
Abstract
Degenerative disc disease (DDD) is the main cause of back pain, one of the most impactful musculoskeletal disorders. In the last decades, significant advances have emerged in the field of intervertebral disc (IVD) biology. Despite cumulative knowledge on cell heterogeneity, there is a lack of understanding about the molecular signature of different IVD cell populations and their potential as therapeutic targets. This is not only due to progress in single-cell analysis being recent, but also to difficulties and lack of standardization in the isolation of distinct IVD cell populations based on the phenotypic markers used. Herein, we review the literature on IVD resident cells and the main challenges faced in the source, isolation, and culture of IVD cell subsets that are yet to be overcome. The profiling of IVD subpopulations and their cellular dynamics with development, aging, and disease progression will certainly advance the knowledge on IVD homeostasis and associated diseases, while contributing to the development of game-changing, personalized targeted therapies.
Impact Statement
Degenerative disc disease (DDD) is a major cause of back pain, with significant societal impact. While advances in intervertebral disc (IVD) biology have revealed cellular heterogeneity, understanding molecular signatures, and therapeutic potential of IVD cell populations remains limited. Addressing challenges in isolating and profiling these cells could transform IVD pathophysiology, enabling the development of personalized, targeted therapies to revolutionize DDD treatment.
Intervertebral Disc Cellular and Molecular Dynamics: From Embryonic Development to Tissue Degeneration
The intervertebral disc (IVD) originates from both notochord and mesenchyme regions 1 through a precise regulatory mechanism, involving several genes and signaling pathways. Indeed, it is the origin from distinct germ layers during embryonic development that is behind the IVD’s cell heterogeneity. The notochord is a rod-like mesoderm-derived structure containing large vacuolated cells surrounded by a basement membrane rich in collagens, laminin, and proteoglycans—the notochordal cells (NCs). 2 The human notochord first appears around the 17–19 days of gestation, until reaching its definitive form around 26–30 days. This structure plays a crucial role during embryogenic processes 3 and orchestrates the formation of several surrounding tissues (neural tube, sclerotome, pancreas, aorta) through the release of growth factors and morphogens.4,5 Moreover, this structure is involved in the direct formation of the nucleus pulposus (NP), the core of the IVD, and in the regulation of cell migration, differentiation, and survival.6,7 The notochord structure attracts sclerotome cells that migrate from the somites (around day 30 of fetal gestation) and concentrate around the notochord, forming a more condensed zone, that will then segment, giving rise to segmented cartilage, and in particular to the annulus fibrosus (AF), the tissue surrounding the outer region of the IVD, and to a less compact zone, from where the vertebral bodies originate. 8 Simultaneously, the notochord contracts through the vertebral bodies and expands within the other IVD regions, giving rise to the NP, being regulated by distinct signaling pathways (e.g., Shh, Wnt, noggin, Pax) 8 (Fig. 1). Importantly, the notochord structure disappears during IVD formation, thus in mammals well before birth.
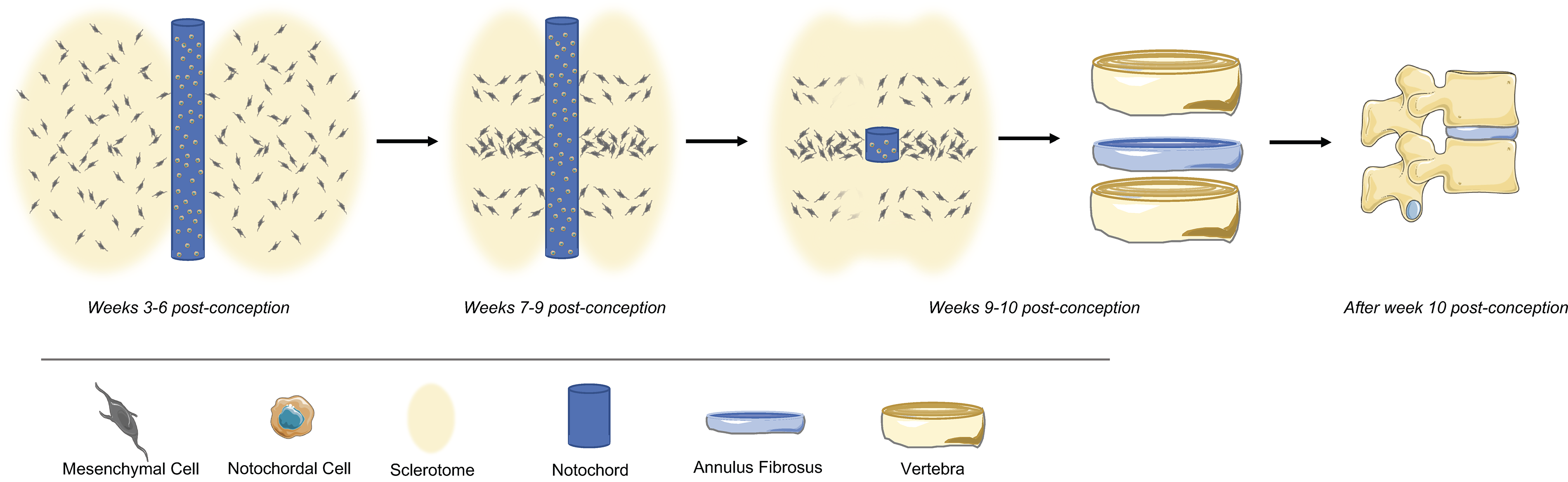
Schematic representation of the intervertebral disc organogenesis (figure created in Biorender).
With increasing age, degeneration of the IVD naturally occurs, being mainly recognized by the characteristic steep loss of native extracellular matrix (ECM) and disc integrity. Aging is accompanied by changes in IVD cell phenotype, an imbalance of proinflammatory markers and metalloproteinases (MMPs), cell senescence, and altered metabolism. 9 These alterations are, in part, linked with limitations of the nutritional intake caused by calcification of the cartilaginous endplates (CEPs). 10 This blockade in cell nutrition also leads to the accumulation of cell waste, thus creating increasingly acidic conditions in the degenerated IVD. 11 Under acidic conditions, MMPs production accelerates, thus contributing to the loss of tissue integrity, accompanied by a significant loss in an ECM rich in proteoglycans. 12 Overall, the intense production of inflammatory molecules greatly contributes to the loss of disc integrity which, ultimately, exposes the immune-privileged inner disc to the systemic immune system, and enabling the diffusion of neurogenic and angiogenic factors.
