
Abstract
The repair and regeneration of critical-sized bone defects remain an urgent challenge. Bone tissue engineering represents an exciting solution for regeneration of large bone defects. Recently, the importance of innervation in tissue-engineered bone regeneration has been increasingly recognized. The cross talk between nerve and bone provides important clues for bone repair and regeneration. Furthermore, the promotion of angiogenesis by innervation can accelerate new bone formation. However, the mechanisms involved in the promotion of vascular and bone regeneration by the nervous system have not yet been established. In addition, simultaneous neurogenesis and vascularization in bone tissue engineering have not been fully investigated. This article represents the first review on the effects of innervation in enhancing angiogenesis and osteogenesis in bone and dental tissue engineering. Cutting-edge research on the effects of innervation through biomaterials on bone and dental tissue repairs is reviewed. The effects of various nerve-related factors and cells on bone regeneration are discussed. Finally, novel clinical applications of innervation for bone, dental, and craniofacial tissue regeneration are also examined.
Impact statement
Innervation and vascularization are highly important in order for bone tissue engineering to be successful. However, there is still a lack of summary that correlates all three key concepts: osteogenesis, angiogenesis, and innervation. This article reviews the cutting-edge researches on the roles of innervation in promoting both angiogenesis and osteogenesis through biomaterials in bone and dental tissue engineering. In addition, the potential clinical application of innervation in bone, dental, and craniofacial tissue regeneration is also represented.
Introduction
Bone, a prominent constituent of the facial and dental systems, can be innervated by both peripheral and central nervous systems. 1 Intricate neural networks promote osteogenesis through various mechanisms, such as recruitment of endogenous osteogenic cells, regulating the microenvironment for bone remodeling, and secreting neurotransmitters to facilitate bone regeneration. They also promote angiogenesis by inducing endogenous endothelial cells, which increases local blood flow and the exchange of nutrients and oxygen. 1 In summary, innervation has prolonged effects on bone regeneration. The newly-formed neural networks synergize with the subsequent osteogenesis and angiogenesis processes, enhancing the repair of bone defects.
Osteogenesis has emerged as a well-researched concept in the field of bone regeneration. Since most of the current repair options for critical-sized bone defects lack efficiency and effectiveness, there has been a growing interest in exploring angiogenesis as a potential solution. 2 However, innervation as an upstream regulative cue has been relatively understudied. This is mainly because innervation occurs early during the bone regeneration process, typically beginning in the first week 3 . It is challenging to pinpoint and isolate the precise regulatory pathway within such a brief timeframe.
Bone tissue engineering is a subfield of hard tissue engineering. 4 It utilizes seed cells, scaffolds, and drugs or active factors for repairing bone tissue defects (Fig. 1). A delivery system can be used to direct various bioactive substances toward specific tissues, further promoting the repair of bone, capillary-like vessels, and nervous networks. This fosters an optimal microenvironment for the proliferation and differentiation of seed cells and recruitment of endogenous cells and molecules. 5 Unlike nervous tissue engineering, hard tissue engineering necessitates the use of a scaffold with suitable mechanical properties. Therefore, we developed a scaffold using calcium phosphate cement having a Young's modulus and compressive strength similar to those of cancellous bone. 6

Roles of innervation in bone tissue engineering. Innervation is beneficial for bone repair in bone tissue engineering.
Both osteogenesis and angiogenesis are essential elements in the regeneration of bone and dental tissues. Critical sized nerve or bone defects do not heal well without therapeutic intervention.7,8 In addition to osteogenesis and angiogenesis, these defects also require innervation to promote these processes. Engineered sensory nerves can effectively activate osteoclasts through a calcitonin gene-related peptide (CGRP)-dependent mechanism, illustrating the coordination and interplay between bones and nervous system. 9 In recent years, research has focused on the fascinating fields of neuro-bone companionship and neurovascularization.10–12
Bone regenerative microenvironment is also essential for bone and dental tissue regeneration. Through the delivery system, neurotrophins like nerve growth factor (NGF), 13 neuropeptides like CGRP, 14 and substance P (SP) can contribute to this osteogenic microenvironment. Aside from the delivery system, nerve-related cells can also positively influence bone formation. For example, Schwann cells, the classic glial cells, can secrete NGF, brain-derived neurotrophic factor, and neurotrophic factor-315,16 and, thus, facilitate bone formation.
Neurovascularized pulp regeneration is an important goal of endodontic therapy. In addition, the ideal regenerated dentin-pulp complex allows the lifetime dentin formation to protect the tooth. Until now, tissue engineering-mediated functional dentin-pulp regeneration has remained both attractive yet challenging.
