
Abstract
The increasing number of elderly people across the globe has led to a rise in osteoporosis and bone fractures, significantly impacting the quality of life and posing substantial health and economic burdens. Despite the development of tissue-engineered bone constructs and stem cell-based therapies to address these challenges, their efficacy is compromised by inadequate vascularization and innervation during bone repair. Innervation plays a pivotal role in tissue regeneration, including bone repair, and various techniques have been developed to fabricate innervated bone scaffolds for clinical use. Incorporating neural-related cells and delivering neurotrophic factors are emerging strategies to accelerate bone regeneration through innervation. However, research into neurogenic cell sources remains limited. Meanwhile, neural stem/progenitor cells (NSPCs) are emerging as promising cells for treating neurodegenerative disorders and spinal cord injuries due to their multifunctional capacity in promoting angiogenesis, neurogenesis, and immunomodulation, making them promising candidates for achieving innervation in bone substitutes. In this review, we discuss the regenerative potential of NSPCs in tissue regeneration. We propose their feasibility for bone therapy through their secreted exosomes during traumatic brain injury, contributing to the acceleration of bone healing. Additionally, we discuss the essential neurotrophic factors released from NSPCs and their osteogenic properties. This review emphasizes the necessity for further investigation of the role of NSPCs in bone regeneration.
Impact Statement
Innervation has gained attention in bone tissue engineering, with nerve-related cells shown to promote bone regeneration. However, few previous reviews have examined the potential of neurogenic cells, particularly neural stem/progenitor cells, for bone repair. This review explores the feasibility of these cells in enhancing bone regeneration, discussing roles in angiogenesis, immune regulation, neurogenesis, and osteogenesis. The effects of these cells under traumatic brain injury and their neurotrophic factors on bone repair are also reviewed. Finally, the impact of secreted factors from neural progenitor cells on osteogenic differentiation is examined, highlighting their potential as a novel cell source for bone repair.
Introduction
Innervation is important for tissue regeneration and has recently been considered in the development of tissue-engineered organs. 1 Across various tissues, from skin to heart, the nervous system plays a significant role in modulating regenerative mechanisms. Sensory nerves detect skin tissue damage via calcitonin gene-related peptide (CGRP) mediating neuro-immune interactions (anti-inflammatory stimulation and M2 macrophage polarization). 2 Musculoskeletal tissue is also influenced by innervation. 3 Neurotrophic factors such as nerve growth factor (NGF), glial cell line-derived neurotrophic factor (GDNF), and ciliary neurotrophic factor have been shown to enhance skeletal muscle reconstruction and restore muscle function through axonal growth and angiogenesis improvement.4,5 The emerging role of neurogenic regulation in tissue regeneration has led to a growing interest in developing strategies to incorporate or promote innervation in tissue-repairing constructs.
In the context of bone tissue, the role of innervation in modulating bone remodeling and repair has gained much attention recently. Bone regeneration requires a balance between osteoclasts, responsible for bone resorption, and osteoblasts for bone formation. The nervous systems including sympathetic and sensory nerves essentially govern this complex bone remodeling. 6 Denervation can impair bone regeneration by decreasing the expression of collagen I, a major component of the extracellular matrix in bone. 7 Clinically, it has been reported that innervated bone grafts effectively prevented bone resorptions of the graft compared to the noninnervated ones, enhancing the successfulness of bone graft treatment.8,9 Recently, there has been a significant proposal highlighting the essential role of nerves as niche cells in promoting bone repair. 10 Furthermore, a previous study demonstrated that nerve fibers are involved in the formation of the stem cell niche in the bone marrow and the regulation of mesenchymal stem cell (MSC) homeostasis, which in turn controls bone remodeling. 11 In bone fractures, neuropeptides such as CGRP and substance P (SP) not only enhance the migration of bone MSCs and induce osteoblastic differentiation but also modulate osteoclastogenesis, reflecting a balance in the bone remodeling mechanism.12,13 Taken together, these results suggest the potential of the nervous system and its secreted molecules in bone tissue regeneration.