Although several authors have clearly demonstrated that the IVD is populated by a heterogeneous mixture of cells whose variability extends to several levels of specificity, its translation into well-established cell phenotypes has yet to be achieved. Nevertheless, many of these cell subsets have not yet been isolated and so, their contribution to IVD degeneration has yet to be addressed (Fig. 2). The isolation and culture of IVD cells, either from NP or AF tissues, has been performed for decades, mostly based on macroscopic tissue separation, dissection and enzymatic digestion. Most literature relies on the study of NP or AF cells as distinct homogeneous cell populations from two neighboring IVD tissues, although their heterogeneity has been documented for decades. Furthermore, in the last years, single-cell transcriptomics has contributed to the identification of several IVD cell subsets, including NCs, mesenchymal-like cells, NP and AF progenitor-like cells, as well as myeloid/immune-like cells.13,14

Overview of the IVD cell populations and their respective markers (NC = notochordal cell; NP progenitor = nucleus pulposus progenitor; NP cell = nucleus pulposus cell; AF cell = anulus fibrosus cell; CEP = cartilage endplate cell). The figure illustrates the heterogeneity of IVD cells obtained from distinct cell sources (human, bovine, pig, rodents) and upon tissue digestion with enzymes (collagenases, pronase, trypsin, hyaluronidase) and the remaining challenges (illustrated by “?”) in: (1) the culture parameters for each of these populations (hypoxia, glucose level, cytokine supplementation, ions, 2D vs. 3S microenvironments [spheroids, hydrogels]); and (2) to address their specific contribution for IVD disorders (figure created in Biorender).
The IVD Cell Populations: Perspectives for Tissue Engineering
The study of IVD cell populations plays a crucial role in advancing the field of tissue engineering, particularly in developing novel strategies for treating degenerative disc diseases (DDD). 15 Understanding the biology, mechanotransduction, and interactions of these cells with each other and with other systems (e.g., nervous, hematopoietic) is essential for designing effective regenerative therapies for IVD.
DDD is a major cause of chronic back pain and disability, often resulting from the loss of IVD cells and degradation of the ECM. 16 Tissue engineering offers a promising solution to restore disc structure and function.17,18 Studying IVD cells enables the identification of optimal cell sources for regenerative approaches. Furthermore, advances in bioengineering techniques, including 3D bioprinting, scaffold development, and bioreactors, rely on a comprehensive understanding of IVD cell behavior under pathophysiological conditions.19–21 By mimicking the native microenvironment of the IVD, tissue-engineered constructs can be designed to enhance IVD cell survival and matrix synthesis, while reducing the production of inflammatory mediators and neurotrophic factors, ultimately improving the efficacy of engineered IVD implants and reducing the associated back pain. 22 Integrating the research in IVD cell subsets with tissue engineering also facilitates the development of biomimetic hydrogels, injectable cell-laden scaffolds, and gene therapy strategies to modulate inflammation and promote disc repair. Moreover, insights into IVD cell senescence, apoptosis, and metabolic activity aid in refining therapeutic approaches to slow or reverse degenerative processes.23,24 Overall, the study of IVD cell populations is essential to the progression of tissue engineering applications, offering a pathway toward innovative, biologically inspired treatments for spinal disorders. By bridging IVD cell biology with engineering principles, researchers can develop more effective and sustainable solutions for IVD regeneration and spinal health.
In the next sections, the phenotypic signature of the abovementioned IVD cell populations will be reviewed.
IVD Notochordal Cells
A subpopulation of large vacuolated notochordal-like cells (NC) has been consistently described in the NP of the IVD. It is believed that NC cells start to dramatically decrease after birth, until their disappearance, being replaced by chondrocyte-like cells. 25 This has been proposed to be caused by an increase in mechanical forces after birth, which could induce NC differentiation and/or apoptosis. 8 In early infancy/adolescence, the IVD environment becomes harsh due to the decrease of oxygen and nutrient supply, limiting cell survival. These environmental changes may induce the disappearance of NCs and the appearance of mature NP cells, more able to survive in low nutrient and oxygen supply than NCs. 8 However, Richardson and colleagues have described a population of NCs in both young and adult human NP coexpressing Brachyury (T), Galectin-3 (Gal3), carboxy-anidrase 12 (CA12), and CD24 using gene expression, immunohistochemistry, and flow cytometry, thus proposing a signature for NC: CA12+Gal3+T+CD24+. The expression of these markers appears to be independent of age and IVD degeneration level, suggesting that a small portion of NP cells may retain a NC-like phenotype. 26 In another study, Liu and colleagues have also demonstrated the existence of CD24+ cells in human NP tissue, but concomitant with a dramatic reduction of CD24+ cells with increased severity of IVD degeneration. 27
Contrarily to humans, other species (e.g., mouse, rat, cat, mink, dog, pig, and rabbit) are able to maintain a population of NC cells throughout their lives. 28 Porcine discs are believed to hold NC throughout adulthood, while bovine discs, more similar to humans, tend to lose the typical physaliferous phenotype early during development, 28 which could justify this difference in the expression of notochordal markers. It is described that 6-month pig coccygeal IVDs express cytokeratin-8 (KRT8+) in almost all the cells, while only 10% of cells from adult coccygeal bovine discs express this NC marker. 29 KRT8+ bovine cells were frequently found in clusters in the central part of the disc, a common NC phenotype. 29 Moreover, its expression was found to be specific to NP cells. In another study, leptin receptor (LepR) was described as a NC marker, being expressed by around 75% of mice NP cells during adulthood. 30 All LepR+ cells also express KRT8, yet not all KRT8+ cells were LepR+, which anticipates a further subdivision within the KRT8+ population. Importantly, these cells (LepR+) were colocalized with most of the aggrecan (ACAN) staining of the tissue, while LepR− cells expressed mostly MMP3, suggesting different roles in the IVD ECM metabolism. This work also confirmed a population of 75% LepR+ cells in human IVD, that increased with degeneration, possibly as a tentative response from the IVD to counter the degenerative process, although this mechanism remains to be dissected.
More recently, single-cell transcriptomics data have allowed the identification of multiple IVD cell populations, including a small but clear cluster of NCs in both human and bovine IVD tissue. These clusters were frequently associated with the typical NC-like markers TBXT/Brachyury, CD24, GAL3, KRT8, and NOG. Despite sharing some gene expression patterns and spatial localization, Panebianco et al. found that the NC cluster could be specifically distinguished from NP subsets by an upregulation of the genes KRT8, ATP6V1G3, C1QTNF3, CD55, and SPP1.13,31,32
Overall, the presence of NC-like cells in the adult NP holds great potential for clinical applications. NC cells may constitute a suitable cell source for regenerative therapies. Nevertheless, the ideal substrate and cytokine cocktail for their in vitro expansion is still to be defined. Once it has been identified, it will certainly enable the reestablishment of a healthy and functional IVD.
