
Abstract
After tendon or ligament reconstruction, the interface between the hard bone and soft connective tissue is considerably weakened and is difficult to restore through healing. The tendon/ligament–bone interface is mechanically the weakest point under tensile loading and is often the source of various postoperative complications, such as bone resorption and graft laxity. A comprehensive understanding of the macro- and microfeatures of the native tendon/ligament–bone interface would be beneficial for developing strategies for regenerating the tissue. This article discusses the structural, biological, and mechanical features of the tendon/ligament–bone interfaces and how these can be affected by aging and loading conditions.
Impact statement
This review provides an up-to-date summary of the structural, biological, and mechanical features of the tendon/ligament–bone interfaces, and how these can be affected by aging and loading conditions. A thorough understanding of these features provides critical foundation for developing advanced techniques for ligament/tendon reconstruction and soft–hard tissue interface engineering.
Introduction
Ruptured ligaments or tendons are typically treated by reconstructing the tissue with grafts,1,2 often with a soft tissue graft secured in bone tunnels drilled into the connected bones. However, bone tunnel enlargement and graft failure are commonly reported after surgery, which both reflect a failed healing of the graft–bone interface.3–6 Although studies have reported suspected graft maturation and healing within the bone tunnel on magnetic resonance images, systematic reviews have shown that the healing tissue at the graft–bone interface is the scar tissue with drastically different structural and mechanical properties to the native tissue.6,7
The native tendon/ligament–bone interfaces have a graded microstructure 8 with at least four zones at the cartilaginous interface: tendon/ligament, uncalcified fibrocartilage (UFC), calcified fibrocartilage (CFC), and bone. The gradual transition of tissue calcification is considered to reduce stress concentrations at the tissue interface and reduce the risk of damage. Some studies have attempted to use tissue engineering to develop graded materials with biomimetic functionality to improve the performance at the graft–bone interface. 9 However, most trials have focused on achieving a simple and distinct material gradation and neglected other factors such as the quantity and spatial distribution of the structural and biological features to mimic the mechanical function of specific tissues.9,10 A comprehensive understanding of the structure–function relationship at the native soft–hard tissue interface is critical for guiding a better development and regeneration of this special organ.
Previous review articles (in 2002 and 2014) highlighted the current knowledge on tendon enthesis and mainly focused on the basic morphology, with some comments on potential functions.11,12 In the subsequent many years, there have been many studies using more advanced investigation techniques on this topic. The current review article assesses the most current evidence-based studies on qualitative and quantitative characterizations of the structural, biological, and mechanical features of the ligament–bone and tendon–bone interfaces, as well as the potential association between the structure and function of the interface. This is followed by a discussion of research gaps and suggestions for future studies. By offering a thorough understanding of the interface, this review article may help to improve the graft design and promote the use of novel materials that can more closely mimic the native tissues.
Methods
Information sources and search strategy
A comprehensive literature search was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses statement. 13 Studies were identified by searching the following electronic databases (until March 30, 2023): Web of science, PubMed, and Scopus. The literature search was performed using the search terms “(“ligament-bone” OR “tendon-bone”) AND (“interface” OR “junction” OR “insertion” OR “enthesis”),” and only publications in English were considered.
Study selection, quality assessment, and data extraction
After removing duplicates, the remaining literature was filtered through two stages. First, a preliminary screening was conducted by title and abstract. The eligibility criteria were (1) in vivo or in vitro experiments characterizing the structural or biological or mechanical features of the tendon–bone or ligament–bone interface, and (2) studies exploring the structure/biology–function, structure/biology–age, and structure/biology–loading relationships at the ligament/tendon–bone interface. Review articles, letters, comments, practice guidelines, and retracted articles were excluded. Irrelevant studies were also excluded. A quality assessment was carried out on the included articles by screening the full text in the second stage, and potentially relevant studies were retrieved and screened by reviewing the list of references.
The quality of the selected studies was assessed in terms of study design, sample size, species of the sample, and the adequacy of reporting on the techniques used. Two independent reviewers conducted the data extraction. The following information was extracted from each included study: (1) level of evidence, (2) sample size, (3) species, (4) tissue type, (5) main methodology, and (6) main objectives. Any disagreements were resolved by consensus, and if necessary, the corresponding author was consulted to make a final decision.
