
Abstract
Osteocalcin (OC) is a bone-specific protein synthesized by osteoblasts that represents a good marker for osteogenic maturation. We examined whether in vitro osteogenic differentiation of rat bone marrow–derived mesenchymal stem cells (MSCs) could be simply assessed at earlier stages by monitoring OC secretion into the conditioned medium, rather than measuring OC deposition on the extracellular matrix (ECM), using a sandwich enzyme immunoassay system involving a specific anti-rat OC monoclonal antibody. During a 16-day culture, OC was secreted into the medium of MSCs from day 8 and increased substantially until day 16. In contrast, OC deposition on the ECM was low, even at day 13, when calcium deposition was at high levels. The histological expression pattern of OC messenger RNA provided in situ evidence that osteoblastic cells appeared at the early stages of 6 to 9 days and matured over time in vitro. Furthermore, the temporal expression of osteogenesis-specific genes, such as the transcriptional factors core-binding factor 1 and osterix, followed by increases in secretory OC proved the commitment of MSCs to osteoblastic differentiation. These results revealed that biomineralization followed secretion of OC, which may reflect early osteoblastic differentiation of cultured MSCs under osteoinductive conditions. We ascertained the osteogenic differentiation capacity of cultured MSCs in a non-destructive manner by monitoring OC secretion into the culture medium and proved that secretory OC could represent a reliable marker for predicting in vivo osteogenic potential in bone tissue engineering.
Introduction
Culture-expanded MSCs can differentiate into osteoblasts not only on polystyrene (PS) dishes, but also on the surfaces and in the pores of biomaterials.4–6 Recently, Maeda et al. 6 reported that in vitro osteogenic differentiation of rat MSCs cultured on titanium surfaces could be confirmed using real-time monitoring of the mineralized matrices produced by the MSCs during the culture period and indicated a strong positive correlation between the Ca and OC contents in the ECMs. However, quantitative analysis of deposited OC is not easy and requires several experimental processes, such as acid-extraction of OC, desalting by gel filtration, and lyophilization. Moreover, these analyses are destructive, and the cells used for the analyses cannot be employed for in vivo transplantation.
In the present study, we focused on developing a simple approach for detecting the osteogenic differentiation of cultured MSCs without damaging the cells. Because OC is a secretory protein with posttranslational gamma-carboxylated glutamic acid (Gla) residues and acts as a negative regulator of bone formation, 7 differentiated osteogenic cells must produce and secrete the protein before mineralization. Here, we measured secretory OC throughout the culture period using a specific anti-rat OC monoclonal antibody-based enzyme-linked immunosorbent assay (ELISA) system and analyzed whether secretory OC represents a good marker for predicting the in vivo osteogenic capacity during in vitro osteogenesis.
Materials and Methods
Materials
Earle's minimal essential medium (MEM) and Nonidet P-40 (NP-40) were obtained from Nakalai Tesque Inc. (Kyoto, Japan), and fetal bovine serum (FBS) was purchased from JRH Biosciences (Lenexa, KS). Dex, ascorbic acid 2-phosphate magnesium salt, and β-GP were obtained from Sigma-Aldrich Co. (St. Louis, MO). Trypsin-ethylenediaminetetraacetic acid (EDTA) solution (0.25% trypsin, 0.53 mM EDTA-4Na) was purchased from Calbiochem (San Diego, CA). Male Fischer 344 (F344) rats were purchased from Japan SLC (Shizuoka, Japan).
Cell culture of rat MSCs
Rat MSCs were isolated and maintained in primary culture as described previously.3,8 Briefly, bone marrow cells were obtained from the femoral bone shaft of 7-week-old male F344 rats and seeded into 75-cm2 flasks (T-75 flasks, Costar Co., Cambridge, MA) containing 15 mL of standard medium consisting of MEM supplemented with 15% FBS and a mixture of antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin; Sigma-Aldrich Co.). Cell cultures were maintained in a humidified atmosphere of 95% air and 5% carbon dioxide (CO2) at 37°C.