IVD Progenitor (Non-Notochordal) Cells
IVD cells with progenitor potential, but not with notochordal origin, have been described. These include a subset of IVD cells with a mesenchymal-like phenotype, as well as clonal ability. 33 These cells were shown to express mesenchymal markers (CD90, CD73, CD105, CD166, and CD106dim) and exclude the expression of hematopoietic markers (CD19, CD34, CD45, and HLA-DR), resembling the profile of adult progenitors’ mesenchymal stem/stromal cells (MSC).34,35 Functional assays demonstrated that these IVD cells can differentiate into chondrogenic and osteogenic lineages, while adipogenic differentiation was not always observed, with their morphology (round in endplate [EP], polygonal in NP, and elongated in AF cells) not always matching the traditional spindle-shaped morphology of MSC.35–37
In another perspective, Sakai and colleagues have identified a population of NP progenitor cells expressing Tie2 (Angiopoietin-1 receptor) and GD2 (disialoganglioside) antigens in both human and mice. 38 Tie2+ and GD2+ NP cells were able to form spheroid-like colonies and produced COL2 and ACAN. These cells were shown to differentiate into mesenchymal lineages, after injection in injured IVDs of nonobese diabetic/severe combined immunodeficient (NOD/SCID) mice, inducing NP tissue reorganization. In addition, the authors have detected that Tie2+ cells decrease with aging, particularly after 25 years old, 39 also correlating with the severity of IVD degeneration. These data suggested an association between NP progenitor cells disappearance and IVD tissue modifications. 38
In another work, bovine NP Tie2+ cells were shown to be approximately 8% of the whole population, 40 while in the work of Molinos et al., a much higher percentage of Tie2+ cells in the NP was reported (around 80%). Despite the fact that both studies analyzed bovine coccygeal IVDs around 1 year old, two different antibodies were used (antirat/CD202b polyclonal antibody, clone bs-1300R) 40 versus antihuman monoclonal antibody, clone 83715. 41 distinct cell isolation protocols (pronase vs. collagenase), and probably different cow breeds, which might explain the discrepancy between Tie2 expression levels in the NP. Additionally, these results also point out the problems of the reliability of antibovine antibodies.
This cell population (Tie2+ cells) also showed differentiation potential into the osteogenic, chondrogenic, and adipogenic lineages, contrary to Tie2−, although both subsets were able to form colonies, indicating the loss of Tie2 expression as a marker of loss of potency capacity and therefore more committed populations of progenitor cells. Regarding cell morphology, Tie2− cells maintained a spread, plastic adherent fibroblastic morphology, while Tie2+ cells appeared to be round and tended to form spheroids, a known hallmark of less differentiated cells. Tie2+ NP cells decrease rapidly in humans, 39 bovines, and represent 0.1% of the entire IVD cell population in adult dogs. Regarding small animals, as mice, the hierarchy for a population of NP progenitor cells was unveiled with coexpression of Tie2, GD2, and CD24. 38 The authors found that Tie2+GD2− cells were able to differentiate into Tie2+GD2+ cells in culture. These cells, initially CD24−, were shown to further differentiate naturally into Tie2+GD2+CD24+ cells with multilineage differentiation potential differentiated at least in two (of three) classical mesenchymal lineages. Interestingly, both populations were positive for the mesenchymal markers CD44, CD49f, CD56, CD73, CD90, CD105, and CD166 and negative for CD45, CD14, CD11c, CD31, CD34, CD144, CD146, and von Willebrand factor, further validating the classical phenotype of mesenchymal progenitor cells. 34
Henriksson et al. have also described the presence of IVD progenitor cells expressing BrdU (proliferating marker) in the border area between NP and AF, suggesting that these cells have a progenitor capacity (and express stem cell-related markers as Notch1, Delta4, Jagged1, C-KIT, KI67, and Stro-1) in an IVD niche. Interestingly, this study was conducted in four species (rabbit, rat, minipigs, and human). 42 In another work, Tan and colleagues analyzed the expression pattern of TAGLN (a cytoskeletal protein reported to regulate the differentiation of human skeletal stem cells, downregulated in aged and degenerated human NP) in TAGLNLacZ reporter mice. The results obtained also indicated a niche of proliferative cells in the NP of young mice, enriched for TGF-β signaling, suggesting that TAGLN+ are potential progenitors crucial for NP homeostasis. 43
More recently, single-cell transcriptomics analysis has also identified specific clusters of NP cells that possess a progenitor cell phenotype. 31 Gan et al. defined multiple NP progenitor cells (NPPCs) populations based on the expression of mesenchymal cell-associated markers such as PDGFRA, PRRX1, and IGF1. The authors additionally found NPPCs subcluster with the ability for colony formation as well as osteogenic and chondrogenic differentiation, similar to MSCs, which they categorized as resident progenitor cells in the human IVD. 13 Most of the identified NPPCs subsets were also translated to rat IVD. 44 Moreover, Calió et al. found a progenitor stem cell cluster in bovine NP that expressed stem cells, but not NC, markers (CD44, CD63, CD55). 32 These works have also suggested that the IVD progenitor cells subsets play an intrinsic role in regulating both NC and NP cells, thereby contributing to IVD homeostasis.
Overall, the discovery of a subpopulation of progenitor cells in the IVD has an immense significance for understanding disc cell biology, as well as the mechanism behind tissue degeneration and possible therapeutic targets, since their decline is strongly correlated to this pathological condition.
NP Cells
NP cells are nowadays recognized as a heterogenous population in the IVD. And while most of the scientific community believes that NP cells directly derive from the embryonic notochord, 45 others have proposed that they might be recruited from adjacent tissues (e.g., EPs).8,46 This cell heterogeneity has been documented already in 1995, when Chelberg and colleagues reported the existence of about 20% of cells in the IVD lacking matrix production profiles aligned with either NP or AF cells, suggesting the presence of more heterogeneous populations in this organ. 47 Besides the inherent diversity linked to these different origins, cellular changes within the NP tissue have also been associated with alterations in the nutritional and mechanical microenvironment experienced during IVD development and aging. 8 These shifts encompass, for example, those that occur during childhood when IVD cell density dramatically decreases. 48 By analyzing IVD specimens isolated from autopsies of individuals ranging from 0 to 86 years old, Boos’ group showed that cell density declines in the NP, AF and CEP between 0 and 16 years old, and in particular from birth up to 3 years old. 49 This study showed that age-associated changes in cell density are more pronounced in the NP, compared with CEP and AF, with 26 ± 4.8 × 103 cells/mm3 found in the NP during the first year of life, that dropped significantly until 2–3 years to 7.8 ± 1.3 × 103 cells/mm3, reaching 2.8 ± 2 × 103 cells/mm3 in the group with age above 60 years old. 49
Taking advantage of cell immortalization, Van den Akker and colleagues enhanced the knowledge on IVD cells, showing that NP-immortalized clones assumed three different morphologies, with 35–40% of the NP cells being described as “cobble-stone cells,” 55–56% as “wave-like cells” and 3–10% as “tiny cells.” 50 Under chondrogenic stimulus, only wave-like responder cells (NP-R) increased the expression of Sox-9 transcription factor, COL2A1 and COL1A1, as well as the production of glycosaminoglycans (GAGs). These NP-R cells were the only ones forming spheroids and expressing CD24 and GD2 markers, but no brachyury (T), suggesting they did not derive from notochordal origin. Following work revealed specific membrane markers of each cell population, further supporting the theory of higher heterogeneity in the NP compartment. 51
In 2015, the Spine Research Interest Group from the Orthopedic Research Society published a consensus paper to try to define the phenotype of NP cells, hoping to facilitate the differentiation between NP, AF, and EP cells. 52 The following NP markers were suggested: hypoxia inducible factor (HIF)-1α, glucose transporter type-1, Sonic/Hedgogh (Shh), Brachyury (T), keratin (KRT)18/19, carbonic anhydrase 12 (CA12), CD24, and ACAN/COL2 ratio >20, 52 based on their extensive use by the scientific community in the IVD field. However, for most of these markers, exclusive expression in the NP has not been confirmed, with contradictory evidence of significant differential expression between NP cells and cells from surrounding tissues, which precludes them from acting as suitable NP-specific markers. Moving forward, an improved harmonization of these details across research groups, as well as larger coordinated multisite studies aiming at an extensive characterization of these cells specifically addressing species and age-related differences, could contribute greatly to the advancement of this field.