The innervation of tissue-engineered constructs can be obtained through several approaches, including conventional neurotization or host nerve transfer, preinnervation with neural cells (e.g., primary neurons and Schwann cells [SCs]), and biomolecule delivery such as NGF and brain-derived neurotrophic factor (BDNF).14–16 Although neurotization is the gold standard treatment, it is constrained by the limited availability of nerve grafts and the potential risk of neuroma formation. As a result, preinnervation with neural cells and biomolecule delivery are preferable and show promise for the future. However, the selection of appropriate cells and biomolecules remains a significant challenge for these strategies. Neural stem/progenitor cells (NSPCs) have emerged as promising candidates for tissue regeneration, particularly brain and spinal cord tissues, due to their inherent plasticity and multilineage differentiation potential. NSPCs are a type of cell in the nervous system that can self-renew and differentiate into various types of neural cells. According to their unique neurogenic features, NSPCs are considered promising candidates for the treatment of neurological diseases and may also serve as potential neurogenic sources for bone repair.
While numerous reports have been published on bone repair strategies and the role of nerves in bone regeneration,17–19 there is a lack of well-documented discussions regarding neurogenic cell sources and the feasibility of NSPCs as a potential treatment for bone injuries. In this review, we first discuss the roles of innervation in bone tissue regeneration, particularly focusing on paracrine signaling molecules. We then summarize the applications of nerve-related cells in bone tissue engineering and introduce NSPCs as potential neural cells for bone therapy. Following this, we explore the therapeutic roles of NSPCs in nerve and bone tissue regeneration, their potential in modulating bone regeneration observed in traumatic brain injury (TBI), as well as the osteogenic capability of their secreted growth factors. We propose that innervation is crucial for bone tissue regeneration, and although integrating innervation is challenging in tissue engineering, embedding neural cells or their secreted factors into tissue-engineered constructs or injury areas could substantially facilitate host innervation ingrowth and accelerate bone regeneration.
Roles of Neural Signaling Molecules in Bone Tissue Regeneration
The regulation of bone regeneration by nerves is largely influenced by neurotrophic factors/neurotransmitters and secreted molecules from neural supporting cells such as Schwann cells. 18 In injured bone tissue, there is an increased expression of CGRP, which arises from alternative splicing of the CALC gene. 20 This enhancement promotes the migration of bone marrow mesenchymal stem cells (BMSCs) to the fracture site and osteogenic differentiation in rats (Jia et al. 2019). Moreover, one study demonstrated that CGRP stimulates BMSC proliferation and osteogenic differentiation by enhancing Wnt/β‐catenin signaling in vitro, 21 highlighting the multiple roles of CGRP in the bone regeneration process. CGRP is often released alongside another major neurotransmitter, SP, which has also been proven to promote the osteoblastic differentiation of BMSCs and MC3T3-E1 cells by activating the Wnt/β-catenin signaling pathway. 22 These interconnected roles of CGRP and SP highlight their combined contribution to bone regeneration.
Apart from those two neurotransmitters mentioned, norepinephrine (NE) and acetylcholine (ACh), also draw attention to bone regeneration. NE, a neurotransmitter released by noradrenergic nerves, influences bone regeneration through α‐adrenergic and β‐adrenergic receptors in various bone-lineage cells. It inhibits osteoblast activity and hBMSC proliferation while inducing osteoclast formation by activating RANKL/OPG.23–25 ACh has also been reported to promote osteoblastic proliferation with minimal impact on differentiation. 26
Furthermore, neuropeptide Y (NPY), NGF, BDNF, and neurotrophin-3 (NT-3) also facilitate bone regeneration as shown in several studies. Liu et al. demonstrated that NPY stimulates osteogenic differentiation and mineralization of BMSCs. 27 NGF has been shown to promote osteoblastic differentiation of mouse embryonic osteogenic precursor cell line MC3T3-E1 via bone morphogenic protein-2 (BMP-2) signaling. 28 BDNF was found to modulate osteogenic differentiation of mouse osteoblast precursor cell lines MC3T3-E1 via TrkB receptor and JNK and p38 MAPK signal pathways. 29 NT-3 was also shown in vitro to promote osteogenesis in rat bone marrow stromal cells. 30
Many of these neurotrophic factors are produced by NSPCs, as summarized in Table 1, suggesting the potential of NSPCs to improve bone formation.