After reaching confluency (∼10 days), the cells were released from the substratum using a 0.25% trypsin-EDTA solution and inoculated into 12- and 6-well plates (Falcon, Franklin Lakes, NJ ) at a density of 1 × 104 cells/cm2 (Passage-1 (P-1)) for biochemical analyses and staining, respectively. The cells were subcultured in 1 mL of standard medium supplemented with osteoinductive supplements (10 nM Dex, 10 mM ß-GP, and 82 μg/mL ascorbic acid–phosphate) for 16 days. The culture medium was renewed every 2 days.
To examine the effects of passaging on OC secretion into conditioned medium, P-1 cells were cultured in T-75 flasks in standard medium until they almost reached confluency, and the passages were repeated until P-3.
Construction of hydroxyapatite ceramics containing MSCs
Porous hydroxyapatite (HA) ceramics (CELLYARD HA scaffolds; 50% porosity; Pentax Co., Tokyo, Japan) were cut into disks (5 mm in diameter, 2 mm in thickness), sterilized in a dry oven at 180°C for 3 h, and placed in MEM. After the air bubbles in the HA ceramics in MEM were aspirated, each ceramic was soaked in culture-expanded MSCs (1 × 106 cells/mL; P-1 or P-3 cells) for 16 h in a CO2 incubator at 37°C, resulting in the formation of HA ceramic constructs containing MSCs (HA/MSC composites), as described previously. 5
Implantation of HA/MSC composites containing P-1 or P-3 cells
We prepared two experimental groups of implants with HA/MSC composites containing P-1 or P-3 cells. These composites were implanted subcutaneously at six sites on the back of individual syngeneic 7-week-old male rats using a previously described procedure. 5 The implanted constructs were harvested 4 weeks after implantation and selected randomly from each experimental group (P-1 or P-3 cells) for histological and biochemical analyses of ALP activity and OC content.
ALP and Alizarin red S staining
For ALP staining, cultured cells were washed twice with phosphate buffered saline (PBS), fixed with 4% paraformaldehyde in PBS, and stained with naphthol-AS-MX phosphate sodium salt and fast red violet B, as described previously. 9
For Alizarin red S staining, cultured cells were washed twice with PBS, fixed with 95% ethanol, stained with Alizarin red S (0.25 g of Alizarin red S in 50 mL of PBS) for 5 min at room temperature and dehydrated in 95% ethanol.
Biochemical analyses
For measurement of protein contents and ALP activities, cultured cells were washed twice with PBS and scraped into 1 mL of 0.2% NP-40 in 50 mM of Tris-hydrochloric acid (HCl) buffer (pH 7.6) containing 1 mM of magnesium chloride. After sonication, an aliquot of each cell suspension was subjected to protein quantification using a bicinchoninic acid protein assay kit (Pierce, Rockford, IL). The same sonicated cell suspension was centrifuged at 13,000 rpm for 10 min at 4°C, and an aliquot of the supernatant was assayed for ALP activity using p-nitrophenylphosphate as a substrate, as described previously. 3 The activity was defined as the amount of p-nitrophenol released after incubation for 30 min at 37°C and normalized by the total protein concentration.
Assays of ALP activity in cultured HA/MSC composites with P-1 cells were performed as described previously. 5
For monitoring the OC levels in cultured cells in the presence or absence of osteoinductive supplements, 1 mL of medium was changed 2 days before sampling on days 4, 6, 8, 10, 13, and 16 of culture, and the cells were incubated for 48 h at 37°C. The amount of OC released into the medium by the osteoblastic cells over the 48-h incubation period was assayed using our ELISA system with an anti-rat OC monoclonal antibody, and half the measured amount was calculated as the 24-h secretion of OC. The 1-mL sample of conditioned medium obtained at each medium exchange was diluted by 1:10 and analyzed using the rat OC ELISA system. Any OC deposited on the ECM was extracted by adding 0.5 mL of 20% formic acid to the washed cell layer in each well and shaking at 4°C for 24 h. After desalting with a prepacked Sephadex G-25 column (NAP-5 column; Amersham Pharmacia Biotech AB, Uppsala, Sweden), the eluted protein fraction was lyophilized. After dissolution in 0.5 mL of ELISA sample buffer, the amount of deposited OC was assayed using the above-mentioned OC ELISA system. The average yield of OC after gel filtration through the NAP-5 column was 82%. Extraction, desalting, and assays of OC deposited on cultured HA/MSC composites were performed as described previously.
