
Abstract
Extensive bone fractures, which can seriously impact both health and quality of life, cannot easily heal naturally, especially if the patient has an underlying medical condition or is aging. The most promising approach to addressing such fractures is bone regeneration through bone tissue engineering. Bone regeneration is a complex process that consists of three distinct phases: inflammation, repair, and remodeling. Macrophages play a bridging role between the various cells involved in each stage of bone regeneration, interacting with different microenvironments and advancing the bone healing process. Although the origin and function of macrophages have been extensively studied, the mechanisms underlying their interaction with the bone healing microenvironment remain unexplored, including the association of microenvironmental changes with macrophage reprogramming and the role of macrophages in cells in the microenvironment. This review summarizes the bone regeneration process and recent advances in research on interactions between macrophages and the bone healing microenvironment and discusses novel biological strategies to promote bone regeneration by modulating macrophages for the treatment of bone injury and loss.
Impact statement
In this review, we focus on the relationship between the bone healing microenvironment and cells during the three stages of bone healing. We focused on the metabolic reprogramming of macrophages by the microenvironment, which in turn leads to alterations in their phenotype, as well as macrophages influencing components of the microenvironment through paracrine secretion. In addition, we present recent advances in the improvement of bone regeneration materials through macrophage modulation, which will be useful for subsequent research and clinical translation.
Introduction
Millions of people worldwide suffer from bone injury and loss each year, with external injuries and changes in bone density being important causes. Studies have shown that over two million patients in the United States experience fractures due to osteoporosis each year. 1 Furthermore, ∼3.5 billion people worldwide suffer from periodontitis, and most patients with intermediate to advanced disease experience severe bone loss, which leads to tooth loss and seriously impacts quality of life. 2 Studies have shown that small fractures heal spontaneously, whereas segmental bone defects, including extensive bone defects caused by cancer and alveolar bone defects, do not heal naturally once they exceed 2–2.5 times their diameter or lack adequate soft tissue coverage. 3
Bone healing is a lengthy process, with bone often requiring 3–6 months to reach sufficient strength, and months or even years to remodel back to its original level. 4 Aging patients and those with conditions such as diabetes and osteoporosis are more susceptible to poor bone healing, 5 such as nonunion, incomplete healing, or delayed healing, which can adversely affect their physical and mental health and quality of life. Therefore, repairing lost bone tissue and restoring its function are major clinical challenges. The ideal solution to these problems is natural regeneration of bone through bone tissue engineering.
The first step involved in restoring bone to its original state through bone tissue engineering is to gain a thorough understanding of the bone healing process. Bone regeneration involves three phases: inflammation, repair, and remodeling. The inflammatory microenvironment contains high levels of proinflammatory factors that initiate an inflammatory response by recruiting immune cells to remove pathogens and apoptotic cells (ACs). As the levels of anti-inflammatory factors gradually increase and inflammation subsides, the inflammatory microenvironment transforms into a repair and remodeling microenvironment.6,7 This transformation initiates the dynamic process of bone matrix production and resorption, and allows some immune cells to undergo metabolic reprogramming, producing different phenotypes to regulate the balance of osteoclasts and osteoblasts. 8
In addition to the effects of the microenvironment on cells, cells such as macrophages also influence the microenvironment. 9 As immune cells, macrophages in the inflammatory phase secrete various proinflammatory factors into the microenvironment, which further recruit inflammatory cells. An excess of proinflammatory macrophages can prolong the inflammatory microenvironment, which adversely affects bone healing. 10 When the macrophage phenotype is altered, the secretion of anti-inflammatory factors and cytokines facilitates the transformation of the inflammatory microenvironment into a reparative microenvironment.11,12
Considering the different roles of three phases of the microenvironment in bone healing, new strategies for modulating the immune response may be useful for the treatment of bone injuries that cannot heal naturally. Some strategies operate during the inflammatory phase to ensure that the inflammatory response is appropriately timed, which helps prevent tissue damage due to excessive inflammation, while ensuring a sufficient duration of inflammation to fully clear pathogens. 13 Other strategies focus on the repair and remodeling phases to promote the phenotypic transformation of immune cells and ensure a balance between osteoclasts and osteoblasts to achieve the desired bone healing effect.14,15
In this review, we summarize the process of bone healing and the role of immune cells, osteoblasts, and osteoclasts in this process and highlight recent advances in research on the interaction between macrophages and the bone-healing microenvironment. Finally, we discuss bone regeneration materials that modulate inflammation, repair, and remodeling phases, or perform temporal regulation by modulating macrophages, providing a reference for future clinical translation (Fig. 1).