Neurotrophic Factors Secreted by NSPCs with Osteogenic Properties
BMSCs, bone marrow mesenchymal stem cells; NPCs, neural progenitor cells; NSCs, neural stem cells; NSPCs, neural stem/progenitor cells; OCN, osteocalcin; TGF-β, transforming growth factor beta.
Nerve-Related Cells for Bone Tissue Engineering
Both neural paracrine signaling and nerve-related cells can be advantageous for bone repair. Previous studies have explored the potential of nerves themselves and various nerve-related cells, such as Schwann cells (SCs) and neural crest cells (NCCs), in repairing bone injuries (Fig. 1). These findings are summarized in Table 2. Although these cells have shown favorable outcomes, there is still room to explore alternative neurogenic cell types in bone tissue engineering.

Potential of neural crest cells (NCCs) and their derived cells in bone repair. NCCs are multipotent stem cells located at the side of the neural tube near the epidermis after neurulation. They can develop into many different cell types, such as neurons, Schwann cells (SCs), mesenchymal stem cells, and osteoblasts. NCC-derived mesenchymal cells and Schwann cells have been widely demonstrated to improve bone regeneration. Created with BioRender.com.
Application of Nerve-Related Cells for Bone Repair
TGF-β, transforming growth factor beta.
Schwann cells
SCs are glial cells residing in and supporting peripheral nerves. They can produce myelin sheath, a fatty substance, wrapping around nerve fibers for efficient electrical impulse transmission. They are widely used to improve peripheral nerve regeneration, thus suggesting the possibility of utilizing Schwann cells to promote innervation for bone defect repair. A recent study reported that β-tricalcium phosphate scaffolds loaded with rat SCs and BMSC-derived endothelial cells enhanced new bone formation in rat femoral bone defect models. 51 This was from the angiogenic potential of SCs, which increases the nestin and vascular endothelial growth factor (VEGF) expression of endothelial cells. Additionally, exosomes derived from rat SCs promoted proliferation, migration, and osteogenic differentiation of BMSC as well as bone repair in vivo. 52
Neural crest cells
NCCs are multipotent, migratory cells that originate along the border of neural plates and play a crucial role in vertebrate development. They give rise to a diverse array of cell types and structures, including elements of the peripheral nervous system and craniofacial cartilage and bone. In the context of bone repair, NCCs contribute significantly by differentiating into osteoblasts, the cells responsible for new bone formation. 62 A previous study showed that cranial NCCs promoted bone formation after transplantation in a mouse calvaria defect. 59 NCCs also exhibit MSC characteristics of differentiation into mesenchymal cell lineages such as chondrocytes, osteocytes, and adipocytes. Glaeser et al. differentiated the induced pluripotent stem cells (iPSCs)-derived NCC into mesenchymal progenitor cells and showed that these cells increased the bone volume of the mouse cranial defect model. 58
Source of NSPCs
Neural stem cells (NSCs) play an important role in forming the central nervous system (CNS) during development. They differentiate into radial glial cells and neural progenitor cells (NPCs). In adulthood, NSPCs remain the only self-renewing cell type in the CNS, essential for neurogenesis and gliagenesis. These cells are considered multipotent, meaning they can give rise to neurons, astrocytes, and oligodendrocytes, the main cell types in the nervous system. During embryonic development, NSPCs are found in the neural crest, where their resident cells (NCCs) support skeletal tissue and bone regeneration, and the cortex.
NSPCs can originate from various species and multiple origins, such as embryonic stem cells (ESCs), iPSCs, adult CNS, and the direct conversion of nonneural cells. Nevertheless, the prevalent method for obtaining and examining neurons remains to isolate primary NSPCs directly from fetal tissue. Neurogenic stem cells have been suggested to share similar characteristics with osteogenic stem cells, including their association with blood vessels, expression of nonspecific markers, and involvement in molecular signaling pathways such as Wnt, BMP, and Notch. 63 Despite their shared potential, BMSCs have received more focus in bone repair due to their well-established osteogenic properties. However, the potential of neurogenic stem cells for enhanced bone regeneration has been increasingly recognized in recent times. This area holds great promise for future exploration.