5 The 24-h deposition of OC was calculated by measuring the amount of OC deposited on the ECM at days 4, 6, 8, 10, 13, and 16 of culture and substituting the appropriate values into the following equation as described previously: 10
24-h deposition of OC (dOC; ng/well per day) at day x = (dOCx − dOCy)(x − y);x > y.
Because we showed an example [For example, the 24-h deposition of OC (ng/well/day) at day 13 can be calculated by (dOC13 – dOC10)/(13-10)] and reference No. 10, we do not have the necessity define Y.
After extraction with 0.2% NP-40, Ca was extracted from the residues by shaking in 0.5 mL of 20% formic acid for 24 h at 4°C. The total amount of Ca in the formic acid extracts was measured using the methylxylenol blue method (Calcium E-test Wako kit; Wako Pure Chemicals, Osaka, Japan). The 24-h deposition of Ca was calculated in the same manner as the 24-h deposition of OC at the same time points.
Western blotting
Rat MSCs were grown in osteoinductive medium on 12-well plates for 16 days, and the conditioned media from days 10 to l6 were pooled. The 10-mL sample of pooled medium was treated with chilled 50% acetone and centrifuged. After dialysis and lyophilization, the samples were dissolved in 1x sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer (4.3% SDS, 10% 2-mercaptoethanol, 30% glycerol, 125 mM Tris-HCl, pH 6.8, 0.01% bromophenol blue) and heated at 95°C for 5 min. For extraction of OC deposited in the mineralized matrix, the culture medium was removed, and the washed cell layers were scraped into 20% formic acid. The formic acid extracts were dialyzed against distilled water at 4°C overnight, lyophilized, and analyzed using SDS-PAGE. 11 Aliquots containing 20 μg of total protein were fractionated using SDS-PAGE using a 15% to 25% polyacrylamide gradient gel (MultiGel; Daiichi Pure Chemicals Co. Ltd., Tokyo, Japan) and electrophoretically transferred to a nitrocellulose filter (Advantec Co., Tokyo, Japan). Immunoblotting was performed using the following three monoclonal antibodies: an anti-rat OC antibody (LXP-5A8) raised against a synthetic peptide corresponding to a rat OC sequence (residues 16-30; 3 Gla residues); an anti-bovine OC antibody (Gla-specific mAb; OC4-30; Takara Bio Inc., Otsu, Japan); and an anti-human OC antibody (Glu-specific mAb; OC4-5; Takara Bio Inc.). Diluted horseradish peroxidase–conjugated goat anti-mouse immunoglobulin (Ig)G (IBL, Takasaki, Japan) was used as the secondary antibody. Detection of antigen–antibody complexes was performed using a Western Lightning Chemiluminescence Reagent Plus Kit (PerkinElmer LAS Inc., Boston, MA) at room temperature for 1 min, followed by exposure to Hyperfilm ECL (Amersham Pharmacia Biotech UK Ltd., Amersham, United Kingdom) for 30 to 60 s.
Isolation of total RNA and quantification of messenger RNA using real-time quantitative polymerase chain reaction
Total RNA was isolated from cell layers using an ISOGEN RNA Extraction Kit (Nippon Gene, Tokyo, Japan). The extracted total RNA (1–2 μg) was converted into complementary DNA (cDNA) with avian myeloblastosis virus reverse transcriptase XL (RNA PCR Kit; Takara Bio Inc.) and regarded as a template for the cDNAs in the sample.
The theoretical basis of real-time quantitative polymerase chain reaction (PCR_ using an ABI PRISM 7700 Sequence Detection System (Applied Biosystems, Norwalk, CT) has been described elsewhere. 12 The mRNA levels of ALP, OC, osteopontin (OP), bone morphogenetic protein-2 (BMP-2), osterix, core-binding factor (Cbfa)1 type 1, Cbfa1 type 2, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were amplified using real-time PCR using the respective primers and fluorogenic probes shown in Table 1. Bone sialoprotein (BSP) mRNA was quantified using a TaqMan Gene Expression Assay Kit (Rn 00561414-m1; Applied Biosystems) for rat BSP. Known amounts of rat cDNA fragments of ALP, BSP, OC, OP, BMP-2, osterix, Cbfa1 type 1, Cbfa1 type 2, and GAPDH were used as standards. The target mRNA levels were compared after correction for the corresponding GAPDH mRNA level as an internal standard to adjust for differences in the efficiencies of reverse transcription between samples.