Regarding bone tissue regeneration, the interrelations between bones, vessels, and nervous system have emerged as an encouraging field of research. Liu et al. provided an extensive review of the molecules and cells associated with nerves in bone tissue engineering. 17 In addition, the beneficial effects of vascularization on bone repair are widely recognized. Li et al. reported the potential of neurovascularization in craniofacial tissue engineering. 18 However, within the domain of bone and dental tissue engineering, the correlations among osteogenesis, angiogenesis, and innervation have not been sufficiently explored.
This is the first review on the role of innervation in promoting angiogenesis and osteogenesis in bone and dental tissue engineering. This review discusses cutting-edge research on the importance of innervation through biomaterials on bone and dental tissue repair. Innervation enhances both osteogenesis and angiogenesis, while nerve-related bioactive factors and cells influence the bone regenerative microenvironment. In addition, the potential clinical application of innervation in bone, dental, and craniofacial tissue regeneration was also discussed.
Effects of Nerve-Related Bioactive Factors on Osteogenesis
Nerve growth factor
NGF, the first discovered neurotrophin, regulates neuronal survival, growth, and differentiation, in addition to stimulating osteogenesis. Topical administration of exogenous NGF promotes osteogenesis and angiogenesis during mandibular fracture healing by upregulating bone morphogenetic protein-9 (BMP-9) and vascular endothelial growth factor (VEGF) levels. 19 NGF primarily signals through tyrosine kinase type A (TrkA) receptor with high affinity and also binds to the universal neurotrophin receptor p75NTR (CD271) with low affinity.
A previous study demonstrated the role of NGF in skeletal repair using mouse models with calvarial bone defects. 20 NGF expression was significantly elevated soon after the surgery (p < 0.01) (Fig. 2A). Localized nerve ingrowth occurs soon after the initial burst of NGF (Fig. 2B–D). Majority of the newly-regenerated nerve fibers were CGRP+ peptidergic sensory fibers, while the TH+ sympathetic fibers were less prominent (Fig. 2E, F). Deletion of NGF through adenovirus encoding Cre recombinase (Ad-Cre) treatment reduced innervation and vascularization, leading a delay in new bone formation (Fig. 2G–N). 1NMPP1-mediated TrkA inhibition model subsequently recapitulated this phenotype, indicating an essential role of NGF-TrkA signaling, as well as sensory innervation in bone repair.

Effects of NGF on calvarial bone repair.
Increased NGF levels and NGF-induced sensory ingrowth were also observed following ulnar stress fracture. NGF-TrkA signaling was found stimulating intramembranous healing through peripheral reinnervation, angiogenesis, and osteogenesis sequentially. 13 Both groups demonstrated a similar temporospatial process of tissue healing after bone injury. It is worth mentioning that small nerves were transected in the first study, which is more similar to conditions of tissue engineering. In addition, engineered NGF-induced sensory reinnervation was also shown to improve bone remodeling. 9 Apart from intramembranous ossification, NGF-TrkA signaling is also involved in endochondral ossification and articular cartilage calcification.21,22
NGF receptors are also expressed on bone marrow mesenchymal stem cells (BMSCs), osteoblasts, osteoclasts, and chondrocytes,23–25 indicating the local effects of neurotrophic factors on skeletal system. NGF can directly regulate bone remodeling through the differentiation of osteoblasts and osteoclasts. 26 TrkA activation inhibits BMSC apoptosis through the Akt/Bad signaling pathway. 27 Meanwhile, p75NTR was found to be essential for NGF-mediated migration of osteogenic precursors during early bone repair. 28
Interestingly, TrkA contributes to peripheral innervation by promoting Schwann cell-like differentiation of BMSCs. 25 The sustained release of NGF promotes both osteogenic and neurogenic differentiation of peri-implant BMSCs. 29 This demonstrates the intercommunicating role of bioactive factors between peripheral nerves and bones.