When exploring the literature focused exclusively on the characterization of human IVD phenotype (29 articles, Supplementary Table),26,30,36–39,42,47,50,51,53–70 it remains difficult to pinpoint a set of markers for NP cells, with the set of reports analyzed producing a list of 227 markers, that was then deeply scrutinized, resulting in only 13 molecules consistently reported in human NP in more than one study: KRT18, keratin-19 (KRT19), CA12, Forkhead box protein F1 (FoxF1), Paired box protein (Pax1), COL1A1, COL2A1, collagen type XII (COL12A1), secreted frizzled related protein 2 (SFRP2), Tie2, Vascular cell adhesion protein 1 (VCAM1), Matrix Gla protein (MGP), and Integrin Binding Sialoprotein (IBSP) (Table 1).
Expression of Nucleus Pulposus Cell Markers at Gene and Protein Level. Comparison of the Expression Levels of Nucleus Pulposus Markers with Annulus Fibrosus

CA12, Pax1, MGP, IBSP, and VCAM1 can distinguish NP cells. COL2A1, COL1A1, COL12A1, SFRP2, and Tie2 can distinguish between NP and AF cells, with all, except COL2A1, being upregulated in AF cells. KRT19 remains as an NP-specific marker, being significantly more expressed in NP cells than in AF cells. From the literature, it is also possible to extract a list of molecules consistently shown to be expressed by NP cells, even if not exclusive to these cells: KRT8, KRT18, KRT19, CA12, CD24, FoxF1, Pax1, PTN, COL1A1, COL2A1, COL12A1, CD44, CD73, CD90, CD105, GD2, Gal3, CD166, VCAM1, CYTL1, DSC3, A2M, COMP, FMOD, ACAN, DSC2, GPC3, VIM, NCAM1, MGP, and LRRC17. In contrast, adult NP cells have been shown not to express CD45, CD34, and Shh.
NP cell heterogeneity has also been investigated using high-throughput techniques, such as flow cytometry. Based on cell size and auto-fluorescence, the work of Molinos et al. showed the existence of three different NP cell populations, either in healthy bovine IVDs 71 or in human aged/degenerated IVDs. 60 In human, NP cell populations were differentiated merely by their size in imaging flow cytometry, being divided into small cells (SC) (39%), large cells (LC) (43%), and super large cells (SLC) (17%). LC and SLC presented a diameter above 20 μm and were surrounded by a dense pericellular matrix. With age, SC cells tended to decrease while the frequency of LC and SLC increased, together with the pericellular matrix area. This study shows that human NP cells express CD90 and CD73, which decreased with aging, while CD146 expression remains unaltered, suggesting that a small portion of progenitor cells could still persist in the aged/degenerated IVDs. 60 Since CD73 and CD90 decrease with aging, these markers are most probably associated with SC, while CD146 that is preserved in aged cells should be more associated with LC or SLC. In bovine, NP cell populations were differentiated both based on size and auto-fluorescence, with a population (P1) of small highly auto-fluorescent cells (25–50%), a population P2 with large highly auto-fluorescent cells (45–68%) and a population P3 of medium sized lowly auto-fluorescent cells (3–8%). P3 bovine NP cells did not express CD146, while CD45, the pan-hematopoietic marker, was found to be expressed in P1 cells. 71 More recently, age-associated changes of bovine NP cells have been characterized by comparing flow cytometry NP cells from young (1 year old) and old (10–16 years old) donors. The authors have shown a reduction of CD29+, CD44+, and Tie2+ NP cells with aging, and an increase in GD2+ cells, while CD146 expression was preserved along with aging. Interestingly, the existence of four NP cell subpopulations (using FCS-A instead of FCS-H flow cytometry parameter), with different sizes, was also uncovered both in young and old donors. 41
In the last few years, single-cell transcriptomic analysis has been employed by several teams to characterize this cellular heterogenicity in the IVD, and in particular in the NP.13,73,74,76 For instance, Tu et al. reported six novel human NP cells populations based on distinct molecular signatures, termed as (1) hypertrophy chondrocyte-like NP cells, (2) effector NP cells, (3) homeostatic NP cells, (4) regulatory NP cells, (5) fibroNP cells, and (6) adhesion NP cells. These subsets were shown to have different functions within the IVD environment and potential roles in its degeneration. Briefly, effector NP cells were linked to metabolic activities and ECM synthesis, and shown to decrease with IVD degeneration while fibroNP cells increased; regulatory NP cells were involved in the regulation of inflammation and immune responses; etc. 14 Similarly, in the work of Gan et al., subclusters of IVD chondrocytes with three different functional patterns were identified: regulatory, homeostatic, and effector chondrocytes. Likewise, gene ontology analysis revealed that some subclusters participated in ECM synthesis and organization processes, while others played a protective role in stabilizing cell phenotype. 13 sc-RNA seq analysis of IVD samples from animal models, such as bovine and rat, has also allowed the identification of novel NP markers: RARRES2, S100B, SNORC, CRELD2, PDIA4, CP, RCN2, etc. These data equally lead to the uncovering of multiple heterogeneous NP subpopulations with distinct functional roles, with cell clusters specializing in biological processes such as cell survival, phenotype maintenance, ECM development, and IVD structure, under low oxygen tension and high mechanical loading.31,44 In conclusion, each study reports a different number of populations with a unique labeling system, and confirmation at the protein/functional level still requires further research. However, these data offer extremely valuable insight into the rich ecosystem found within the IVD and its changes through aging and degeneration. Moreover, these studies offer consistent evidence for the presence of immune cells in the disc. Altogether, these results highlight the dynamics of aging in the IVD cell compartment, which needs to be well understood to improve the knowledge on IVD degeneration.