Rn 00561414-m1 (ABI).
In situ hybridization
A digoxigenin (DIG)-11–[5′]-uridine-5-triphosphate (UTP)-labeled single-stranded RNA probe for rat OC was prepared using a DIG RNA Labeling Kit (Roche Molecular Biochemicals, Mannheim, Germany) according to the manufacturer's instructions. Two clones containing a 360-bp Eco RI fragment of a rat OC cDNA, designated pSP6/T7/rev OC-DNA and pSP6/T7/OC-DNA, were used to generate antisense and sense probes, respectively, according to the action of SP6 RNA polymerase. Subcultured cell layers on 4-well glass chamber slides (Lab-Tek Chamber Slide System; Nalge Nunc International Co., Naperville, IL) were fixed with 4% paraformaldehyde in PBS and hybridized with the probes, as described previously. 3
Histological examination
Implanted specimens were fixed in buffered formalin for 3 days and decalcified with K-CX solution (Falma Co., Tokyo, Japan). After dehydration through a graded series of ethanol solutions, the specimens were embedded in paraffin and cut into 8-μm sections at the middle of each specimen, parallel to the round surface of the HA disk. The sections were stained with hematoxylin and eosin.
Statistical analysis
The values for ALP activity, total protein, Ca and OC contents, and mRNA levels were calculated as means and standard deviations. The significance of differences between two experimental groups in the presence or absence of osteoinductive supplements was evaluated using the Mann-Whitney U-test. Values of p < 0.05 were considered to indicate statistical significance.
Results
ALP activities and Ca contents in the ECMs of cultured MSCs