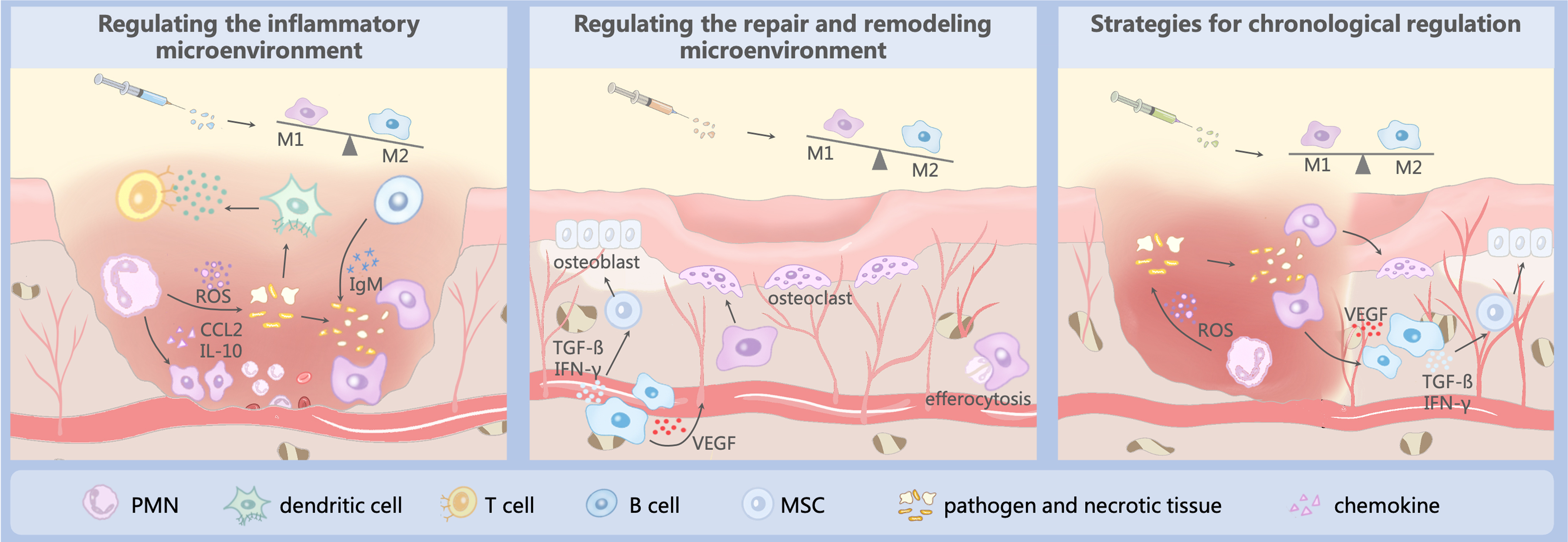
Bone healing microenvironment and bone regeneration strategies: (1) Promote the secretion of anti-inflammatory factors by M2 macrophages to reduce inflammation, (2) promote M2 polarization to ensure a balance of bone formation and resorption, and (3) chronological regulation: Step 1: suitable inflammation to remove pathogens and bone fragments; Step 2: promote M2 polarization to reduce inflammation and promote bone regeneration. Color images are available online.
Bone Healing
One healing is the process of bone reconstruction after a fracture through a series of cellular and molecular events that enable the regeneration of bone tissues and the restoration of their original tissue structure. The complex regeneration process comprises two stages: primary (direct) and secondary (indirect) fracture healing.10,16 In primary fracture healing, the bone heals directly through the reconstruction of unit bone fragments under the conditions of precise anatomical restoration and stable fixation. In most cases, fractures heal through secondary fracture healing in three stages: inflammation, repair, and remodeling (Fig. 2). 17 As secondary fracture healing is associated with bone tissue engineering, only the cellular mechanisms involved in secondary fracture healing are reviewed in this section.
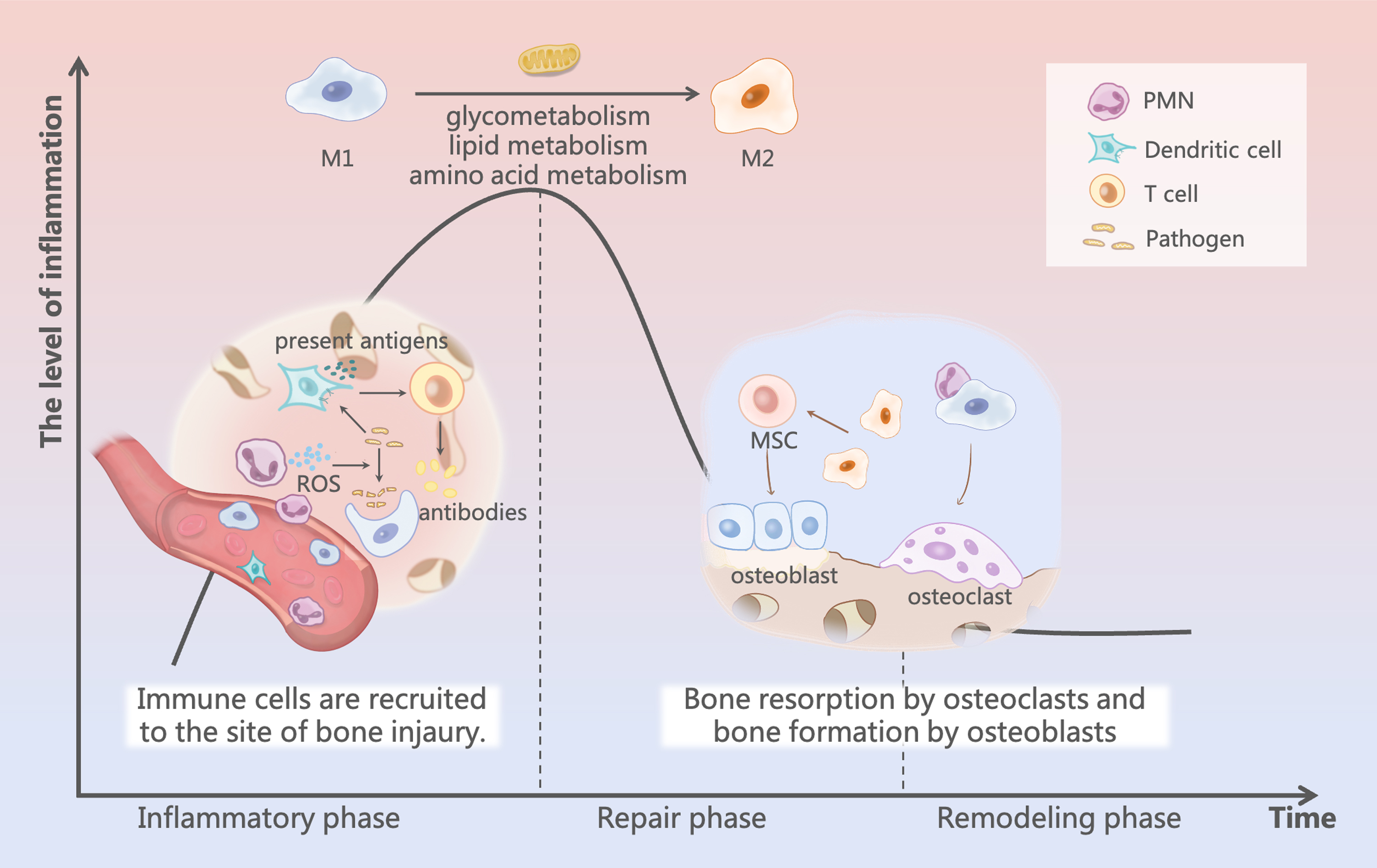
Bone healing process: during the inflammatory phase, blood vessels rupture and an inflammatory response occurs with immune cell infiltration at the site of bone injury. As inflammation subsides, changes in the microenvironment enable metabolic reprogramming of macrophages and initiate a period of repair and remodeling. Color images are available online.
The inflammatory phase is the initial stage of fracture healing and the key stage that determines final healing. When bone damage occurs, blood vessels in the bone and the surrounding soft tissue rupture, causing acute inflammation and hematoma formation. Polymorphonuclear leukocytes (PMNs) are the first cells to be recruited to fracture sites, where they release enzymes and oxygen radicals to scavenge pathogens and necrotic tissue and secrete chemokines, such as CCL2 and IL-10, to recruit immune cells, including macrophages. The active phase of PMNs lasts for ∼1 day, after which other immune cells (including dendritic, T, and B cells) are recruited as the main participating cells.
Dendritic cells can present antigens to activate T cells for an adaptive immune response, while producing chemokines to accelerate the inflammatory response, while B cells secrete IgM antibodies and remove remaining pathogens and cellular debris. Macrophages at this time can link between the preceding and the following: (1) avoiding damage caused by excessive PMNs through a cloaking mechanism; (2) secreting chemokines such as CCL2 and CXCL8 to recruit mesenchymal stem cells (MSCs) to promote osteogenic differentiation; and (3) secreting cytokines such as IL-6, IL-1b, and TNF-α to initiate osteoclast formation. 18