IVD Myeloid Cells
In addition to the main IVD cell subtypes, a growing number of reports have stated the presence of an immune-like resident cell subset. In general, myeloid progenitor cells derive from hematopoietic stem cells present in the bone marrow, giving origin to monocytes, macrophages, dendritic cells, granulocytes (neutrophils, basophils, and eosinophils), erythrocytes, platelets, and mast cells. 77 Upon local injury or tissue damage, myeloid cells can be rapidly recruited, eliciting a strong inflammatory response. 78 These cells can also have an anti-inflammatory role, depending on the phenotype adopted in response to their microenvironment. 79 CD14, a commonly used marker for monocyte-derived cells (including macrophages and dendritic cells 80 ), has been consistently identified in human degenerated IVDs, specifically in the NP. Its expression has been associated with the presence of proinflammatory cytokines (TNF-α, IL1-β, and IL6), and pain-related molecules (NGF), 81 suggesting a link between these cells and the pathogenesis of discogenic inflammation and/or pain. CD14+ cells coexpressing CD68, another widely recognized pan-macrophage marker, have also been described in the disc, 82 further strengthening this hypothesis. Nerlich et al. detected CD68+ cells in degenerated but not in healthy human IVDs, and more frequently in NP than AF tissue, concluding that CD68+ macrophages were more likely derived from resident cells rather than recruited monocytes. 78 Miyagi et al. showed that approximately 40% of CD14+CD68+ macrophages present in the IVD were positive for CD86, another known marker for proinflammatory M1-like macrophages, 81 suggesting a proinflammatory role for these resident macrophages rather than a regulatory one. Kawakubo et al. proved that CD86+ macrophages also increased following IVD injury in mice chimeric models. 83 However, some precaution needs to be considered since CD14 has also been recognized as a marker expressed in chondrocytes. 84
Additionally, CCR7 (a M1 macrophage marker) has been identified in human cadaveric nonherniated IVDs, increasing with IVD degenerative grade. 85 On the other end of the macrophage spectrum, M2-like macrophages, more associated with an anti-inflammatory or regenerative role, have also been reported in the IVD. Nakazawa et al. described CD206 and CD163 expression in cadaveric, nonherniated IVDs. 85 Moreover, human IVD single-cell RNA sequencing (scRNA-seq) studies have identified other macrophage-associated markers, including CD40, iNOS, and CD192 for M1 polarized macrophages, and Arg1 and CD209 for M2 macrophages. 86 CD11b, also a typical phenotypic marker for monocytes/macrophages, was also identified in both human and mice IVDs. 80 Rat IVD injury was shown to upregulate CD11b, as well as pain-related inflammatory cytokines (TNF-α, IL1-β, NGF, and VEGF). 87 Nakawaki et al. also demonstrated an increase in F4/80+CD11b+ cells overtime after IVD injury. 88 All these studies build a strong case for the existence of resident macrophages in the IVD, predominantly in the NP, with their phenotype changing from more regulatory (M2) to proinflammatory (M1) with age and degeneration. Additionally, there is evidence that this inflammation may be linked to the genesis of discogenic pain.
Other type of myeloid cells, out of monocytic lineage, have also been reported in the IVD. Granulocyte-associated populations have been uncovered primarily through scRNA-seq. Granulocytic myeloid-derived suppressor cells (G-MDSCs), known to present immunosuppressive activity mainly through the suppression of T lymphocytes and production of reactive oxygen species, decrease in severely degenerated human NP tissue, showcasing their probable involvement in maintaining healthy IVD homeostasis and protecting against degeneration. ORL1, in combination with CD11b and/or CD24, was highlighted as a marker for G-MDSCs. It was hypothesized that G-MDSCs may originate from granulocyte–monocyte progenitors, which in turn express MS4A3, MPO, and ELANE markers. Furthermore, these G-MDSCs were also thought to be closely related to neutrophils, with markers CD15 and CD16b having been described in the IVD. 14 Moreover, Tryptase and Chymase, two specific types of proteases released by mast cells, have also been described in human IVD. 89 Tryptase was present in both surgical and cadaveric tissue, with higher expression in the NP and CEP than in the AF. Both markers were more prevalent in painful and/or herniated IVD (∼50%) compared to control samples (∼20%).89,90
Finally, it is also important to consider the dynamics of neovascularization and cell infiltration that occur with IVD degeneration. The infiltration of blood vessels not only brings in immune cells to the IVD, but also associates with the ingrowth of nerve fibers, and both aspects have been linked to pain symptoms.14,91 Furthermore, since blood vessels are present in the early stages of IVD development, it is also hypothesized that some remnant cells may prevail into adult stages. 13 Endothelial cells and pericytes have also been recently identified in the IVD, once more through scRNA-seq techniques. 31 Recently, a subtype of NP cells expressing higher MMP3, IL-8, CCL20, and CSF1 levels, among other markers, was associated with higher sensory innervation and pain. 92 Still, additional functional studies are needed to understand the role of this IVD resident myeloid cell population in homeostasis and degeneration. In conclusion, despite myeloid cells having been reported in the IVD and their presence associated with higher degenerative stages, suggesting them as important regulators of inflammation in the IVD, their signature is not yet completely defined, which hinders their isolation and further study.