Numerous nodules positively stained for ALP activity were observed macroscopically on each PS dish after 8 to 16 days of subculture (Fig. 1A), whereas little Ca deposition was detected at day 8 (Fig. 1B). Because the total protein contents were similar in cells cultured with and without osteoinductive supplements during the culture period (Fig. 2A), the corresponding protein contents normalized ALP activities. Under osteoinductive conditions, the ALP activity of subcultured MSCs began to increase at day 8 of culture, peaked at day 10, and then declined at day 16, and these activities were significantly higher than those of MSCs subcultured under non-osteoinductive conditions (Fig. 2B). Ca deposition increased exponentially after day 10 of culture under osteoinductive conditions, whereas the levels under non-osteoinductive conditions remained low throughout the culture period (Fig. 2C).
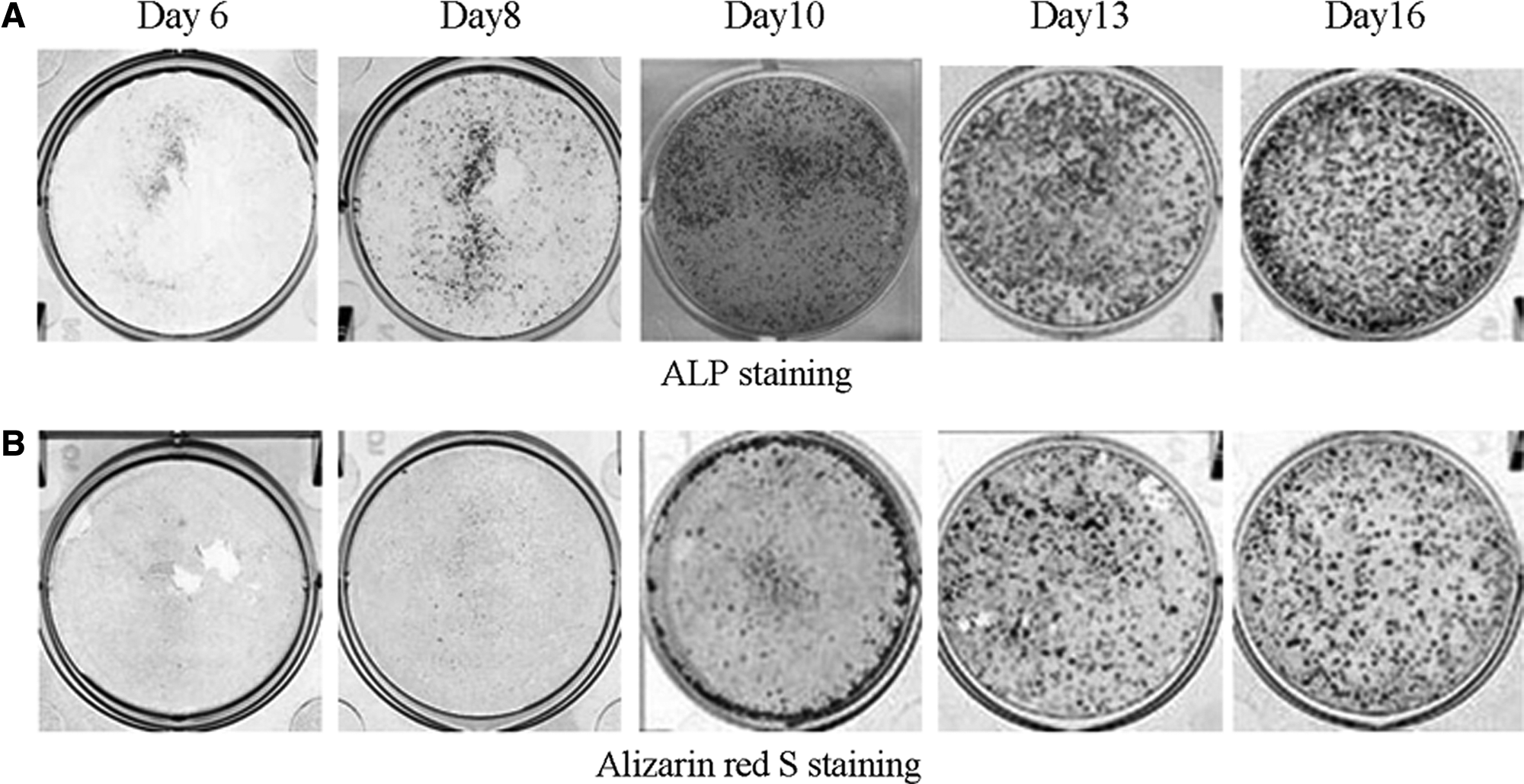
Photographs of mesenchymal stem cell layers stained for alkaline phosphatase (ALP) activity (

Total protein contents (
Secretion and deposition profiles of OC in cultured MSCs
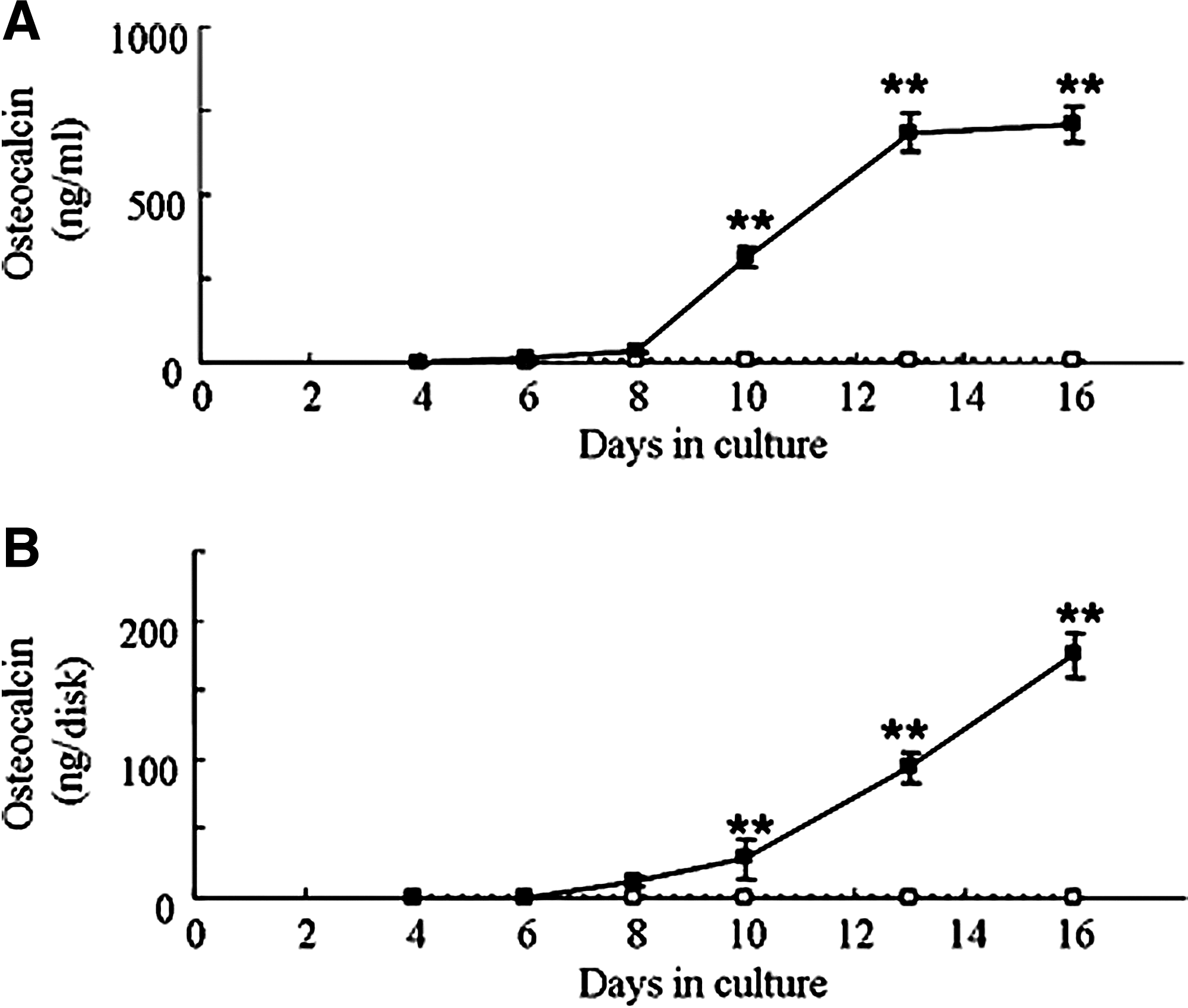
ELISA analysis revealed that OC was continuously secreted into the culture medium under osteoinductive conditions after day 8 of culture and increased until day 16 (Fig. 3A). In contrast, deposition of OC onto the ECM was observed after a lag period of 2 to 3 days (Fig. 3B). Furthermore, the amount of OC secreted into the culture medium per well was three times as high as the amount deposited on the ECM. The time course of the 24-h secretion of OC into the conditioned medium under osteoinductive