Advances in neural stem cell biology and the promising results from transplantation studies in animal disease models have established NSPCs as a leading candidate for the advancement of clinically viable exogenous stem cell therapies.64,65 Their secretomes have also demonstrated a great potential to enhance angiogenesis and neurogenesis in the mouse spinal cord injury model and Alzheimer’s disease mouse model, respectively.66,67 Not only NSPCs can repair CNS, but they are also widely examined for the repair of the peripheral nervous system, which largely innervates bone tissues. 68 These regenerative capabilities of NSPCs may efficiently promote nerve regrowth and angiogenesis within bone defects, thereby supporting bone repair. This underscores the significance of further research to explore novel approaches in bone regenerative therapies.
Therapeutic Potential of NSPCs
In this section, we discussed the regenerative signaling molecules of NSPCs in promoting angiogenesis, neurogenesis, and modulating the immune system based on previous literature (Table 3). We further elaborated on these signaling molecules that enhance osteogenesis, suggesting the potential of NSPCs in bone repair. Importantly, we summarized previous studies showing the capability of NSPCs to promote bone regeneration. The reported signaling molecules are summarized in Figure 2.
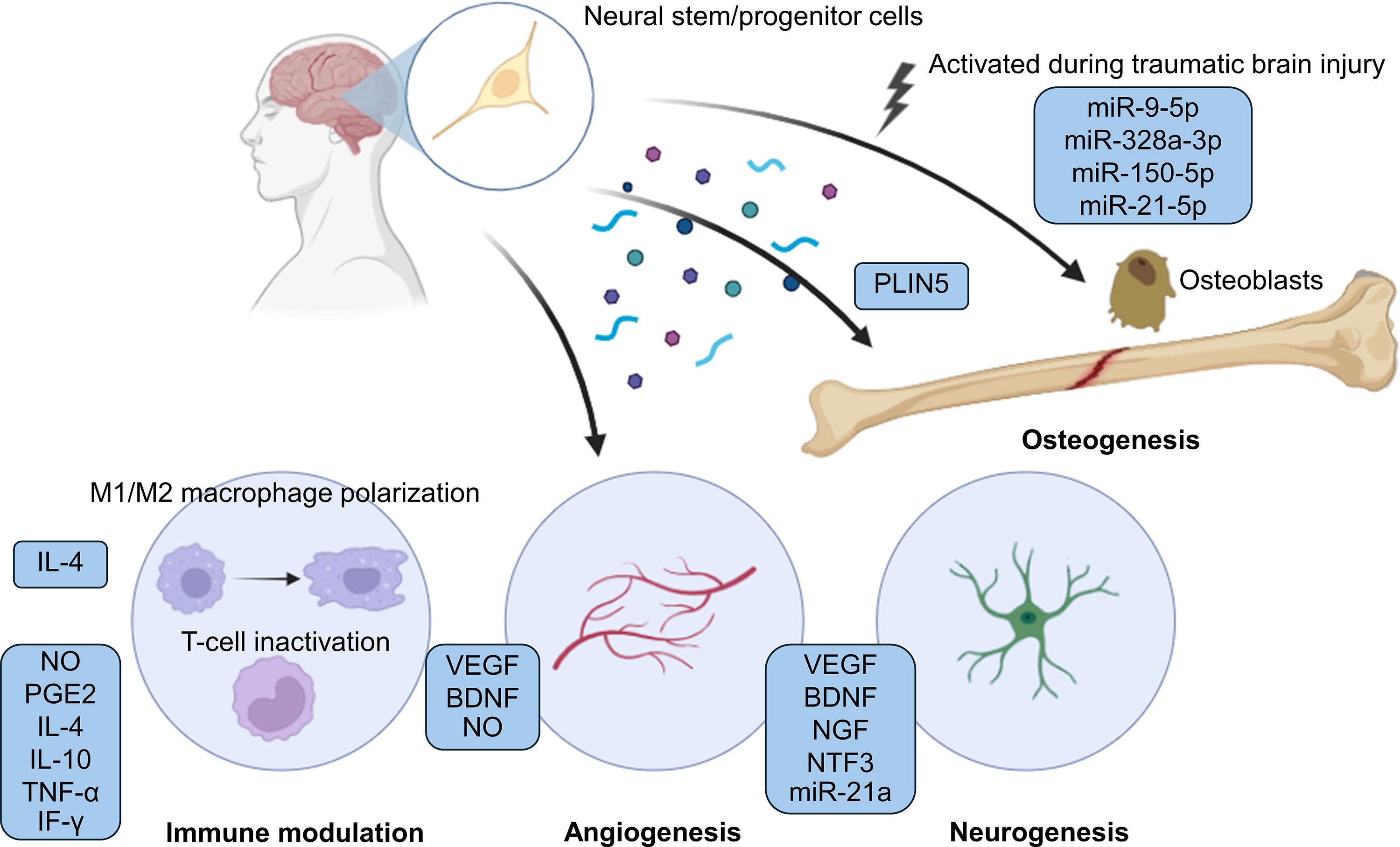
Illustration showing how NSPCs regulate bone regeneration, immune modulation, angiogenesis, and neurogenesis through secreted factors. During traumatic brain injury, NSPCs produce miRNA-enriched extracellular vesicles that promote osteogenesis in osteoblasts and enhance bone regeneration. Under normal conditions, these cells also enhance bone regeneration via exosomal PLIN5. Besides their osteogenic potential, NSPCs exhibit various regenerative abilities through secreted factors that support immune modulation, angiogenesis, and neurogenesis, all of which may potentially support bone repair. BDNF, brain-derived neurotrophic factors; IF-γ, interferon-gamma; IL-4, interleukin-4; IL-10, interleukin-10; NGF, nerve growth factor; NO, nitric oxide; NSPCs, neural stem/progenitor cells; PGE2, prostaglandin E2; PLIN5, perilipin 5; TNF-α, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor. Created with BioRender.com.
The Multifaceted Roles of NSPC and Its Secretome in Nerve and Bone Repair
