MSC-based therapies should integrate strategies to combat cellular senescence to enhance treatment outcomes. Long-term follow-up of patients receiving MSC-based regenerative treatments will provide valuable insights into the efficacy and safety of these approaches in vivo.
Continued research in this field will pave the way for innovative and effective strategies to combat cellular senescence, ultimately improving patient outcomes and the success of MSC-based therapies in tissue regeneration.
Authors' Contributions
W.T. wrote the article. F.H. contributed to material collections and figure design. J.L. provided essential conceptual insights and supervised the project. S.Z. and W.T. critically reviewed and edited the article. All authors approved the final version of the article.
Footnotes
Disclosure Statement
The authors declared no potential conflicts of interest.
Funding Information
This study was supported by the National Natural Science Foundation of China (82201029), Sichuan Science and Technology Program (2022YFS0283), Research and Develop Program, West China Hospital of Stomatology Sichuan University (RD-02-202113).