Inflammation gradually subsides as levels of anti-inflammatory and proinflammatory factors increase and decrease, respectively, and bone healing subsequently enters the repair phase. 7 The deposition of platelets and fibrin contributes to hematoma formation and osteogenic differentiation is initiated. Depending on the blood supply to the newly formed tissue, MSCs differentiate in different directions toward osteogenesis. 10 When neovascularization is insufficient and blood supply is inadequate, the resulting low oxygen tension in the microenvironment inhibits osteoblast activity, while being suitable for the differentiation of chondrocytes for endochondral ossification. Conversely, when the blood supply is adequate, higher oxygen partial pressure stimulates the differentiation of MSCs into osteoblasts for intramembranous ossification, which subsequently results in callus formation. 19
In addition to determining the partial pressure of oxygen in the microenvironment, neovascularization recruits MSCs and monocytes. Monocytes develop into spindle-shaped macrophages and small round mononuclear tartrate-resistant acid phosphatase (TRAP)-positive cells, which ultimately differentiate into mature osteoclasts that begin to resorb calcified cartilage and form resorption traps. 20 Osteoblasts differentiated from MSCs produce new bone to fill the resorption traps and form woven bone with trabecular structures. During this process, macrophages phagocytose ACs through efferocytosis and produce nucleotides that promote the proliferation of noninflammatory macrophages, which function as anti-inflammatory and reparative agents and regulate osteogenic differentiation and osteoclast production, respectively. 21
When the fracture gap is bridged by a callus, fracture healing enters the remodeling phase. This process requires a high degree of cooperation among osteoblasts, osteoclasts, and macrophages. During this phase, levels of most inflammatory factors, except interleukin 1 (IL-1), tumor necrosis factor (TNF), and bone morphogenetic protein 2 (BMP-2), are significantly reduced. Osteoclasts active on the outer surface resorb the woven bone, whereas osteoblasts in the fracture gap deposit lamellar bone around the central capillaries. 22 Notably, the osteoblast-to-osteoclast ratio affects the final state of the bone. Excessive osteogenesis or insufficient bone resorption can result in the formation of abnormally calcified nodules, while excessive bone resorption or insufficient osteogenesis can lead to poor bone healing.
Macrophages in the microenvironment control the balance between osteogenesis and bone resorption, regulating the ratio of osteoblasts to osteoclasts by secreting various factors to ensure an appropriate amount of bone. Under mechanical stress, bone tissues return to their original structure and shape after months or even years. 23 During this phase, macrophages in different polarization states affect the osteoblast-to-osteoclast ratio. Macrophages act as central hubs to coordinate various cells and factors that perform their respective tasks throughout bone healing.