Results
Study selection
The literature search generated 2058 relevant citations in total from the 3 databases (Fig. 1). After adjusting for duplicates, 1234 articles were left and screened by the title and abstract. Forty-six were left and retrieved for full-text assessment. One study was excluded because the same work was presented in two different journals. Additional eight studies were included by reviewing the reference lists of the eligible studies. Fifty-three studies in total were finally included in this review.

The flow diagram illustrating the study selection.
Study characteristics are presented in Table 1. Most of the included studies are observational laboratory studies. 8/53 of the studies did not declare a sample size. Most of the studies (70%) were conducted on animals, and different tissues were used as the subjects, with the anterior cruciate ligament (ACL)–bone interface being the most investigated tissue. Most of the studies (44/53) explored the biological and morphological features of the interface tissues, while fewer assessed the mechanical properties and functionality (9/53) or age-related changes (12/53).
Study Characteristics
Null represents that the sample size was not declared in that study.
ACL, anterior cruciate ligament; ALP, alkaline phosphatase; CFC/CF, calcified fibrocartilage; CFU assay, colony-formation unit assay; COMP, cartilage oligomeric matrix protein; CT, computed tomography; DIC, differential interference contrast; ECM, extracellular matrix; EDXA, energy dispersive X-ray analysis; F-CPH, fluorescein-labeled collagen-hybridizing peptide; FFT, fast Fourier transform; FIB, focused ion beam; FTIR-I, Fourier transform infrared spectroscopic imaging; GAG, glycosaminoglycan; H&E, hematoxylin and eosin; IHC, immunohistochemistry; IP, imaging plate; LCL, lateral collateral ligament; MCL, medial collateral ligament; MPCM, multiphoton confocal microscopy; OCT, optical coherence tomography; PCL, posterior cruciate ligament; PL, patellar ligament; QT, quadriceps tendon; RT-PCR, reverse transcription-polymerase chain reaction; SEM, scanning electron microscopy; TEM, transmission electron microscopy; TOF-SIMS, time-of-flight secondary ion mass spectrometry; UFC/UF, uncalcified fibrocartilage.
Because the study design, sample species, and reported outcomes varied markedly, the current study focused on describing the main findings and limitations and making a qualitative synthesis rather than a meta-analysis.
Basic structural features of the tendon/ligament–bone interface
Dolgo-Saburoff 14 and Schaffer 15 were pioneers in this field to report a four-zoned structure of the fibrocartilaginous tendon–bone interface: ligament, UFC, CFC, and bone (Fig. 2a). Later, Biermann 16 and Knese and Biermann 17 distinguished three types of muscle attachments: (1) apophyseal/epiphyseal attachment with cartilaginous apophyses between the tendon and bone (Fig. 2a), (2) diaphyseal attachment with fleshy attachment to the periosteum (Fig. 2b), and (3) diaphyseal attachment with circumscribed tendinous attachment to bony crests (Fig. 2c). The epiphyseal attachment was reported to have a four-zoned structure. Cartilage may be present at the circumscribed diaphyseal attachments, but it is not as prominent as in apophyseal tendons. The diverse attachments can also be found at ligament–bone interfaces.