AF Cells
Contrarily to NP, AF cells originate through the segmentation and patterning of the mesoderm, which leads to the generation of the sclerotome and, consequently, of the sclerotomal cells. 93 AF tissue exhibits a higher cell density, when compared to NP, ranging 3.0–9.0 × 106 cells/cm−3,94 with are elongated and arrange themselves into concentric circles around the NP during development. 95 AF cells also tend to decrease with aging, with a drop of about 36% in the anterior AF and 34% in the posterior AF around the 2 years of age, gradually decreasing to 1,5 ± 0,6 × 103 cells/mm3 (anterior AF) and to 1.7 ± 0.6 × 103 cells/mm3 (posterior AF). 49
This arrangement results in the organized lamellar structure of AF tissue, driven by the elongation and orientation of cells and collagen matrix. The developmental process heavily depends on the expression of growth factors such as TGF-β and the presence of mechanical loading.96,97 Still, already in the work from Chelberg and colleagues, the authors have observed that about 67% of all AF cells stained positive for both COL1 and COL2, with the remaining AF cells being divided into rarer cell populations that either only produced COL1 (14%) or COL2 (17%). These observations hinted at the existence of subpopulations of cells specifically dedicated to the production of different ECM components. 65 More recently, the work from Van den Akker revealed that, under chondrogenic stimuli, two different AF phenotypes were identified in the immortalized AF cell clones generated, based on the formation of a matrix/cell sheet in 2D (sheet-forming cells, AF-S). 67 From the literature surveyed (Supplementary Table), AF cells can express CA12, CD24, FOXF1, Pleiotrophin (PTN), COL1A1, COL2A1, COL12A1, SFRP2, CD44, CD73, CD90, CD105, CD166, AdhesionG-protein-coupled receptor L4 (ADGRL4), Desmocollin-3 (DSC3), α2Macroglobulin (A2M), cartilage oligomeric matrix protein (COMP), fibromodulin (FMOD), Desmocollin-2 (DSC2), or Tie2. Importantly, like adult NP cells, these cells did not express CD34, CD45, and Shh. Mature AF tissue is also distinctly characterized from the NP due to the presence of an anisotropic COL1 matrix, which constitutes 50–70% of tissue dry weight, along with other ECM components as proteoglycans (10–20% dry weight) and elastic fibers. 98 During IVD degeneration, AF tissue is compromised by various factors, including mechanical forces and inflammatory processes 11 that alter cell profile, namely inhibiting COL1 production,99,100 and promoting the production of inflammatory mediators (as IL-8 and monocyte chemotactic protein-1 [MCP-1]).101,102 Torre et al. have suggested that AF cells originating from the sclerotome be termed “annulocytes.” 103 Annulocytes have been associated with the production of COL1A1, COL5A1, and COL12A1.67,104 Additionally, tenomodulin (TNMD) and Mitogen-activated protein kinase kinase (MKX), markers found in different fibrous connective tissues, as well as CD146 and smooth muscle protein 22-alpha (SM22α), have been suggested to characterize a subpopulation of AF cells in the outer region of the AF tissue.57,106,107
Through scRNA-seq techniques, some authors went further and subdivided the AF tissue into two partitions: inner AF (iAF) and outer AF (oAF). New markers specific for oAF were discovered (IGFBP6, CTSK, LGALS1, CCN3), with limited expression in iAF or NP subsets. Functionally, while oAF may be associated with cell survival, most of the oAF clusters were linked to tissue development and ECM maintenance, suggesting that oAF is critical for preserving the healthy IVD structure. On the other hand, the results highlight the fibrocartilaginous nature of iAF, due to overlapping gene expression with both the oAF and NP portions. Moreover, some markers were significantly more expressed in iAF cells (COMP, GSN, SOD2, DCN, FN1, etc.), suggesting a novel role for iAF clusters was found in regulating the vasculature, circulatory, and blood vessel development, which cements these cells as a population of interest given that neovascularization is a hallmark of IVD degeneration.31,44
Despite these findings, no definitive set of markers has been established to identify and distinguish AF cells unequivocally from other IVD cell populations. While some identifying markers can be found in the previous section, further investigation into these cells in parallel with cells from the other IVD tissues will be critical to offer a clear view over the cellular dynamics in the AF.
CEP Cells
The CEP is an osseous/cartilaginous tissue linking the IVD to its adjacent vertebras. 108 These thin hyaline-like cartilage layers cover the ends of NP and the inner part of the AF. 109 At birth, the human CEPs account for approximately 50% of the IVD space, compared to adulthood, which is around 5%, and has large vascular channels passing through them. 108 Following birth, these vascular channels are filled with matrix, and no vascular vessels remain by the first decade of life, turning the IVD in the largest avascular organ in the body in adulthood. 110 In adults, the CEP is a thin layer (0.1–1.6 mm thick) of hyaline cartilage with calcified cartilage adjoining the bone.108,111 Similar to the other IVD tissues (AF and NP), the CEP is also mainly composed of water (70–80% depending on the age), followed by col type II and proteoglycans. 15 However, the proteoglycan composition in the CEP remains to be highlighted.
The cells in the CEP share morphological similarities with those in articular cartilage (AC), generally appearing rounded and slightly elongated in the direction of collagen fiber alignment. 113 Their density is approximately 15 × 106 cells/cm³ (with increased density closer to the vertebral bone), which is higher than that of the AF (two times higher) and NP (four times higher).108,114 While the CEP has traditionally received less attention compared to the AF and NP cells in IVD research, there has been a significant increase in studies focusing on CEP cells in recent years. Similarly to the NP and AC cells, CEP-derived cells have a chondrocyte-like morphology and the ability to synthesize matrix components such as COL2, ACAN, and hyaluronan (HAS2). 108 In a study by Lakstins et al., various techniques as histology, immunohistochemistry, quantitative RT-PCR, and biochemical assays, were used to analyze the CEP’s structure, composition, and cellular characteristics in comparison to adjacent tissues. When comparing the CEP to the NP, major differences were found in COLX protein expression and cell morphology, with CEP cells showing higher expression of COL2A1, ACAN, and versican (VCAN) compared to NP cells. Regarding AC, the primary distinctions were in cell morphology and gene expression profiles. CEP cells exhibited an elongated morphology aligned parallel to the vertebral body, contrasting with the round morphology of deep cartilage AC cells. In comparison to the AF, the main differences in gene profile were observed in the expression of COL2, Mohawk Homeobox (MKX), and Fibulin-1 (FBLN) gene expression. However, similarities between the CEP and AF were also found, such as cell morphology and matrix structure, indicating that the CEP may be more similar to the IVD tissues than to articular cartilage. 113 Liu et al., reported in 2001, for the first time, the existence of CEP-derived stem cells, which could be isolated, cultured, and maintained several similarities (morphology, proliferation rate, phenotype, and gene expression) with bone marrow-derived MSCs, 115 unveiling new perspectives on cell-based candidates for IVD regeneration therapies.
The CEP exhibits a distinct gene expression profile that sets it apart from other IVD components. Key markers for CEP cells include ECM proteins such as COL2A1 and ACAN, which reflect the tissue’s chondrogenic nature and proteoglycan content, respectively. 113 The transcription factor SOX9, a master regulator of chondrogenesis, is highly expressed in healthy CEP cells. 116 Signaling pathway components like ERK and BMP-related genes play crucial roles in CEP development and homeostasis. 117 Recent research has identified novel markers, including COLX, 113 CFH, 118 and FRZB, 118 which may help distinguish CEP cells from other disc cell populations. The expression ratio of COL2A1 to COL1A1 is frequently used as an indicator of cartilage health and chondrogenic differentiation across various cartilaginous tissues. While not specifically studied in CEP cells, this principle is likely applicable to CEP health as well, given the similarities between CEP cells and chondrocytes. A higher COL2A1/COL1A1 ratio is generally associated with a more chondrogenic state and better cartilage quality. This ratio has been shown to decrease in conditions such as osteoarthritis, suggesting its potential as a marker for cartilage degeneration.119,120 It is important to note that the expression of these markers can vary depending on the CEP’s state and specific cell subpopulations within the tissue. 113 Of note, no specific CEP cell surface markers were reported in the literature, demonstrating the lack of profound knowledge in these cells.
Main Challenges in the Study of IVD Cell Populations
When taking into consideration the immense IVD cell variability described so far, the difficulty in finding suitable sets of population-specific markers becomes clear. By analyzing the protocols used in the literature, aimed at finding either cell-specific markers or new IVD cell populations, it is difficult to find a standard isolation/expansion protocol or even an IVD sample selection criterion (e.g., choice of species and/or patient age/degenerative grade) between different works (Fig. 2). In the following paragraphs, we describe the main challenges in the study of IVD cells from human samples and animal models.