Role of Macrophages in Bone Regeneration
Macrophages interact with the bone healing microenvironment through cytokines, chemokines, and other factors. Macrophage metabolites accumulate in the microenvironment and circulate between cells. Simultaneously, metabolites from other cells in the microenvironment act as messengers that are taken up by and alter the phenotypes and functions of macrophages. Considering the bridging role of macrophages in bone healing, this section discusses the origin and classification of macrophages and then describes the interaction between macrophages and the microenvironment, focusing on the three stages of bone healing.
Origin of macrophages
Macrophages involved in bone healing can be divided into two categories based on their origin: Tissue-resident macrophages (TRMs) and monocyte-derived macrophages. 24
TRMs differentiate from erythromyeloid progenitors of the yolk sac by formation of premacrophages, which have the capacity for self-renewal and exhibit markers such as CD169+, ER-HR3+, F4/80+, TRAP−, and VCAM-1/CD106+. 25 TRMs maintain microenvironmental homeostasis in various tissues and promote cell growth and normal tissue development. During bone healing, TRMs facilitate bone tissue repair and reduce inflammation and fibrosis, while removing ACs through efferocytosis. Three main types of TRMs exist in the bone marrow: Erythroblastic island (EBI) macrophages, osteomacs, and osteoblasts. 26 Specifically, EBI macrophages play a major role in the formation, adhesion, and clearance of expelled erythrocyte nuclei; osteomacs cover osteoblasts on the bone surface in a coronal structure, providing continuous anabolic signals to the underlying osteoblasts to contribute to bone deposition and mineralization 27 ; and osteoclasts are a highly specialized population of macrophages in the bone marrow, which are responsible for bone resorption.
Monocyte-derived macrophages, which exhibit surface markers such as Ly6c1+, ITGAM/CD11b+, CD68+, FCGR3A/CD16+, and CD14+, are initially derived from yolk sac-derived myeloid-biased progenitor cells, which differentiate into monocytes and further into macrophages after entering tissues through peripheral blood circulation. 28 When tissue is damaged, monocyte-derived macrophages are rapidly recruited to the affected area, where they initiate an early inflammatory response. Furthermore, some studies have reported that monocyte-derived macrophages can replenish TRMs, 29 but the mechanisms underlying this phenomenon and its evolution at different stages remain unexplored. As both cell types can respond to stimulation, the following introduction of macrophages contains both TRMs and monocyte-derived macrophages.
Classification of macrophages in bone regeneration
Macrophages are highly plastic and can change their phenotype in response to their microenvironments. In previous studies, the activation pattern of macrophages has been divided into two phenotypes: proinflammatory (M1) and prorepair (M2) (Fig. 3).30,31 In vitro tests have shown that M1-like macrophages, which result from stimulation with interferon γ (IFN-γ), produce proinflammatory factors such as IL-6 and TNF-α, and have an inhibitory effect on osteoblasts.32,33 M2-like macrophages, which polarize under the influence of IL-4 and IL-13, produce anti-inflammatory factors, such as IL-10, to help rebuild the vascular network at the site of damage and are involved in the mineralization of the bone matrix. 34 M2 macrophages are further subdivided into M2a, M2b, M2c, and M2d according to their different markers and functions. 35 Notably, most bone healing initially occurs through noninfectious inflammation due to the absence of “specific antigens” such as pathogens.

Membrane receptors, enzymes, chemokines, cytokines, and functions of macrophages with different polarization phenotypes. Color images are available online.
In such cases, macrophage activation often occurs through damage-associated molecular patterns (DAMPs) produced by necrotic tissue and ACs. 36 For example, dead neutrophils release DNA to form large net-like structures called NETs, which can activate macrophages with the help of Fc receptors using Toll-like receptor 9, thus exerting an immune effect. 37 In addition, the innate signal IL-4 released by basophils and mast cells in the microenvironment converts resident macrophages into a prohealing macrophage population. 38
Macrophages act as regulators in tissue healing, which requires both proinflammatory macrophages to initiate the inflammatory response for foreign body removal, and prorepair macrophages to reduce inflammation and initiate the repair phase. It would be one-sided to attribute the role of M1-like macrophages in bone healing to a negative influence and the role of M2-like macrophages to a positive influence. Furthermore, owing to the complexity of the healing microenvironment, the activation phenotypes of macrophages are diverse, and a simple description of the M1 and M2 phenotypes is insufficient to accurately describe their functions. Therefore, a comprehensive and widely accepted classification of macrophage activation remains elusive.