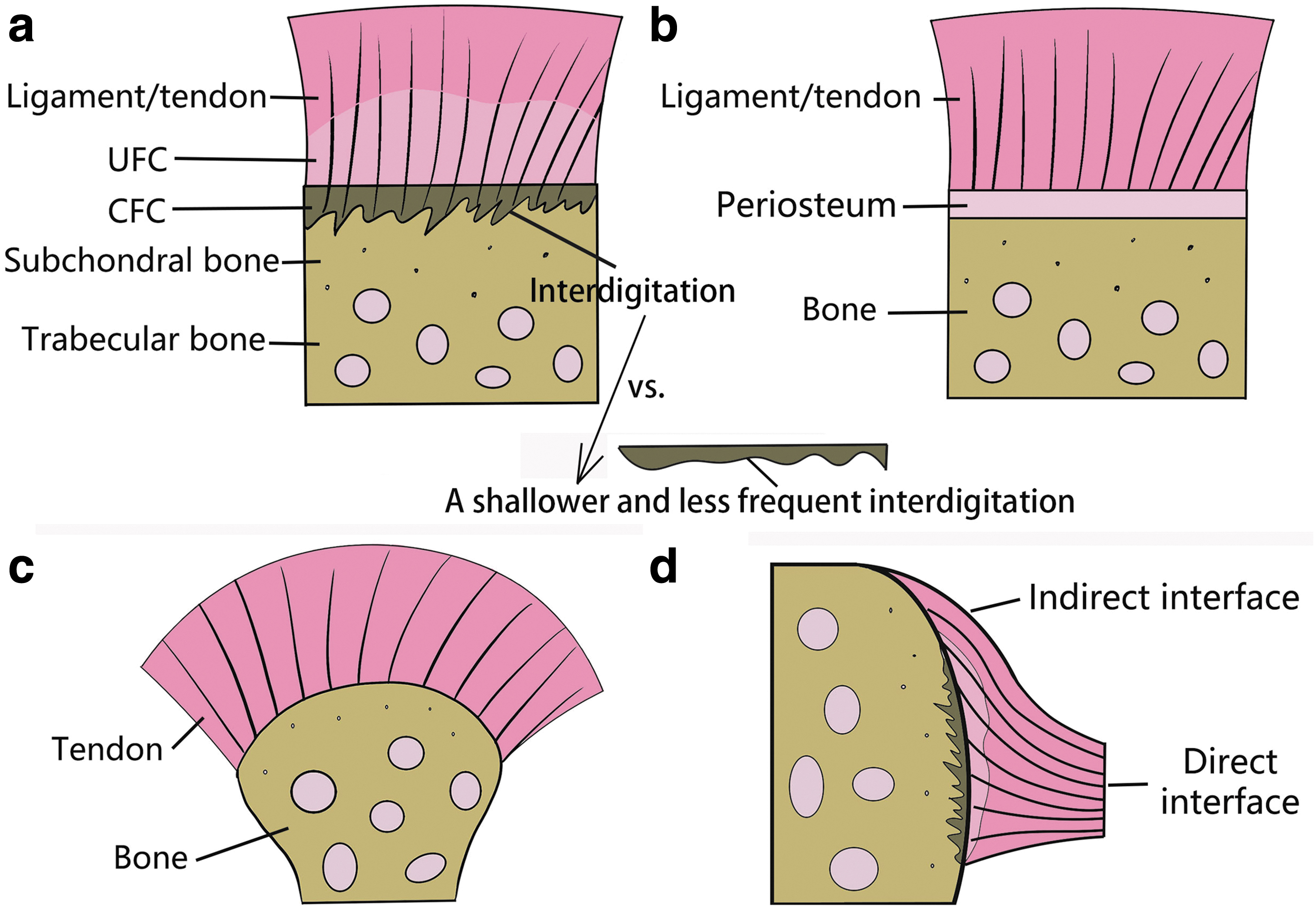
Different types of ligament/tendon–bone interfaces.
In some cases, a single ligament may even have one type of interface at one end and another type at the other, such as the medial collateral ligament (MCL). The MCL was reported to have an epiphyseal interface on the femoral side, but a mainly fibrous attachment on the tibial side. 18 Furthermore, it was found that 19 the femoral insertion of the MCL consisted of two types of insertions, with the superficial portion attached via a periosteum to bone while the deep portions were cartilaginous. Messner 20 reported that the most posterior fibers of the femoral insertion of a rat ACL were continuous with the periosteal fibers. This description of the “posterior fibers” corresponds to the concept of a “fan-like extension” or “indirect enthesis” proposed later on by other researchers 21 (Fig. 2d).
Since the connection of the direct and indirect insertions of the ACL to the femur is obviously different, it was hypothesized that the two types of insertions may play quite different roles in anchoring the tissues. 22 However, differences in the microstructure, biological contents, and mechanical function between the two types of interfaces (direct and indirect insertions) remain unclear.
To date, most microscopic studies of ligament/tendon–bone interfaces focused on four-zoned epiphyseal/cartilaginous interfaces (Fig. 2a). In the body of ligaments and tendons, the fibroblasts are elongated when aligned along the collagen fibers, while the chondrocytes in the UFC zone are spherical shaped. The CFC zone contains hypertrophic chondrocytes surrounded by an extensive pericellular matrix, and bone tissue is the last region along the insertion axis in which there are round or oval osteocytes embedded in the bone matrix. 23 The boundary line between the UFC and CFC zones is termed the tidemark, and the division between the CFC and bone is the cement line. The tidemark is basophilic and is continuous with the tidemark of the surrounding articular cartilage. 24 The tidemark has a much smoother contour than the cement line.