Applications of NGF in nerve tissue engineering have been widely accepted.30–32 NGF-induced innervation has also recently gained attention in bone tissue engineering. A silk-hydroxyapatite-based three-dimensional (3D)-printed scaffold was developed to deliver NGF to the bone formation site. In this innovative scaffold, silk is regarded as the drug delivery system. The results revealed that both NGF and VEGF enhanced osteogenic differentiation in human mesenchymal stem cells (hMSCs), although an antagonistic cross talk was observed between the two factors. 33
Constructs combining NGF through the coaxial electrospinning technique have also been proposed. Silk fibroin was adsorbed with NGF to serve as the shell of the scaffold. NGF release promotes neuronal differentiation, thereby facilitating structural and functional bone repair. 34 Furthermore, CD90/CD271 double positive mesenchymal stem cells, delivered using hydroxyapatite/PDLLA scaffolds, have been used as seed cells to promote bone regeneration. This method effectively enhanced new bone formation in vivo. 35
Calcitonin gene-related peptide
Bone is richly innervated by sensory and sympathetic nerves, which play a vital role in maintaining the delicate balance of peripheral tissues by releasing neurotransmitters. Receptors for these neurotransmitters are widely distributed in bone lineage cells. CGRP is a neuropeptide primarily released by sensory nerves, and its deficiency is associated with various conditions characterized by bone loss.14,36,37
CGRP promotes osteogenic differentiation of BMSCs and inhibits their adipose differentiation. 36 Hippo-YAP pathway was found to be involved in CGRP-induced osteogenic differentiation and BMSC migration.38–40 PPARγ signaling was shown to be disrupted in CGRP-deficient mice, resulting in compromised bone regeneration and bone loss because of the inhibition of BMP activity. 14 In addition, the absence of CGRP may lead to an increase in dysfunctional mature osteoblasts with age. 41
CGRP is widely used in bone and dental tissue engineering because of its osteogenic properties. A 3D-printed mesoporous bioactive glass/sodium alginate/gelatin scaffold loaded with CGRP was constructed. CGRP incorporation further promoted bone regeneration. 42 In addition, it has been demonstrated that a 3D-printed composite hydrogel scaffold, comprising hydroxyapatite and sodium alginate, efficiently delivers CGRP, leading to increased BMSC proliferation. 43 Borosilicate bioactive glass-alginate scaffolds incorporating CGRP also offer an improved osteogenic potential. 44 Altogether, these findings indicate that CGRP possesses the ability to functionalize bone regeneration materials. This also demonstrates the importance of sensory innervation in bone tissue engineering.
Various strategies using magnesium have been proposed to elevate CGRP levels for enhanced osteogenesis.45,46 A mechanobiologically optimized magnesium scaffold was constructed for orbital bone regeneration, which upregulated CGRP expression in trigeminal sensory neurons and promoted angiogenesis and osteogenesis. 47 Since phosphate facilitates nerve growth and CGRP secretion, the use of magnesium-ion-modified black phosphorus nanosheets was found to enhance the osteogenic effects. 48
Furthermore, a novel engineering technique has been proposed for alveolar bone repair using extracellular vesicles to slowly deliver CGRP. The vesicles were taken up by human periodontal ligament stem cells, which improved their osteogenic differentiation potential. 49 Improved osteogenic differentiation potential of CGRP gene-modified BMSCs as seed cells in bone tissue engineering has been demonstrated in vivo. 50 The application of CGRP in bone and dental tissue engineering requires further exploration.
Semaphorin 3A
Semaphorin 3A (Sema3A), an essential axon guidance factor, enables innervation by specifically targeting sensory nerves and induces osteogenesis. 51 Indeed, Sema3A has been demonstrated to act directly on bone tissue. Exogenous Sema3A exerts dual effects on both osteoblasts and osteoclasts to promote osteogenesis. 52 It has been suggested that hypothalamus and peripheral neurons express Sema3A, as do the cells in bone lineage. 53 Furthermore, osteocyte and osteoblast Sema3A has also been shown to independently regulate bone metabolism.54,55
Direct placement of Sema3A into the bone microenvironment leads to diminished biological effects due to protein instability. Therefore, the application of silicon-stimulated Sema3A secreted by sensory nerves was presented. The classic mechanistic target of rapamycin (mTOR) pathway was involved in the process of Sema3A elevation. The silicified collagen scaffold enhanced neural Sema3A production, leading to improved bone regeneration. In addition, angiogenesis and innervation were synergistically activated in this process. 53
Moreover, lentiviral vectors carrying the Sema3A gene have been used in tissue engineering to cause Sema3A overexpression in the seeded cells.56,57 However, some studies have suggested that Sema3A inhibits VEGF-induced angiogenesis. 58 In contrast, HIFα could promote angiogenesis by upregulating VEGFA expression while suppressing osteogenic differentiation. Interestingly, co-overexpression of Sema3A and HIF1α simultaneously enhanced angiogenesis and osteogenesis. The modified induced pluripotent stem cell (iPSC)-derived mesenchymal stem cells were used as seeded cells to achieve osteogenic differentiation without any changes in cell characteristics.56,57
Other nerve-related bioactive factors
SP, released by sensory nerves, is often coexpressed with CGRP. 59 The effects of SP on bone mass are dose dependent; high levels are associated with osteogenesis, whereas low levels suppress it. 60 SP deficiency increases age-related osteoclast formation and bone resorption. 61 SP-activated autophagic activity may induce osteogenic differentiation of BMSCs. 62
Although these two factors are thought to be co-released, CGRP expression is decreased, while SP expression is elevated, after nerve transection. An increase in the CGRP/SP ratio can also effectively alleviate bisphosphonate-induced osteonecrosis. 63 In addition, a negative feedback relationship between CGRP and SP has been reported. 15 Improved understanding of the interaction between SP and CGRP may provide insights for bone and dental tissue engineering.