Role of macrophages in the bone healing inflammation phase
In the early stages of inflammation, the formation of a hematoma by ruptured blood vessels creates a local hypoxic microenvironment, followed by the production of lactic acid by macrophages through glycolytic metabolism, which lowers the local pH. This development is not conducive to progenitor cell growth and reproduction, but provides an opportunity for immune cells to produce cytokines that recruit relevant cells to initiate an inflammatory cascade response. 39 During this time, DAMPs recognized by Toll-like receptors and proinflammatory cytokines produced by local cells stimulate monocytes to differentiate into macrophages with proinflammatory and proangiogenic phenotypes, known as M1-like macrophages. The number of these cells peaks in the middle of granulation tissue formation, accounting for 40–50% of all trauma cells.40,41
During this phase, macrophages are active mediators. Under hypoxic conditions, macrophages produce pyruvate through glycolysis, leading to citrate and succinate accumulation due to an incomplete tricarboxylic acid cycle (TCA). 42 Increased succinate levels promote the expression of proinflammatory factors in M1-like macrophages, and citrate is further broken down to produce itaconic acid, which destroys microorganisms. 43
Recent studies have shown that incomplete TCA also increases ferredoxin levels in the cytoplasm after the activation of M1-like macrophages, inducing the release of RNA from mitochondria to trigger inflammation.44,45 In addition, the pentose phosphate (PPP) is enriched in M1-like macrophages, resulting in the production of nucleotides that support their proliferation. 46 Macrophages secrete inflammatory cytokines such as TNF-α, IL-1β, IL-6, IL-12, and CCL2 to enhance the inflammatory response to clear pathogens and cellular debris. Furthermore, PPP induces the production of nicotinamide adenine dinucleotide phosphate, contributing to a large increase in mitochondrial reactive oxygen species (ROS) to stabilize HIF1-α expression, to induce vascular endothelial growth factor A (VEGF-A) production to aid in wound vascularization. 47
In addition to their proinflammatory effects, macrophages in this phase also prepare for tissue regeneration and new bone formation. IL-1β and TNF-α in the microenvironment stimulate macrophages to produce nerve growth factor, which in combination with tyrosine kinase A (TrkA) stimulates innervation for the formation of new nerves in the injured bone.48,49 In terms of bone regeneration, M1-like macrophages primarily regulate MSCs and osteoclasts through the paracrine pathway. For one thing, chemokines, such as CCL2 and CXCL8, secreted from macrophages can bind to receptors and activate MAPK, PI3K-Akt, and other signaling pathways to promote the recruitment of MSC. 50
M1-like macrophages also secrete PGE2, which binds to the EP4 receptor of MSCs to activate the cAMP-PKA signaling pathway or to activate the Wnt/β-catenin signaling pathway, which can promote osteogenic differentiation. Meanwhile, M1 macrophages increase the expression of COX-2, a rate-limiting enzyme, to increase the production of PGE2 and further enhance the osteogenic effect. 51 For the other thing, cytokines such as IL-6, IL-1β, and TNF-α, secreted from M1-like macrophages, initiate the formation of osteoclasts to engulf bone fragments.11,52–54
Role of macrophages in bone healing repair and remodeling phase
During the repair and remodeling phases, macrophages polarize into a repair phenotype, known as M2-like macrophages. The main mechanisms include the monitoring and phagocytosis of ACs and the regulation of epigenetic mechanisms. 55 In vitro and in vivo experiments have shown that intracellular polyamine accumulation following phagocytosis of ACs by macrophages inhibits the expression of proinflammatory mediators such as IL-1β and IL-6. Some researchers have constructed neutrophil apoptotic vesicles that resemble the natural apoptotic process of neutrophils in trauma, and have shown that this approach effectively promotes the polarization of macrophages toward the M2 phenotype and exerts anti-inflammatory effects, consequently improving the functional recovery of rat myocardium after myocardial infarction. 56
Exploration of mechanisms underlying macrophage polarization has revealed that the phenotypic transformation of macrophages involves the metabolic reprogramming of glycans, lipids, and amino acids (Fig. 4).47,57,58 After the inflammatory response subsides, progressive neovascularization alters the hypoxic microenvironment, resulting in downregulation of the expression of glycolytic enzymes and initiation of oxidative phosphorylation by macrophages. 47 Intact TCA inhibits inflammatory factor production by macrophage and upregulates the expression of anti-inflammatory factors such as IL-10. 59 The macrophage state at this time is similar to the immune-tolerant or dormant state that promotes tissue protection and homeostasis during the repair phase. 60 When IL-4 and IL-13 stimulate macrophages to polarize to the M2 phenotype in vitro, STAT6 induces fatty acid (FA) oxidation in mitochondria and increases the expression of genes such as CD36, CPT1, and PPAR. 57

Metabolic reprogramming of macrophages during the inflammatory, repair, and remodeling phases, including glucose, lipid, and amino acid metabolism. Color images are available online.
The increased expression of CD36 induces the uptake of FAs by macrophages, followed by the transmission of FAs into mitochondria through CPT1, which enhances and maintains oxidative metabolism. The activation of PPAR inhibits the nuclear factor-κB signaling pathway, which ultimately induces macrophages to polarize to the M2 phenotype.61–63 To promote tissue repair, macrophages remove ACs through efferocytosis, which is maintained by amino acid metabolism. Studies have shown that when macrophages engulf ACs, arginine and ornithine in the ACs are catabolized by arginase 1 and ornithine decarboxylase 1, and their breakdown product, butane diamine, sustains efferocytosis. 58 Furthermore, urea and L-ornithine generated through arginine hydrolysis regulate macrophage proliferation and collagen synthesis. 64
The altered microenvironment results in a shift in the phenotype of macrophages to one that favors tissue repair, driven by metabolic reprogramming and a subsequent change in the cytokines secreted by macrophages. During the repair phase, neovascularization continues to be induced and remodeled by growth factors such as VEGF and epidermal growth factor. In addition, cytokines such as IL-10, TGF-β, and IFN-γ secreted by M2-like macrophages effectively promote the proliferation and osteogenic differentiation of MSC-osteoblast lineage cells. 30
Simultaneously, MSCs have been shown to promote macrophage viability through paracrine secretion. For example, factors such as MIP-1 and MCP-5 secreted by MSCs can significantly enhance the migration ability of macrophages and promote tissue healing 65 ; MSCs can also increase the ability of macrophages to secrete IL-10 by enhancing the production of PGE2.66,67 Furthermore, anti-inflammatory factors, such as IL-10 and IL-4, produced by macrophages inhibit osteoclast production by downregulating NFATc1, which has a protective effect on new bone.68,69
In summary, during the three phases of bone healing, macrophages exhibit different polarization states in response to different microenvironmental stimuli, which are accompanied by metabolic reprogramming. Simultaneously, macrophages in different polarization states act on cells in the microenvironment by releasing responsive cytokines, thus steadily advancing bone healing.