Some studies19,24 reported that collagen fibers are recognizable up to the irregular cement line. However, others8,25 found that collagen fibers did not stop at the cement line, but rooted deep into the bone.
Basic biological and biochemical features
Collagen is responsible for maintaining the structural integrity of connective tissues. 26 To date, collagen types I, II, III, V, VI, IX, X, XI, XII, and XIV have been found at the tendon/ligament–bone interfaces (Fig. 3a). Type I collagen was found to primarily locate in the bone and ligament regions.27,28 Type VI collagen was reported to be the main component of nonfibril collagen in ligaments, while type XIV was found predominantly in the fibrocartilaginous zone. 27 Type III and V were the main fibrillar collagens found in the ligament proper. Nawata et al. 29 identified type III collagen at pericellular regions and along the surface of the ligament and concluded that type III collagen is often present in tissues requiring high levels of mechanical compliance. Type II, IX, and XI collagens were identified in the fibrocartilaginous zones. 30

Distribution of
Type II collagen was prominent in the UFC zones and to a lesser extent in the CFC zone, but none was found in the bone and ligament regions. 28 Type II collagen contained half of the hydroxypyridinium crosslinking residues found in the collagen isolated from hyaline cartilage, indicating that the ligament–bone junction is cartilaginous in nature. 30 Type IX shows a similar distribution to type II but is present in much smaller quantities.25,31 Type X collagen is located predominantly in the CFC zone and is synthesized by hypertrophic chondrocytes, and thus is assumed to play a role in forming scaffolds at the ligament insertion for mineral deposition to maintain this zone in a mineralized state.23,32,33
Proteoglycans serve a vital role in regulating the nature of the extracellular matrix. 34 Keratan sulfate was found to be limited to the fibrocartilaginous zone (Fig. 3b) in older animals and was considered to inhibit fibrillogenesis, which may explain why the fibers in the UFC zone have a smaller fiber diameter than the ligament proper. 18 Chondroitin and dermatan sulfates were also mainly found at the fibrocartilaginous zones and were considered to assist the enthesis with resisting pressure. 18 Waggett et al. 34 found a complimentary distribution of versican and aggrecan. Versican was present in the midtendon while aggrecan was mainly present in the fibrocartilages. Decorin was identified in the ligament and fibrocartilaginous regions, and the cartilage oligomeric matrix protein was shown to primarily surround cells at the cartilaginous region. 23 Strong alkaline phosphatase activity was found in the bone region, with slight but positive staining in the fibrocartilaginous region.
It was shown that a gradient of mineral content exists at the tendon/ligament–bone interface to facilitate load transmission without developing stress concentrations. 35 Previous study 23 found dense activity of phosphate at the bone region and around the hypertrophic chondrocytes in the CFC zone, with no phosphate detected in the ligament or UFC region (Fig. 3c). The strongest alkaline phosphatase (ALP) activity was found at the bone region.
Blood supply
Sasaki et al. 21 reported a well-vascularized fibrous membrane covering the ACL. Benjamin et al. (Fig. 3d). 24 reported that blood vessels are absent from the fibrocartilage zones of the tendon–bone interface. However, Visconti et al. 30 reported blood vessels in the deep layer of the MCL–femur interface, which is composed of a mixture of CFC and bone, which is consistent with the studies by Wei and Messner 36 and Wang et al. 23
Quantitative characterization of structural features
Characterizing the morphological features of the native tendon/ligament–bone interface can provide a microstructural basis for developing biomimetic scaffolds to recreate the tissue.
Table 2 summarizes the previously measured thicknesses of different zones of the tendon/ligament–bone interface. The UFC zone was reported with a thickness of between 41 and 1386 μm,24,37–41 and the CFC zone has a thickness of between 101 and 580 μm.8,37,38,40,41 The thickness of the subchondral bone plate has a range of 111–179 μm, 40 and the thickness of the (UFC + CFC) and the (CFC + subchondral bone plate) zone was reported to be within 356–78023 and 205–800 μm,21,24,40 respectively. Most studies reported the thickness of the UFC zone to be larger than the CFC zone.19,38–41 Consistent with the previous qualitative observations, 42 Evans et al. 39 reported a larger quantity of UFC in the deep region of the quadriceps tendon insertion than the shallow region, indicating that the UFC is thicker in regions with more mobile fibers.