Sympathetic signaling is known to cause bone loss. 64 Norepinephrine (NE) and neuropeptide Y (NPY) are neurotransmitters of sympathetic nerves. A recent study demonstrated that the NE receptor (β-AR) initiates transfer of osteoblast exosomal miR-21 to osteoclasts, which promotes osteoclastogenesis. 65 NPY is also negatively associated with osteogenic activity. In addition to releasing NPY directly, autonomic nervous system also regulates osteocyte NPY production through NE and acetylcholine (ACh). 66
There is now sufficient evidence to support local regulation of bone metabolism by cholinergic nerves. However, it remains unclear whether parasympathetic nerves can directly regulate bone mass. 67 ACh, released by cholinergic nerve fibers, is the main postganglionic parasympathetic neurotransmitter. Smoking-induced alveolar bone resorption is associated with ACh nicotinic ACh receptor dysfunction due to long-term nicotine exposure. 68 Vasoactive intestinal peptide (VIP), also released from cholinergic neurons, was the first discovered intestinal hormone involved in vasodilation.1,69 Emerging evidence suggests its interaction with the skeletal system. In addition, VIP biofunctionalized MeHA hydrogels have been shown to promote angiogenesis by enhancing VEGF expression during osteogenesis. 70
Effects of Nerve-Related Cells on Osteogenesis
Schwann cells
Schwann cells, specific glial cells that surround myelinated peripheral nerves, undergo changes after nerve injury to promote neural regeneration through Wallerian degeneration. Repair Schwann cells are involved in the removal of axonal and tissue debris and in NGF secretion. 18 Thus, the regenerative capacity of peripheral nerves partly depends on the phenotypic transformation of Schwann cells. 71 In addition, Schwann cell-derived exosomes (SCs-exo) is an important mediator of this paracrine pathway. 3 Studies have shown that SCs-exo promote axonal regeneration and attenuate inflammation after spinal cord injury. 72
In addition to axon terminal secretion of neurotransmitters, paracrine actions of Schwann cells are also essential for peripheral nerve-mediated bone repair. Schwann cell-derived paracrine factors, including platelet-derived growth factor-AA, parathyroid hormone, and oncostatin M, contribute to the regenerative capacity of digit tips. 73 Schwann cells from inferior alveolar nerves can induce osteogenesis of mouse skeletal stem cells through paracrine effects. 74 Moreover, SCs-exo promote migration, proliferation, and osteogenic differentiation of BMSCs in vivo. 75 SCs-exo can also mediate immunomodulation and vascularization to accelerate bone repair. 3
A 3D-printed reduced graphene oxide composite hydrogel scaffold loaded with both Schwann cells and BMSCs was fabricated to synergistically promote nerve and bone regeneration. This multicellular delivery technique offers promising opportunities for innervated bone tissue engineering. 76 In addition, a novel cell-free exo/hydrogel scaffold was also designed for bone tissue engineering, allowing sustained release of SCs-exo in the bone microenvironment. The SCs-exo group showed improved bone formation with richer neural networks. 3
Neurogenic mesenchymal stem cells
A substantial portion of the cranial skeleton originates from neural crest cells (NCCs) in ectoderm, including the frontal and interparietal parts and the sagittal sutures. 77 The remaining cranial skeleton and long bones originate from mesoderm.77,78 In fact, compared with the calvarial bones of mesodermal origin, those derived from NCCs have a greater regenerative capacity. 79 As a multipotent stem cell population, the multidirectional differentiation potential of NCCs has received attention in recent years. Although NCCs are rarely seen in adults, they are preserved as neural crest-derived cells. 80 Carr et al. first indicated the osteogenic potential of neural crest-derived mesenchymal precursor cells in adult endothelium. 73 After detaching from the nerve fascicles, these cells migrate to the injured mesenchymal tissue for osteogenic lineage differentiation (Fig. 3A–C). This osteogenic process has been shown to be dependent on intact innervation (Fig. 3D–F).