CGRP, calcitonin gene-related peptide; HIF-1α, hypoxia-inducible factor 1-alpha; HUVEC, human umbilical vein endothelial cell; NO, nitric oxide; OPN, osteopontin; TGF-β, transforming growth factor beta.
NSPCs promote angiogenesis in damaged tissues
The observation of the angiogenesis role of NSPCs across in vitro and in vivo models suggested the possible benefits of their therapeutic applications. These include the promotion of revascularization and the repairing processes after injuries. NSPCs isolated from embryonic mice promoted microvascular regeneration and protected endothelial cells under the ischemia condition through a controlled in vitro oxygen and glucose deprivation mouse brain model via the hypoxia-inducible factor 1-alpha (HIF-1α) and VEGF signaling pathways. 69 Lee et al. 72 also provided evidence of the relationship between VEGF levels and angiogenesis. In that study, NSCs, which are isolated from the culture of the human fetus telencephalon tissue, were engineered to overexpress the VEGF levels, and the result showed that the overexpression has increased angiogenesis of the grafted NSCs compared to the control in a mouse intracerebral hemorrhage model. 72 All these findings contribute to a deeper understanding of the mechanisms underlying the angiogenesis capability of NSPCs, which may also play a significant role in the processes associated with bone regeneration.
HIF-1α is a signaling molecule that provides the essential pathway for bone development. A previous in vivo study using the mouse distraction osteogenesis model of skeletal repair highlighted the importance of the HIF-1α pathway in promoting angiogenesis for bone regeneration. 85 The study found that the HIF-1α pathway is essential for both the angiogenic and osteogenic aspects of bone healing. Furthermore, Chen et al. demonstrated that HIF-1α directly promoted bone resorption in fractures and improved osteogenesis via cardiotrophin-1 during bone healing. 86 Kleinheinz et al. (2005) investigated the impact of recombinant human VEGF (rhVEGF165) on forming new blood vessels and bone tissue in a mandibular defect model. They found that activating angiogenesis with rhVEGF165 increased blood vessel formation and enhanced bone regeneration. 87 As a result, soluble factors released from NSPCs may positively affect bone regeneration both through angiogenesis and a direct link to the osteogenic pathway.