Quantitative Characterization of the Thicknesses of Different Zones in the Tendon/Ligament–Bone Interface
However, significantly less total calcified tissue was found in the deep regions than the superficial regions, 40 which signifies that the superficial regions bear more load than the deep region. It may be concluded that the UFC and CFC have a complementary trend in thickness variation. However, this is in contrast with results from Dai et al., 38 who concluded that the thickness of UFC and CFC had a consistent trend of variation. Whether the load level or the fiber mobility plays a more dominant role in modulating the interface thicknesses is not clear.
The shape of the interdigitation of the cement line into subchondral bone (Fig. 2a) is also potentially related to local loading conditions at the soft–hard tissue interface. Gao and Messner 19 reported that among the MCL, patellar ligament, ACL, and posterior cruciate ligament (PCL), the interdigitations at the MCL insertion were shallower and less frequent than the other insertions. Thus, the MCL was assumed to experience lower tensile forces than other ligament insertions. This, however, appears to conflict with results from Woo et al. 43 who found that the MCL was stronger and had a higher tensile failure load than the ACL. Zhao et al. 25 reported a relatively shallower interdigitation of the posteromedial (PL) bundle of the ACL insertion than the anteromedial (AM) bundle.
Previous studies showed that the PL bundle plays a greater role in restraining tibial rotation than the AM bundle, which mostly functions to restrain anterior tibial translation. 44 The above study thus proposed that the differences in microstructure of the two bundles might reflect their macrolevel differences in biomechanical function, but the mechanism behind this correlation is not clear.
Changes in the size of collagen fibers may have an effect on the mechanical properties of the tissue and reflect the functional adaptations to physiological loads. Larger fibers might be stiffer and more resilient to loading. Qu et al. 45 reported a larger diameter of collagen fibers in the fibrocartilaginous and bone regions than the ligament region. However, Rossetti et al. 28 found that fibers in the tendon proper unravel into thinner fibers in the UFC zone. It was hypothesized that a thinner fiber may have a larger area of force transmission and reduce stress concentrations. Consistently, Sartori and Stark 46 reported a smaller fiber cross-sectional area at the Achilles tendon enthesis when comparing with the tendon proper. However, regardless which results represent a more accurate picture of physiological conditions, the exact mechanism driving the differences in the fiber diameters between different gradient zones is not yet clear.
The orientation of fibers and bony trabeculae at the interface may reflect the direction of load transmission and corresponding remodeling of the interfacial tissues. Chandrasekaran et al. 47 reported that there was a more uniform fiber orientation from the tendon to the bone. Beaulieu et al. 37 reported that the ACL fibers had a more acute angle at the femoral insertion than the tibial side, which may reflect the distinguished direction of load between the two sides. Suzuki et al. 41 reported that the orientation of the bony trabeculae was prominently parallel to the ACL fiber and this feature was more intense on the tibial side. This may be caused by the fact that the femoral side receives more multidirectional loads than the tibial side during knee movements.
Mechanical properties and adaptation to different loading conditions
Moffat et al. 48 conducted compressive testing combined with digital image correlation on the ACL–bone interface and reported that the strain in the UFC region was greater than the CFC region. This depth-dependent mechanical inhomogeneity was found to be correlated with the gradation in mineral content. Besides, the compressive Young's modulus of the tibial UFC and CFC was greater than the femoral side, which also corresponded to a greater mineral content on the tibial side. However, no studies to date have quantitatively assessed the tensile properties of the tendon/ligament–bone interface, which is important to consider since the tendons and ligaments mostly function to resist tensile forces.
Several studies provided a limited qualitative evaluation of the tensile properties of the tendon/ligament–bone interfaces. Gao et al. 49 conducted a load-to-failure tensile test on several knee ligament entheses at low strain rates and found that the failure modes were elaborate, involving different degrees of failure to all microregions. Golman et al. 50 reported that a loading-to-failure regime caused the energy being accumulated within a short time to avulse the bone at the tendon enthesis, while in cyclical loading, energy was continuously absorbed by tendon and enthesis to eventually cause failure of these tissues. Besides, higher strain rates were found to increase the area and number of the avulsed fragments and the measured strength and toughness of the enthesis.