Macrophage-Related Bone Regeneration Biological Strategies
Owing to their strong plasticity and important role in bone healing, macrophages are considered therapeutic targets for improving bone healing. This section classifies macrophage-related bone regeneration strategies according to the stages of bone healing that they primarily regulate, detailing the temporal regulatory strategies that regulate the inflammatory, repair, and remodeling microenvironments or throughout the bone healing process (Table 1 and Fig. 5).
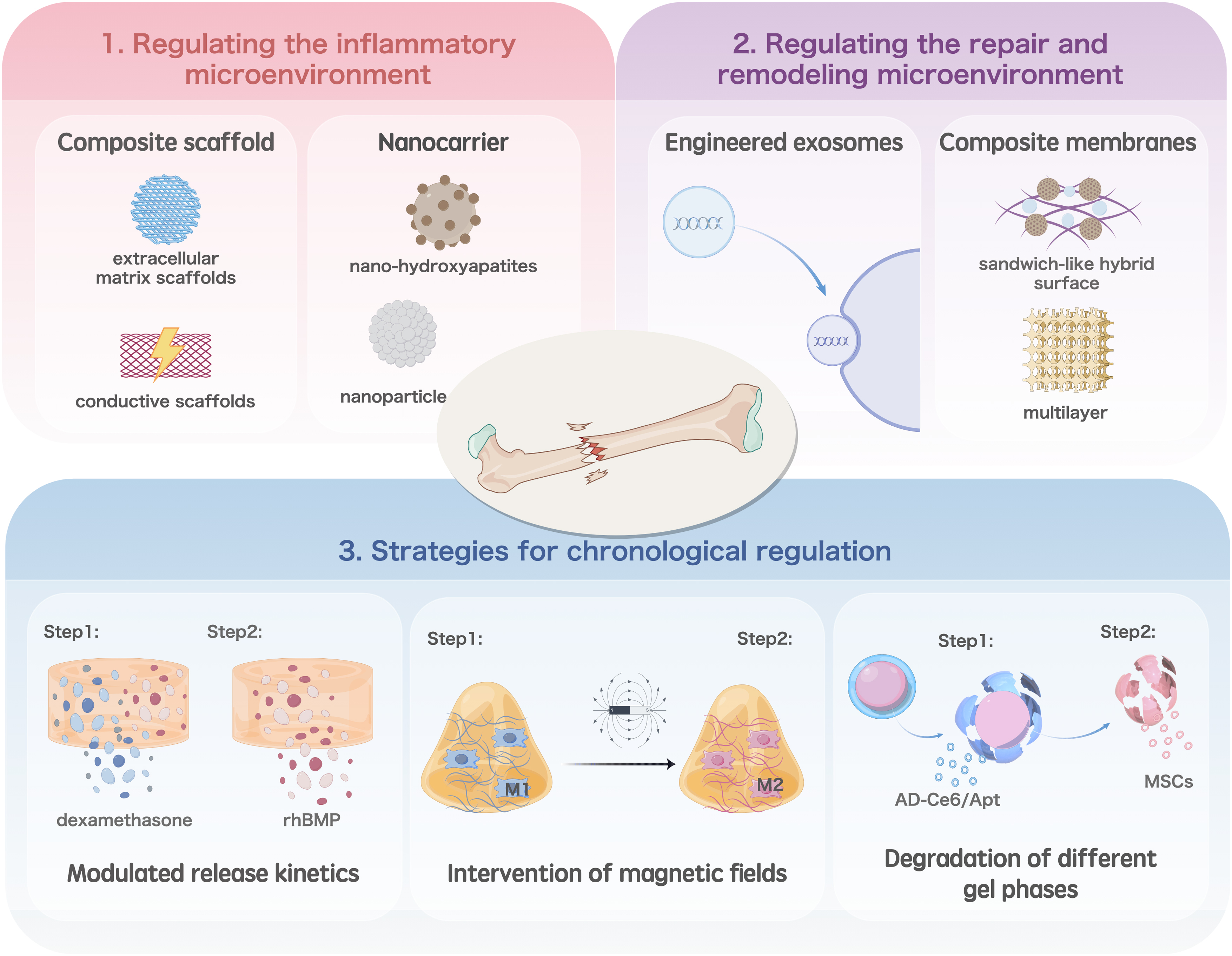
Macrophage-related bone regeneration biological strategies: (1) Strategies for chronological regulation, (2) regulating the repair and remodeling microenvironment, and (3) regulating the inflammatory microenvironment Color images are available online.
Macrophage-Related Bone Regeneration Biological Strategies
GHANP, glycosylated nanohydroxyapatite.