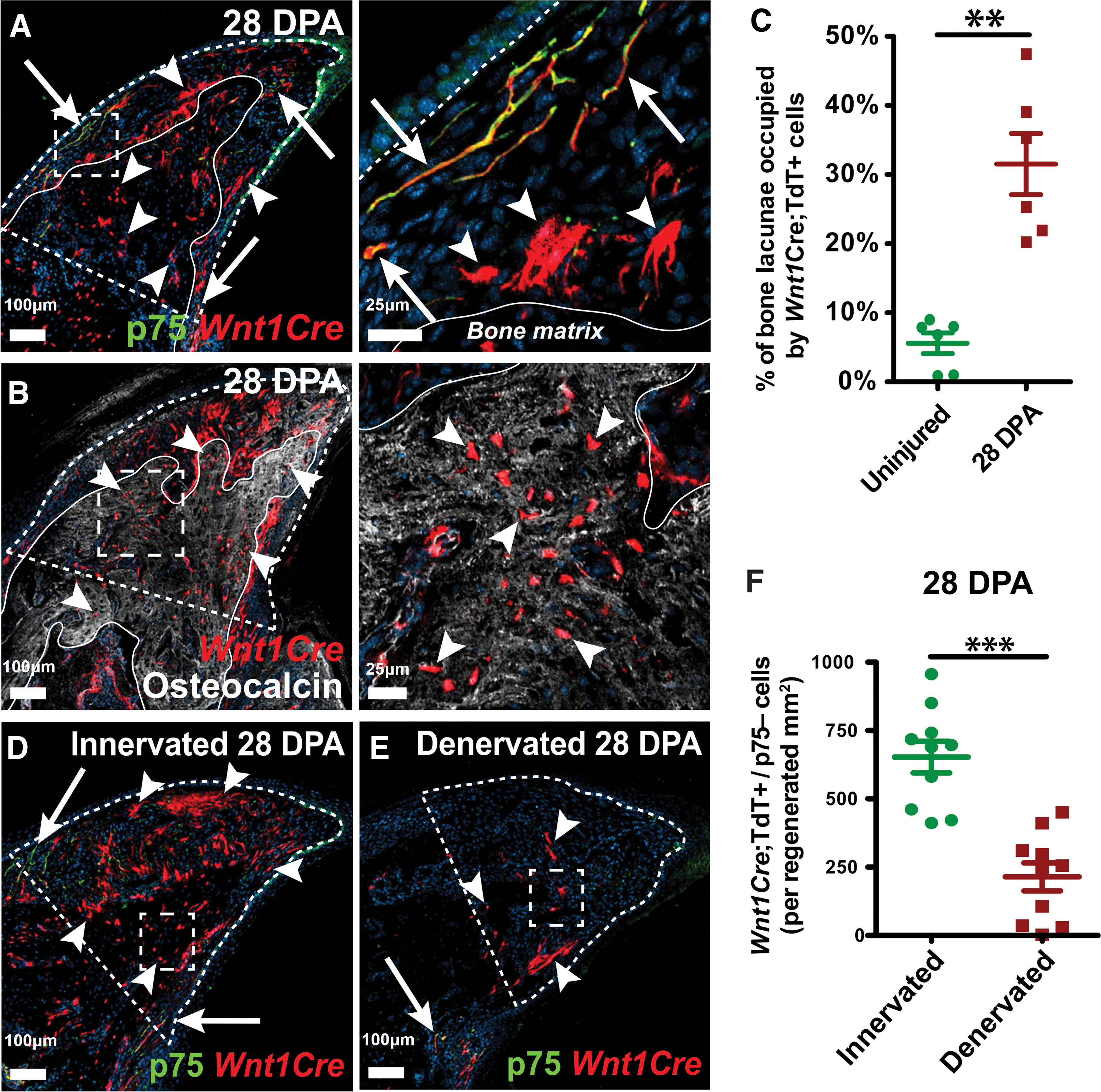
Neural crest mesenchymal cells promote digit tip bone regeneration.