Decreased abduction angle of the supraspinatus tendon enthesis was found to decrease the measured strength and stiffness but increase the toughness value. Removal of mineral was shown to decrease the strength, stiffness, and toughness of the tendon enthesis, when removal of proteoglycans decreased the strength and stiffness but did not change the toughness of the enthesis. Under monotonic loading, overuse of the enthesis was shown to have little postyield behavior and failed primarily with one bony avulsed fragment, when underuse of the enthesis showed lower failure load, distinct postyield behavior, and increased fracture area. Both overuse and underuse were shown to reduce the enthesis toughness.
It was also reported by Golman et al. that overuse did not affect enthesis strength but led to an increase in stiffness, when underuse led to a decrease in both strength and stiffness. Besides, overuse was reported to result in more failures at the CFC–bone interface, while underuse resulted in more failures at the bone–tendon interface. Besides, Golman et al. 51 found that the energy-storing enthesis prioritizes toughness over strength, when the “positional tendons” prioritize consistent stiffness. The bony anatomy dictated fiber recruitment and the bone ridge curvature dictated the enthesis strength. Spalazzi et al. 52 used ultrasound elastography to investigate the response of the bovine ACL–bone interface to tensile force. Lower strain was found near the insertion site when compared with the midsubstance. Besides, both compressive and tensile strains were identified throughout the whole bone–ligament–bone complex.
Boys et al. 53 used confocal elastography to assess bovine ligament insertions and found that under a tensile load, the strains were highly localized to the ligament region of the meniscal attachments and there was a logarithmic increase in strain from the UFC to the ligament region. Rossetti et al. 28 reported that the mechanical response of the tendon enthesis is highly heterogeneous along the interface, and the mechanical property showed a dependence on the force angle, which reflected a different recruitment of fiber bundles with a different angle of force application.
A few studies have explored the adaptation of enthesis morphology with different loading environments. Takahashi et al. 54 found that with lower muscle contraction forces at the entheses, the CFC stopped lengthening with time and the collagen fibers became disorganized. Thomopoulos et al. 55 reported that decreased loading resulted in delayed development of fibrocartilage and decreased bone volume. Castro et al. 56 reported that the cross-sectional areas of four tendon entheses of mice increased with higher activity levels. Deymier et al. 57 reported a decreased size and alignment of mineral crystals with unloading.
Changes in the structural, biological, and mechanical characteristics with age
Ligament/tendon injuries are reported in all age groups, mostly ranging from adolescents to adults. 58 The regenerative process that the interface tissue undergoes may follow the same basic scheme as its natural development with aging. Therefore, an in-depth evaluation of the age-dependent changes at the interface is critical for guiding treatments for different populations.
The basic process behind the growth of interface tissue has been widely reported. Gao et al. 18 reported that the femoral enthesis of the rat MCL was formed early on by the ligament connecting to the cartilage. Then the enthesis was successively observed with an ossification of the cartilage and fibrocartilage developing in the ligament. In the midphase of the enthesis development, type I collagen was continuous from the ligament to the bone. The UFC zone was most pronounced in older rats where the type II collagen labeling was more than two times thicker than that in younger rats. Messner 20 reported a similar evolution of the rat cruciate ligaments, where it was also shown that the whole process did not happen simultaneously in the ACL and PCL, and not even synchronously at the two ends of the same ligament or within a single attachment. It was postulated that this asynchronous process may provide a less dramatic change in the mechanical environment at the entheses.
The morphological features of the collagen fibers, cells, and zone thicknesses at the interface have been reported to change with aging. Wang et al. 36 reported that in immature bovine ACL insertions, the collagen fibers in the ligament were relatively parallel to the ligament–cartilage interface, but in the mature group, the fibers inserted vertically to the interface. Kanazawa's 59 findings reported that collagen fibers became organized and the cell density decreased with aging, contradicting Wang's findings 23 that there was no significant difference in cell density with aging. Long et al. 60 reported an attenuated potential of cell proliferation and cell migration with aging. Yamada et al. 61 found that the number of cell processes decreased in the adult insertion, but the length of each process increased.
Wang et al. 23 reported a decreased thickness of the fibrocartilaginous zones with aging in bovine ACL insertions, while in Long's study, 60 area of the UFC was shown to decrease with aging, but the CFC area showed an increase with aging.