NSPCs modulate immune response of damaged tissues
NSPCs are essential for regulating immune responses in damaged tissues. These involved the balancing between proinflammatory and anti-inflammatory factors within the damaged area. Wang et al. found that NSCs isolated from the CNS of developing adult mice appeared to suppress mouse T cells through nitric oxide and prostaglandin E2 (PGE2) production. 74 Kim also showed that secreted factors from human NSCs such as interleukin-4 (IL-4), IL-10, and tumor necrosis factor-alpha can suppress the proliferation of activated T cells in an in vitro culture model. 76
Bone regeneration is crucially influenced by immune response because of their common cytokines and signaling molecules. T cells, B cells, and macrophages play an important role in regulating bone regeneration. The modulation of these immune cells using anti-inflammatory cytokines like IL-4 and IL-13 has demonstrated the capacity to enhance bone healing when administered locally. 88 Nonetheless, the efficacy of these effects is dependent upon the timing and mode of delivery. Previous studies suggested that COX-2/PGE2 may mediate osteoinductive communication between inflammatory macrophages and BMSCs, contributing to bone regeneration. 89 Additionally, PGE2 is not only expressed in immune cells but also in bone cells like osteoblasts and osteoclasts, 90 suggesting that targeting PGE2 may be another promising approach to treating bone injury. A previous study reported that PGE2 promoted osteoblast proliferation and differentiation through the activation of sensory nerves, which are innervated in bone tissues. 24 To sum up, NSPCs could serve as valuable agents in modulating the inflammatory environment to promote bone repair and regeneration.
NSPCs improve neurogenesis and nerve regeneration
NSPCs play a significant role in neurogenesis and nerve regeneration. This was evidenced by an improvement of motor function recovery from transplanted NPCs in a rat spinal cord injury model (Aceves et al., 2023), along with the evidence from a clinical trial of the transplantation of human NSCs, isolated from fetal brain tissues, in patients with progressive multiple sclerosis that promoted levels of cerebrospinal fluid proteins and a neuroprotective remodeling of the CNS microenvironment (Genchi et al., 2023). In Onode’s study, the bioabsorbable polymer nerve conduit that was coated three-dimensionally with hiPSC-derived neurospheres could promote functional recovery and axon regeneration in rat sciatic nerve injury models, suggesting the great potential of hiPSC-derived neurospheres. 79 In a study by Lee et al., the NSCs derived from aborted human fetal telencephalon improved numerous aspects of Alzheimer’s disease and restored memory in mice. 35
Previous studies reveal a close relationship between the nervous and bone systems. Therefore, the nerve regeneration capability of NSPCs, either directly from cells or indirectly from their secreted factors, can potentially enhance bone regeneration processes. Additionally, NSCs can be differentiated into SC-like cells, 91 highlighting their potential in bone repair similar to the reported ability of SCs to improve bone regeneration.51,54 Their released neurotrophic factors, which could contribute to osteogenic regeneration, are summarized in Table 1.
NSPCs improve osteogenesis in bone
Currently, a direct effect of NSPCs on bone regeneration has been evidenced. Zhang et al. showed that primary rat NSC-loaded GelMA–Li–Ca–Si hybrid bioinks significantly enhanced osteogenesis-related proteins, such as bone sialoprotein and osteocalcin, in BMSCs. 82 This approach resulted in the highest bone formation in a rat cranial bone defect model compared to the acellular group. Additionally, exosomes derived from primary NSCs have been demonstrated to promote osteogenic differentiation of BMSCs in vitro and improve new bone formation through both exosomal PLIN5 and miR-9-5p.83,84
NSPC-Derived Exosomes Enhance Bone Fracture Healing Under TBI
Apart from the multifaceted regenerative potential of NSPCs, the activation state of these cells has been evidenced to enhance bone regeneration both in vitro and in vivo in a scenario like traumatic injury.