Therefore, for craniofacial bone repair, iPSCs were sequentially reprogrammed into NCCs and then to mesenchymal progenitor cells (MPCs). Allografts seeded with such induced neural crest cell-MPCs (iNCC-MPCs) were found to be more potent for craniofacial bone regeneration, particularly for frontal bones, compared with those using BMSCs. The iNCC-MPC-seeded allograft exhibited higher connectivity density and bone mineral density compared to the control group, although it fell short in terms of bone volume. 77 In addition, the bone regenerative potential of NCC-MPCs combined with 3D-printable bio-ink in cranial skeleton has been demonstrated. BMP-6 overexpressing NCC-MPCs led to increased frontal bone regeneration compared with the corresponding control group. 79 Another study compared the osteogenic potential of neural crest hMSCs and BMSCs in craniofacial bone tissue engineering. The results showed that neural crest hMSCs exhibited an improved proliferative capacity. This confirmed the superiority of neural-derived hMSCs in the repair of large calvarial defects. 78
Effects of Innervation on Angiogenesis
The formation of capillary-like vessels is considered the gold standard for angiogenesis in bone regeneration.2,81 Newly-formed vascular-like tissues allow an influx of nutrients and oxygen and the removal of metabolic waste products. Although angiogenesis improves the regeneration of critical sized bone defects, the vessels retract more rapidly than they form due to instability, resulting in their disappearance. Early-stage innervation matches the early-stage angiogenesis (within the first week). 3 In addition, there is an anatomical correlation between angiogenesis and innervation. 82 These observations strongly suggest a close association between innervation and angiogenesis within the microenvironment of bone regeneration.
It was suggested that innervation serves as an initiating factor for bone regeneration and regulates subsequent angiogenesis and osteogenesis. 83 In addition, a recent study indicated that vascular endothelial cell-derived VEGF can promote neurogenic differentiation of neural cells, thus promoting osteogenic effects. 84 These findings indicate that innervation and angiogenesis may mutually influence each other.
As a potent vasodilator, CGRP stimulates bone formation through its vascular effects.9,38,85 CGRP induces the formation of type-H vessels that are required for coupling angiogenesis and osteogenesis. 86 Studies have also shown that CGRP modulates endothelial cell function by upregulating VEGF expression. 87 Furthermore, the role of CGRP-FAK-VEGF signaling axis in promoting endothelial cell migration and tube formation has been reported. 45 Similar to CGRP, SP also controls bone mass by regulating vascular formation.62,87
As mentioned previously, neural Sema3A promoted angiogenesis in the process of bone formation. 53 Furthermore, in a distraction osteogenesis model, Seme3A exerted its angiogenic effect by the Ang-1 pathway. Meanwhile, VEGF-A mediated angiogenesis was observed to be suppressed. 58