IVD cell source
Investigation on human IVD cells has been hampered by the lack of access and limited number of healthy discs from the same donor. Researchers try to overcome this limitation by either using: (1) discs collected postmortem from donors without history of lumbar disease (which does not guarantee absence of IVD degeneration)47,57; (2) discs collected from scoliotic patients (typically younger/adolescent patients with low/absent visible IVD degeneration) 57 ; or (3) discs from patients with varying degrees of DDD.26,121 Nevertheless, some authors have already shown alterations in the distribution of IVD cell populations with the degenerative state. For example, one (out of three) IVD cell populations seen by Chen et al. in the NP could only be found in scoliotic and degenerated discs. 122
Certain NP markers such as Gal-3, FoxF1, Pax1, CD24, KRT8, KRT18, KRT19, T, SNAP25, CDH2, and CA12 have been shown to decrease (or even disappear) with age or degeneration degree, while others (MGP, PTN, T, Shh, IBSP, LAM-A5, PDGFR, CD105, CD73, CD90, Stro1, TNMD, BASP1, and SDF-1) have been reported to increase with aging/degeneration,58,124,125 with contradictory results in the literature. For example, scoliotic discs have recently been shown to hold cell populations with very different proteomes from healthy discs (obtained from organ donors in brain dead without history of spinal disorders), with a mere 54% overlap in terms of protein content. 125 Such a difference resurges the concern over how much the source of IVD samples can affect the results obtained.
The recent advances in the field of induced pluripotent stem cells (iPSCs) differentiation into NP cells and NCs will certainly overcome the limitations associated with the use of human IVD cells.126–129 However, this is still an emerging field, with low protocol replication among different labs, high costs, and lack of knowledge on iPSC differentiation into other IVD cell subsets. Still, this is a huge gap in the field that is expected to be addressed in the following years.
Animal models, including rabbits, pigs, dogs, cows, and rodents (mice and rats), 28 have been used as an IVD cell source for decades. Nevertheless, studying IVD cell populations in these animal models compared to human sources presents significant challenges. These challenges stem from species-specific differences to methodological issues and technical limitations, all of which can significantly impact the interpretation and translation of research findings.130,131 Large animal models are described to be more similar to human in terms of cellular content and biomechanical properties in the IVD. 132 For instance, sheep and goats have emerged as valuable models, offering several key advantages that closely mimic human disc characteristics. 133 Their IVDs are more similar in size to human discs,134,135 allowing for more relevant testing of therapies and implants. Moreover, despite being both quadrupeds, spine loading pattern that more closely resembles human biomechanics is attributed to sheep.133,136 Notably, like humans, sheep and goats lose their NCs early in life, a critical similarity linked to the onset of disc degeneration in humans. 136 Yet, specific marker comparisons for sheep and goat IVD cells remain limited, highlighting the need for further research. Canine models, particularly chondrodystrophic breeds such as Dachshunds, Beagles, and French Bulldogs, offer valuable insights into IVDD due to their spontaneous development of the condition. 138 Studies on cell markers show that canine CEP differs from human CEP, with higher COL2 expression and lower expression of COL1, KRT19, MKX, FBLN1, and COLX. 113 Porcine models have gained significant attention due to their anatomical similarities to human discs. Their disc size closely matches that of humans, making them suitable for evaluating human-sized implants and devices. 139 Still, it is important to note some limitations since porcine discs retain NCs into adulthood (unlike humans) 136 and can be stiffer than human discs. 140 Porcine NP cells share some similarities with human NP cells in marker expression, including integrins α3, α6, and β4, and laminin α5. 52 Emerging large animal models, such as llamas, alpacas, horses, and kangaroos, are also being explored for their potential in IVDD research, replicating certain aspects of human disc biology more effectively than traditional models.130,141 However, further studies are needed to characterize IVD cell populations and marker expression in these emerging models to determine their relevance to human IVD biology and pathology.
Despite the variety of species that has been investigated in IVD research, rodents are by far, the animals most favored to use due to their small size, greater availability, lower maintenance requirements, and with a high number of genetic tools available, providing advantages over human source. 142 Similar to humans, rodent NP cells also express specific markers such as CD24, CD239, CD151, CD54 (ICAM), CDH2, and galectin-1 (GAL-1) KRT19, PTN at higher levels compared to AF tissue, but without significant changes throughout age, contrary to humans. 143 Several advantages are evident when considering the use of IVDs from rodent models. Rats are usually the model of choice given their larger tissue size, providing a greater number of caudal discs to collect and a larger interdiscal allowing for an easier delivery of a wider variety of treatments at doses more physiologically relevant for the human context. 144
However, translating results from rodents to humans is not straightforward due to key differences between the two models. 143 Rodents, being small quadrupeds, experience much less spinal load than humans, 145 and (like rabbits) retain a NC population during adulthood. 28 This retention supports balanced matrix turnover and helps preserve the structural integrity and function of the IVD. 146 Overall, further studies are needed to characterize how standard human cell markers alter in rodents following induced degeneration. This will ensure the reliability and translational value of rodents as preclinical models.
IVD cell isolation protocol
Other major source of variability embedded in the study of IVD cell populations is the diversity of protocols used for cell isolation. As stated in the beginning, NP or AF cell isolation is, in most cases, achieved by coupling mechanical dissociation with enzymatic digestion, commonly based on the use of collagenase. Still, there are many different collagenase types (e.g., I, II, IV, or P) available. In addition, it can be combined with other enzymes such as pronase, hyaluronidase, trypsin, or chemically defined dissociation agents (protocol details summarized in Table 2). The choice of enzymes used for the digestion of the matrix is crucial for the selection of cell populations obtained due to the specificity of each one of them. As IVD cells are intimately connected to their surrounding ECM, as evidenced by the isolation of cells still presenting a varying size of pericellular matrix, 60 the use of enzymes that digest different ECM molecules can also preferentially release specific cell populations. Moreover, it is also important to consider that enzyme concentrations and incubation times can, in some cases, vary more than one order of magnitude. Additionally, some authors report the isolation of cells without any kind of enzymatic action via explant culture, out of which cells migrate naturally.37,56 Such variability influences not only the yield of viable cells isolated from the tissue but also differences in gene expression. The latter is highly sensitive to enzyme concentration and incubation time. 147 Differences in the expression of surface markers can also be observed when an enzyme with a low substrate specificity, such as pronase, is used. 148 But, besides that, there are even smaller protocol details that can influence the cell subsets obtained, such as centrifugation steps during which specific populations are easier to lose than others. 39
Enzymatic Digestion Protocols for the Isolation of Intervertebral Disc Cells

In the case of NC isolation, these present some particular challenges, namely relating to their large vacuoles composition and network of clusters ranging in size. NP tissue rich in NC is very gelatinous and forms relatively loose pellets upon centrifugation, requiring careful handling. Additionally, NC are also difficult to count due to the higher complexity of dissociating cell clusters and accurately enumerating single cells. Williams et al. have recently investigated the recommended methodology to extract NC from isolated NP tissue of multiple species. Moreover, regarding the possibility of cryopreservation, maintaining high cell viabilities is especially challenging for NC, as their vacuoles are rich in water content, increase the susceptibility to the formation of ice crystals compared to NP cells. 149 In a different work, Basatvat et al. collected data from numerous research groups worldwide on protocols for IVD extraction and NP isolation from human, large animal, and small animal samples. Data was analyzed in order to create a set of recommendations on the most appropriate methodology to obtain NP cells, particularly aiming for high cell viability upon isolation while maintaining the natural NP-like phenotype. 142 It would be important to go deeply and improve the characterization of the IVD cells signature and function to develop targeted approaches for IVD disorders and more precise tissue engineering approaches for IVD cells.