The interaction between the brain and bone has largely gained attention in the field of bone tissue engineering. One example can be seen during an exercise that involves mechanical stimulation sending signals to the brain via the nervous system. These signals are transmitted to the brain, which allows communication back to the skeletal system and, thereby, bone tissues. Not only the electrical signals but also the chemical signals from the brain that profoundly influence bone growth and repair. Interestingly, TBI or craniocerebral injury has been associated with accelerated bone fracture healing in several clinical settings.92–94 This regenerative effect was governed by the released factors from the brain transporting through the blood to the bone tissues. It has been shown that extracellular vesicles (EVs) released from the brain circulate to the bone injury sites in vivo. Importantly, exosomes derived from NSPCs, which were isolated from TBI rats, improved osteogenic differentiation in vitro via miR-9-5p. 83 This study demonstrated the potential of secreted factors such as exosomes from NSPCs to repair bone regeneration both in vitro and in vivo, prompting further studies to investigate the role of NSPCs in bone repair. Exosomes in blood plasma improve osteogenesis, 92 suggesting that activated cells within the brain under TBI may affect the exosome profiles in the blood plasma. Xia et al. also showed that injured neurons in the hippocampus released EVs that are transported through the circulatory system to promote bone regeneration. 93 They reported that miR-328a-3p and miR-150-5p in the EVs are responsible for this effect targeting osteoprogenitor cells. Additionally, miR-21-5p in the EVs from mouse serum enhanced mouse femoral fracture healing. 95 These suggested that miRNA-enriched EVs or exosomes from injured neurons or NSPCs in the brain can target bone regeneration. However, there are limited studies examining the role of NSPCs in bone regeneration during normal conditions.
Several reported miRNAs from NSCs have been shown to regulate osteogenic differentiation. For example, miRNA-124, a key player in NSC proliferation, 96 has been associated with the healing of metaphyseal fractures of the distal tibia in patients. 97 MiR-219a is involved in the regulation of NSC during differentiation and has demonstrated osteogenic potential to modulate the retinoic acid receptor‐related orphan receptor beta during osteoblast differentiation. 98 Based on the data on miRNA from NSPCs, these cells could potentially exert positive outcomes on bone remodeling.
Conclusion and Future Perspectives
Bone regeneration is a complex mechanism involving intricate interplays of not only osteoblastogenesis and osteoclastogenesis but also angiogenesis, immune modulation, and innervation. Recent research emphasizes the importance of the nervous system in supporting bone formation and preventing graft resorption. Despite advancements in bone repair treatments, the role of neurogenic factors has often been overlooked, limiting regenerative efficacy. While previous studies have explored the potential of neurotrophic factor delivery in bone repair, there remains a gap in investigating the role of neural-related cells in bone repair. Current research has predominantly examined SCs and NCCs, highlighting the need of seeking alternative cell sources for bone therapy. NSPCs represent promising neurogenic cell sources, having been utilized in treating neurological diseases and spinal cord injuries. Our review elucidates the significant regenerative properties of NSPCs, including the stimulation of osteogenesis, angiogenesis, immune modulation, and nerve regeneration. In recent years, researchers have gained interest in investigating the regenerative properties of these cells for bone repair, demonstrating promising outcomes. Of particular interest is the observation that NSPC-derived exosomes could accelerate bone regeneration under TBI conditions. Furthermore, NSPC-loaded constructs have been shown to improve osteogenesis both in vitro and in vivo. By deepening our understanding of NSPCs and their secreted biomolecules’ effect on bone regeneration, more effective therapies to improve bone fracture healing can be achieved.
Footnotes
Authors’ Contributions
P.M.: Conceptualization, funding acquisition, methodology, validation, formal analysis, investigation, writing—original draft, and writing—review and editing. P.Y.: Conceptualization, methodology, validation, formal analysis, investigation, and writing—original draft. W.S.: Conceptualization, supervision, and writing—review and editing.
Disclosure Statement
The authors have no potential conflicts of interest to declare.
Funding Information
This project is funded by the National Research Council of Thailand (NRCT), contract number N42A670586.