In vivo, angiogenesis manifests as early neovascularization. In vitro, vascularization often entails the migration and proliferation of endothelial progenitor cells and formation of tubular structures. 88 Bone tissue engineering utilizing SCs-exo promoted angiogenesis in rat bone defects. 3 CD31 and a-SMA are specific markers that indicate neovascularization. Immunohistochemical staining of CD31 and a-SMA increased in the SCs-exo group at 1 week. In addition, the number and area of new blood vessels also increased significantly (p < 0.01) (Fig. 4A, C). At week 4, mature and larger diameter vascular-like tissues began to form in the SCs-exo group (Fig. 4B, D). Moreover, in vitro tube formation quantitative tests demonstrated a greater number of nodes and segments in the SCs-exo group.
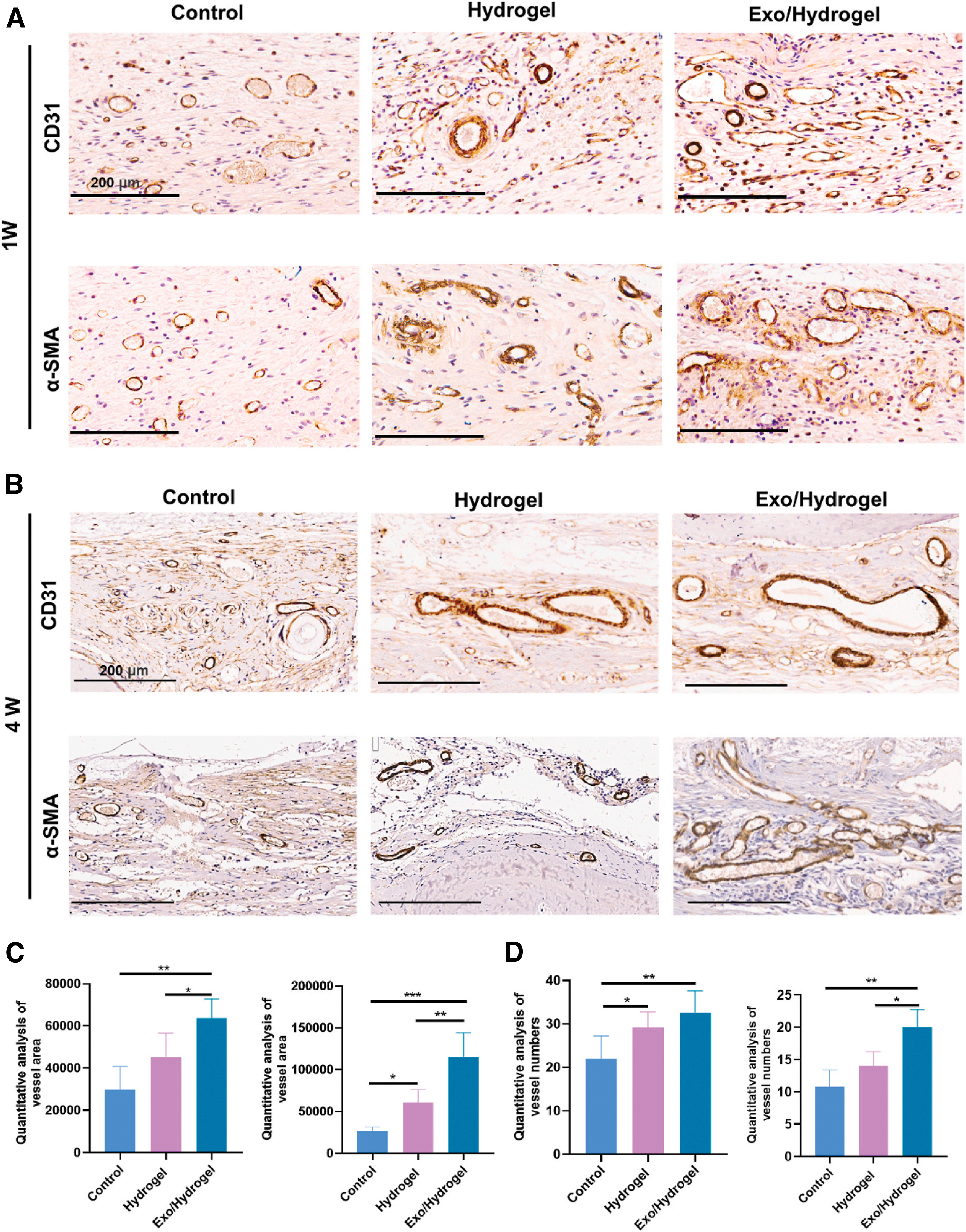
Role of innervation in angiogenesis.
Innervated and vascularized tissue-engineered bone grafts using Schwann cells with induced endothelial cells (IECs) and induced osteoblasts as seed cells have shown good osteogenic results. Schwann cells promote osteogenesis through vascularization by interacting with IECs. Schwann cell-derived VEGF stimulates endothelial cell proliferation, migration, and tube formation by upregulating IEC nestin expression. 16 It was also shown that SCs-exo exert its angiogenic effects by stimulating the migration and differentiation of endothelial progenitor cells. 88
Innervation and Vascularization in Dental Tissue Engineering
The dental pulp is a highly vascularized and innervated tissue that plays a crucial role in maintaining the vitality and functionality of the tooth. Traditional root canal therapy aims to remove the inflamed or necrotic pulp, resulting in a pulp cavity that lacks of nerve and blood vessel supply. This can make the tooth more brittle. 89 Achieving functional dentin-pulp complex regeneration is the ultimate goal of endodontic therapy. Although significant strides have been made, great efforts are still needed in the field of tissue engineering-mediated pulp regeneration.
Dental pulp stem cells (DPSCs) exert crucial function in differentiating into odontoblasts and pulpal fibroblasts. 90 It was shown that combined with decellularized tooth matrix, DPSC aggregates secrete exosomes that promote odontogenic and angiogenic differentiation of DPSCs. This leads to the formation of an odontoblast-like layer and innervation within the vascularized dentin-pulp complex (Fig. 5). 91 In addition, in vivo studies have shown that iPSC-derived neural crest-like cells (iNCLCs) have the potential to regenerate a vascularized dentin-pulp complex. Through odontoblastic differentiation induced by BMP-4, iNCLCs can create a new dentin layer. 92 Similarly, after performing pulpotomy on the first molar in rats, BMSCs were delivered into the pulp cavity using poly L-lactic acid scaffolds. This approach resulted in the regeneration of both nerves and thin dentin bridge-like structures. Furthermore, it was observed that CGRP and SP expression were significantly upregulated at days 7 and 14 (p < 0.05). 93

Bioengineered tooth implantation facilitates functional regeneration of the dentin-pulp complex.