Regulating the inflammatory microenvironment
Regulation of the inflammatory microenvironment mainly involves the alteration of the macrophage phenotype, which allows inflammation to subside and progress to the repair phase. Deng et al. modified 3D printed poly(lactide-co-glycolide) extracellular matrix scaffolds using human umbilical cord MSC-derived extracellular matrix proteins through tissue engineering and physical decellularization techniques. 70 This scaffold contained various soluble cytokines that attenuate the immune response, while the sustained release of transforming growth factor β mediates the migration and polarization of M1 macrophages, reducing inflammation and promoting bone regeneration. Li et al. developed a composite scaffold consisting of a gastrodin-containing biodegradable polyurethane and nano-hydroxyapatite. 71 A rat implantation model showed that the composite scaffold induced the polarization of macrophages toward the M2 phenotype, significantly decreasing the formation of fibrous capsules and promoting angiogenesis and osteogenesis.
Because metabolic reprogramming plays an important role in the polarization of macrophages, some researchers have introduced it into the design of bone regeneration materials. Qiao et al. designed mannose receptor-targeted glycosylated nanohydroxyapatites to facilitate bone regeneration by enhancing mitochondrial oxidative phosphorylation and activating autophagy, which reprogrammed macrophages to the M2 phenotype and improved the bone immune microenvironment. 72 Yan et al. combined carbonic anhydrase IX (CA9) siRNA and an NO scavenger into a nanocarrier to modulate the hypoxic and acidic microenvironment by scavenging nitric oxide and inhibiting CA9 expression in macrophages, thereby reprogramming macrophages to reduce inflammation. 73
Excessive inflammatory reactions can cause tissue damage as well as lead to chronic inflammation. Inflammatory markers and ROS released during chronic inflammation inhibit osteoblast differentiation and promote bone resorption, resulting in prolonged wound healing and severely compromising bone healing outcomes in patients with underlying diseases such as diabetes. 74 As a result of being in a hyperglycemic environment, diabetic wounds contain high levels of highly ROS, which stimulate macrophages to secrete proinflammatory factors such as IL-1β, TNF-α, and IL-6, thus creating a vicious cycle of chronic inflammation. 75
Electrical signaling regulates macrophage function. To reduce the impact of inflammation on bone regeneration in patients with diabetes, Li et al. used dopamine to transform graphene into graphene oxide, which was dispersed in a network of alginate and gelatine along with hydroxyapatite nanoparticles to form a conductive immunomodulatory scaffold. 76 Dopamine in this scaffold scavenged ROS, inhibited M1 macrophage polarization, and activated M2 macrophages to secrete osteogenesis-related cytokines by mediating glycolysis and the RhoA/ROCK pathway in macrophages.
Furthermore, this conductive scaffold promotes bone regeneration by delivering endogenous electrical signals to cells and activating Ca2+ channels. Dai et al. constructed a ferroelectric nanocomposite membrane that resembled the endogenous electrical microenvironment of natural bone tissue. 77 Mechanistic studies showed that the membrane could inhibit the expression of AKT2 and IRF5 in the PI3K-AKT signaling pathway, thereby reducing the production of M1 macrophages in a hyperglycemic environment. This provides an effective strategy for designing bone regeneration biomaterials for patients with diabetes.
Regulating the repair and remodeling microenvironment
During the repair and remodeling phases, the main role of macrophages is to balance osteoblast and osteoclast activities. Therefore, the main focus in this phase is to regulate the formation of the MSC-osteoblast axis or osteoclasts through the polarization of M2 macrophages. Toita et al. successfully constructed phosphatidylserine liposomal multilayers on titanium implants through layer-by-layer assembly, which induced the transformation of M1-type macrophages into the M2 phenotype, thereby triggering the normal production and activation of osteoblasts for osteogenesis and osteoclasts for bone resorption. 78 Wu et al. loaded a Wnt signaling activator (BML) onto a polydopamine layer on the surface of a porous β-tricalcium phosphate (β-TCP) scaffold through self-assembly to form a sandwich-like hybrid surface. 79
The sustained release of BML from the sandwich-like hybrid surface significantly promoted the adhesion, migration, proliferation, spreading, and osteogenic differentiation of MC3T3-E1 cells, as well as the angiogenic activity of human umbilical vein endothelial cells. In a critical-sized cranial defect model, biofunctionalized β-TCP scaffolds may trigger a series of biological events, such as stimulation of M2 macrophage polarization, recruitment of endogenous stem cells and endothelial cells at the site of injury to provide a favorable microenvironment for bone ingrowth and angiogenesis, and inhibition of osteoclast production, thereby promoting bone healing.
The levels of certain cellular molecules affect the phenotype of macrophages and their association with other cells. Jiang et al. found that low levels of BMP-2 cannot stimulate new bone formation, but induce IL-1β release from M1 macrophages. 80 With increasing doses of BMP-2, bone marrow MSCs (BMMSCs) released IL-1Ra, which inhibited IL-1β expression. The infiltration of M1 into the local microenvironment and production of new bone tissue were excessive. Thus, the osteogenic effect of BMP-2 involves macrophage-MSC interactions that are dependent on BMP-2 dose and IL-1R1 ligands, including IL-1β and IL-1Ra.