The contents of collagens and proteoglycans have also been shown to change with age. Wang et al. 23 reported that type X collagen increased with skeletal maturity. Gao et al. 18 found that the keratan sulfate was labeled at the MCL enthesis of younger rats but limited to the fibrocartilaginous zone in older animals. The dermatan sulfate was labeled after birth and continued to exist in older rats. Chondroitin 4 was labeled after birth but dissipated later, while the presence of chondroitin 6 was observed continuously. Wang et al. 23 reported that the ALP was positively stained in the interface region but the stain intensity decreased with age. Higher activity of ALP in the younger groups suggested a higher potential for mineralization and the ability to adapt to external loading.
Discussion and Future Directions
This review presents the current knowledge of the structure, biological contents, and mechanics of tendon/ligament–bone interfaces.
The basic features of tendon/ligament–bone interfaces have been shown to be highly specialized and vary from tissue to tissue and between the two ends of the same tissue. This probably reflects the unique mechanical environments and the corresponding demands placed on the enthesis. In the future studies, it is suggested to treat each interface as a special case to thoroughly understand the biological, morphological, and mechanical characteristics and to correlate these features with the unique in vivo loading environment. Specialized studies on each enthesis may facilitate a comprehensive understanding on the unique functional mechanisms and provide a scientific basis for relating tissue engineering work for improving clinical treatments.
Although it has been shown that the features of the ligament/tendon–bone interfaces have a unique three-dimensional spatial distribution,21,37,38,62 most previous studies only reported the measuring outcomes on a single plane/location, which may not represent the overall feature of the whole interface region. Failure to consider the surrounding mechanical environment and directional properties of the tissues may affect the main conclusions.
There are similarly very few studies on human subjects, and most available studies are based on elderly people. Since tendon/ligament ruptures are often caused by sports injuries, patients tend to be from a younger population. Therefore, it would be valuable to include adolescents or young adults in future studies to quantitatively characterize the interface features and thus to better elucidate the criteria for tissue engineering these organs for clinical use.
By far, most proposals on the mechanism of load transmission within the graded interface were concluded by comparing differences in the biological and structural features of different interface zones. These findings demonstrate that important biology–structure–function relationships may exist at the tendon/ligament-to-bone interface, which should be thoroughly considered when regenerating the interface in a repair or reconstruction surgery. However, there is still some debate on the structure–function mechanism of the interfaces, especially when mapping the spatial distribution of the zone thickness to its role in load bearing. There is no consensus about how the remodeling of different longitudinal zones correlates with each other, and whether the magnitude of normal tensile force or the shear tensile force, or the variation/uniformity of the force orientation, plays the dominant role in regulating the remodeling of the interface tissues.
Besides, high-level studies with evidence-based conclusions on the functional mechanisms are quite limited and require further investigation.
This review has several limitations. (1) The enrolled studies were restricted to English-language publications, although the authors did attempt to include related findings in other languages by reading the reports from the enrolled studies, who interpreted the main outcomes for the non-English publications. (2) Most of the enrolled studies have a low evidence level (e.g., laboratory observational study) and are conducted using different samples from different species and ages, which made it difficult to conduct a meta-analysis.
Summary
There are different types of ligament/tendon–bone interfaces according to the way the soft tissue connects to the bone, among which the fibrocartilaginous type is the most common. Previous studies have widely investigated the longitudinal features of the interfaces and found distinguished characterizations among different zones in terms of the biological content, microstructure, material properties, and blood supply. These features were shown to change with aging, loading, and tissue types. However, there are few studies on the transverse distribution of these features and the functional mechanisms behind it. Future studies should consider characterizing the features of each specialized tissue in younger human subjects. Further studies are needed to explore the functional mechanisms driving the differentiated features and provide evidence-based data for engineering specialized soft–hard tissue interfaces.
Footnotes
Acknowledgment
The authors would like to thank Mr. Colin McClean for his assistance with editing this article.
Authors' Contributions
H.W.: Conceptualization, methodology, formal analysis, writing—original draft, and writing—review and editing. K.H.: Formal analysis and writing—review and editing. C.-K.C.: Supervision, conceptualization, and writing—review and editing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. 32101050); the China Postdoctoral Science Foundation (Grant No. 2021 M702129); and the Fundamental Research Funds for the Central Universities (Project No. AF0820060).