In the field of dental tissue engineering, several biomaterials with odontogenic effects have also been identified. Neural EGFL-like 1 (Nell-1), an osteogenic factor, has demonstrated to promote the regeneration of dentin and alveolar bone in vitro and in vivo. 94 However, the impact of Nell-1 on dental nerve regeneration has not received much attention, despite its close association with neural development and disorders. Moreover, lithium was found involved in the process of dentin deposition and mineral loss through WNT/β-catenin signaling pathway. 95 It was shown to promote odontogenic differentiation and mineralization of DPSCs in a dose-dependent manner. 96
Clinical Applications of Innervation in Bone Tissue Regeneration
The absence of innervation in regenerated bone correlates with diminished structural and functional recovery. 34 The objective of promoting innervation in bone defects is to stimulate osteogenesis and enhance the overall quality of life for patients. Although neuro-bone tissue engineering is an innovative approach for addressing critical size bone defects, its implementation in clinical practice remains limited. The challenge lies in effectively promoting neurogenesis during the bone healing process. 76 Careful selection of seed cells and factors, particularly those with ability to facilitate innervation, is imperative for successful tissue-engineered bone regeneration.
Various types of stem cells have been used for innervated bone tissue engineering. Co-loading of Schwann cells and other stem cells effectively enhances regeneration of both nerves and bones. 76 Moreover, the capability of iPSCs to differentiate into NCCs holds great potential for cranial regeneration, providing promising avenues for further research. 79
Efforts to enhance the levels of nerve-related active factors within the osteogenesis microenvironment continue to attract attention. Several approaches have been suggested, including immersion loading, overexpression, extracellular vesicle delivery, and endogenous induction.47,49,50,57,97 Magnesium ions have been demonstrated to not only stimulate CGRP expression but also improve Schwann cell function, leading to the upregulation of NGF and neurotrophic factor-3 expressions. 98 Moreover, given the instability of proteins, peptide substitution is gaining popularity as a means to achieve sustained levels of active factors in the defect area. 18 In recent years, cell-free tissue engineering techniques involving exosomes have emerged as an area of interest. These exosomes can be released steadily over time and can simultaneously enrich numerous secretory factors. 3
Current research is focused on addressing numerous practical issues. For instance, studies have revealed that endochondral ossification can aid in the formation of tissue-engineered bone,99,100 particularly for nonweight-bearing bones with limited angiogenic capacities. In addition, CGRP can act as an anti-inflammatory bioactive agent, effectively enhancing the biocompatibility of biomaterial scaffolds. 44 This makes it particularly suitable for infection-prone cases, such as orbital and alveolar bone repairs. However, the long-term strength and safety of tissue-engineered bone remains uncertain. High costs and the complexity of fabrication further hinder its clinical application. Future research must address these obstacles and investigate the interplay between various factors and cells in large animal models.
Conclusions
Innervation plays a crucial role in the repair and regeneration of bones and dental tissue. This comprehensive review was the first to explore the intricate connections between neurogenesis, angiogenesis, and osteogenesis, focusing specifically on bone and dental tissue engineering. The role of innervation in angiogenesis and osteogenesis of critical size bone defects has gradually gained recognition. Researchers have investigated the application of innervation in bone and dental tissue engineering, affirming its effectiveness for stimulating bone formation. Various nerve-related bioactive factors have been identified that either stimulate or suppress bone formation. The downstream molecular mechanisms of these factors are intricate and multifaceted. Nerve-related cells are also fundamentally involved in the bone repair process. In addition, we discussed the specific applications of these factors and cells in the field of bone and dental tissue engineering. Further research is necessary to explore the application of innervation in different types of bone defects.
Footnotes
Authors' Contributions
Conceptualization, M.Z., K.Z., and L.X.; investigation, Z.D., M.D.W., A.S., T.W.O., K.Y., J.M., and Q.Z.; writing—original draft preparation, L.X., M.Z., K.Z., and J.M.; writing—review and editing, K.Z., Z.Z., and H.H.K.X.; visualization, L.X., M.Z., K.Z., and K.Y.; supervision, K.Z.; and funding acquisition, Y.B. and K.Z. All authors have read and agreed to the published version of the article.
Disclosure Statement
The authors declare that there are no conflicts of interest.
Funding Information
This work was supported by grants from the Innovation Research Team Project of Beijing Stomatological Hospital, Capital Medical University (grant No. CXTD202203 to Y.B.) and the Capital's Funds for Health Improvement and Research (grant No. CFH2020-2-2142 to K.Z.).