IVD cell culture protocol
Since the number of isolated cells is limited to the amount of IVD tissue collected, in some cases, there is the need to expand IVD cells before further analysis. This additional step can greatly affect cell behavior, especially if throughout this process nondividing or slow-dividing cells are lost from the population. Moreover, it is difficult to find one common protocol. Culture medium choice is not consensual,26,38,50,150 nor is its supplementation with varying percentages of Fetal Bovine Serum (FBS) (5–20%),26,56,151 osmolality adjusting elements,152,153 or even differentiation-enabling reagents such as ascorbic acid.54,154,155 Given the harsh nature of the IVD, some researchers opt for replicating a hypoxic microenvironment (2–6% O2),38,40 while others culture cells in normoxia. 26 In addition, IVD cell seeding density can range from 5000 cells/cm2,56 to 2.1–5 million cells/cm2,154,155 which may influence the prevalence of specific cell subsets. Furthermore, most protocols culture cells in 2D, while only a few ventures in a more physiological 3D setting employ porous scaffolds, beads, and fiber meshes.152–155 Each one of these factors has the potential to influence IVD cells’ phenotype. For instance, culture in 2D monolayers has been shown to alter the morphology and phenotype of NC across multiple species, as they lose their vacuolated and clustered features as well as expression of some NC markers.156,157 Alternatively, the often-recommended option of culturing NC in 3D alginate beads is capable of maintaining cell phenotype, 158 but hinders cell expansion. 149 The effect of increasing cell passages in 2D culture also prompts changes in NP cells, with altered gene expression of multiple phenotypic markers (COL2A1, KRT18, ACAN, COL1A1), leading to the de-differentiation of NP cells overtime. 142
Comparison of several different culture media demonstrated that although both NP and AF cells are expandable in all of them, Ham’s F12 medium limited IVD cell growth. 159 Besides, Ham’s F-12 and α-MEM medium induced changes in the KRT-18 expression profile on NP cells, raising the question of whether an extended transcriptomic analysis would find other relevant alterations. Glucose levels have also been identified as crucial, with low (∼1.0 g/L) glucose levels seriously affecting IVD cell viability 160 and matrix metabolism. 161 On the other hand, high (∼4.5 g/L) glucose accelerates AF cell apoptosis 162 and decreases proliferation and downregulates stemness markers on NP progenitor cells. 163 In turn, alterations in extracellular osmolarity have been repeatedly reported to significantly alter IVD cells’ transcriptome. 100 Serum deprivation inhibits IVD cell proliferation, increases cell senescence, and induces morphological changes with cells presenting a stellate phenotype. 161 Moreover, given that FBS is a poorly defined supplement affected by substantial batch variability, several strategies have been investigated to attempt to culture NP cells without it, in order to improve consistency between different works and groups. However, a feasible alternative has not yet been identified. Le Maitre et al. reported that neither dog nor human NP cells were able to adhere well to tissue culture plastic in the absence of Fetal Calf Seru (FCS), even when performing plate coatings with laminin or fibronectin. More promising results were found for culturing human NP cells in 3D culture, in which serum deprivation did not have a profound effect on cells viability and phenotype. 142 Still, culture of specific IVD cell subsets has been only poorly investigated so far.
In summary, all the microenvironment parameters can impact on IVD cell culture and consequently on IVD cells’ phenotype. As such, moving forward, it is crucial that a consensus regarding the optimal culture conditions for the use of IVD cells in different contexts is reached, so that the data generated may be compared and/or correctly contextualized.
Overall, there is still limited information regarding the culture of IVD cell subpopulations for tissue engineering purposes. Already in 2008, Kandel et al. discussed the complexities of applying tissue engineering approaches to treat IVD degeneration, highlighting issues as the importance of mimicking the IVD unique environment. 18 In the same year, Chou et al. investigated how cells from different IVD tissues (AF inner and outer, NP) respond to 3D culture, suggesting that a single-cell type might suffice for regenerative therapies. 154 However, due to the diversity of IVD cells that has been described in the last decade, this is currently unlikely. With the advances in the bioengineering field, also related with the increasing complexity of in vitro models, it is expected that the field of IVD tissue engineering will evolve.
Conclusions
Based on the current knowledge on the heterogeneity of IVD cellular content, it is urgent to address in more detail the functional role of each IVD cell subset. A detailed analysis of the different IVD subpopulations in the context of: (i) disease initiation and progression; (ii) crosstalk with other cells in a pathological context (e.g., immune system, nervous system, vascular system) and consequent impact on pain; and (iii) response to novel advanced therapies for IVD regeneration, would certainly enrich the IVD field and contribute to refine the models and therapies under development.
Moreover, it is necessary to overcome the technical challenges faced when studying IVD cells (i.e., lack of human IVD tissue, quality/specificity of antibodies used for different species, standardization of isolation and culture protocols to maintain specific IVD cell subsets). Only overcoming these specification challenges, harmonizing the approaches in the different labs, we will enhance the knowledge on the mechanisms of disc degenerative disorders and provide a strong basis for designing targeted therapies specific for different IVD cell subtypes.
Authors’ Contributions
J.R.F. and R.M.G. conceived the idea. J.R.F. collected most of the information and drafted the article. M.L.M., S.P., C.L.P., A.L.C., and M.F. collected specific information and drafted specific sections. J.C. critically revised and reorganized the article. R.M.G. supervised the work and acquired funding. All the authors critically revised the work.
Footnotes
Funding Information
This work has been performed with funding support from Alexander von Humboldt Foundation in the framework of the Research Group Linkage Program through the project “IVDD-RESIDE: InterVertebral Disc Degeneration resolution through RESIDEnt immune cells targeting by Extracellular Vesicles from Mesenchymal Stem/Stromal Cells.” The authors also acknowledge the Portuguese Foundation for Science and Technology (FCT) (UIDB/04293/2020). A.L.C. is also thankful to IBEROSmais Project (Instituto de Biofabricación en Red para el Envejecimiento Saludable; Ref: 0072_IBEROS_MAIS_1_E).
Disclosure Statement
R.M.G., J.C., and M.F. are co-founders of FETALDISC Lda, an i3S spin-off company. But the authors declare no competing financial interests and that they did not received money to perform this work.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.