Exosomes are extracellular vesicles formed from cytosolic granules and secreted outside the cell through cytosolic emission. 81 Taking advantage of this property, researchers have engineered exosomes to impart new properties for specific protein/RNA delivery.82,83 Recent studies have shown that macrophage-derived exosomes are rich in mRNAs and miRNAs, which can promote macrophage activation, polarization, and cytokine production. 84 Wang et al. found that BMMSCs could take up macrophage-derived exosomes and enhance the osteogenesis of BMMSCs, and detected key lncRNAs by in situ hybridization, which ultimately determined that LOC103691165 is the key lncRNA for M1 and M2 macrophages to promote osteogenesis in BMMSCs. 85
Strategies for chronological regulation
Each bone healing stage is initiated at a specific node. Therefore, an increasing number of researchers have attempted to control bone regeneration through temporal regulation. Liu et al. loaded dexamethasone and recombinant human BMP in a porous bioglass scaffold and modulated the release kinetics to inhibit the recruitment of M1 macrophages and promote M2 polarization. The scaffold promoted cartilage osteogenesis by activating the Hif-3 signaling pathway in MSCs. 86 Ding et al. constructed a fibrous scaffold that sequentially released basic fibroblast growth factor and BMP-2, which effectively promoted angiogenesis of periodontal membrane stem cells (PDLSCs) and induced polarization of M2 macrophages. 87 The immunomodulatory effects of macrophages further promote the osteogenic differentiation of PDLSCs in vitro, and ultimately, the regeneration of periodontal bone.
Most current strategies focus on polarizing M1 macrophages to M2 macrophages to prevent inflammation and enter the repair phase as early as possible. However, as mentioned above, M1 macrophages are not responsible for impeding bone healing, nor is premature entry into the repair phase necessarily beneficial to bone healing.
An early inflammatory response is essential for the processing of pathogens and ACs; therefore, it is necessary to maintain the inflammatory response for a certain period through chronological regulation and allow bone healing to enter the repair phase at the appropriate point with the intervention of specific substances. Huang et al. structured a superparamagnetic hydrogel that preserved the essential role of M1 in the inflammatory phase by delaying the switch to a static magnetic field, while later polarizing the encapsulated macrophages into M2-type macrophages in the presence of a magnetic field, thereby enhancing tissue healing. 88 In this study, macrophage polarization was precisely regulated by remote timing to optimize bone healing.
Several researchers have constructed temporal regulation models based on the degradation order of different gel phases to achieve automatic temporal ordering. Xu et al. used microsol electrostatic spinning to encapsulate IL-4 in the core structure of electrostatically spun fibers and cross-linked carboxymethyl chitosan (CMC) and type I collagen (Col-I) to the electrostatically spun fibers using genipin to form a dense CMC-collagen hydrogel. 89 This hydrogel acts as a barrier to trap the IL-4 released during the initial bursting of the mimic periosteal fibers, thereby maintaining the moderate inflammatory response of M1 macrophages and promoting the recruitment of MSCs and vascular growth during acute fractures.
As the gel phase degrades, the released IL-4 synergizes with collagen to promote polarization toward M2 macrophages, thereby doubly promoting vascular maturation and osteogenesis through the secretion of PDGF-BB and BMP-2 to reconstitute the local microenvironment. Xie et al. prepared a bilayer hydrogel of genetically engineered peptides AC10A and AC10ARGD. 90 The Ag2SQDs@DSPE-mPEG2000-Ce6/aptamer (AD-Ce6/Apt) loaded in AC10A cells induced the polarization of M1 macrophages that activated the immune system. Subsequently, AC10ARGD-containing bone marrow MSCs were dissolved to modulate M2 macrophage polarization, promoting an anti-inflammatory microenvironment.
Outlook
Macrophages play a critical bridging and regulatory role in bone healing, participating in various vital activities such as inflammatory response, revascularization, nerve regeneration, and bone regeneration. However, the specific mechanisms are still not fully understood, and a consensus on the classification of macrophages is still needed. Macrophage polarization is closely associated with changes in the bone healing microenvironment. Stimulation of different microenvironments can cause metabolic reprogramming of macrophages, including glucose metabolism, amino acid metabolism, and lipid metabolism, and improving bone healing by metabolic reprogramming may be an effective means of future clinical treatment. Furthermore, macrophages can regulate a variety of cells, such as MSCs, osteocytes, blood cells, and neural cells, through paracrine regulation, and these cells have also been shown to counteract macrophages and regulate their migration and polarization, a crosstalk phenomenon of great interest.
Owing to the importance of macrophages, bone regeneration strategies designed around them have attracted attention from researchers in recent years. In the context of bone healing, most studies have focused on regulating macrophage polarization toward the M2 phenotype to subdue inflammation and promote osteogenesis. It is important to note that an appropriate degree of inflammation is necessary for the clearance of the pre-existing bone injury site; therefore, modulation of inflammation at the appropriate time is necessary. The solution to this problem is to design temporally modulating materials that enable the phased release of different molecules to achieve modulation through the dissolution of the material or in vitro intervention. However, the influence of the environment on the solubility and precise timing control of in vitro interventions still needs to be optimized.
Footnotes
Acknowledgments
We would like to thank Editage (www.editage.cn) for English language editing. We also thank Figdraw (www.figdraw.com) for the assistance in creating ![]() .
.
Authors' Contribution
X.L. and J.G. contributed to the literature search, writing the original draft, and design of in-house figures. W.B. contributed to the literature search and design of in-house figures. J.X. contributed to conceptualization, review and editing, and funding acquisition. X.S. contributed to review and editing. Y.W. contributed to supervision. B.L. contributed to conceptualization, review and editing, and supervision. All authors read and approved the final article.
Disclosure Statement
The authors declare no competing financial interests.
Funding Information
We acknowledge support from the academic construction “Feng Yuan” cooperation project in Stomatological Hospital and College of Anhui Medical University (no. 2021kqxkFY02 and no. 2022xkfyts12).