conditions is shown in Figure 3C. OC secretion began at day 8 of culture, and the amount of secreted OC subsequently increased at a variable rate throughout the 16-day culture period. The time courses of the 24-h depositions of OC and Ca onto the ECM of the cultured MSCs are also shown in Fig. 3C. Sharp increases in the 24-h depositions of OC and Ca were observed at day 13, which represent biomineralization. These data suggest that the beginning of biomineralization of the ECM was significantly later than the osteoblastic differentiation of MSCs. At day 16 in culture, the 24-h deposition of OC had decreased, whereas the 24-h OC secretion and 24-h Ca deposition levels were to the same as or greater than their levels at day 13.

Amounts of osteocalcin (OC) secreted and deposited by mesenchymal stem cells (MSCs) cultured under osteoinductive (solid squares) or non-osteoinductive (open squares) conditions. (
Next, we investigated the molecular features of the secreted and deposited OC using western blotting using three monoclonal antibodies. In Figure 4A, OC was assessed using immunoblotting with an anti-rat OC monoclonal antibody (LXP-5A8; Gla type). Secreted (lane 2) and deposited (lane 3) OC appeared as single immunoreactive bands after SDS-PAGE with similar intact molecular weights of approximately 7 kDa. As shown in Figure 4B, secreted OC also exhibited intense immunoreactivity for the other Gla-specific mAbs used but not for the Glu-specific mAb, for which no band was detected (Fig. 4B, lane 4). These results confirmed that the de novo–synthesized OC from cultured rat MSCs was a Gla-containing intact protein.
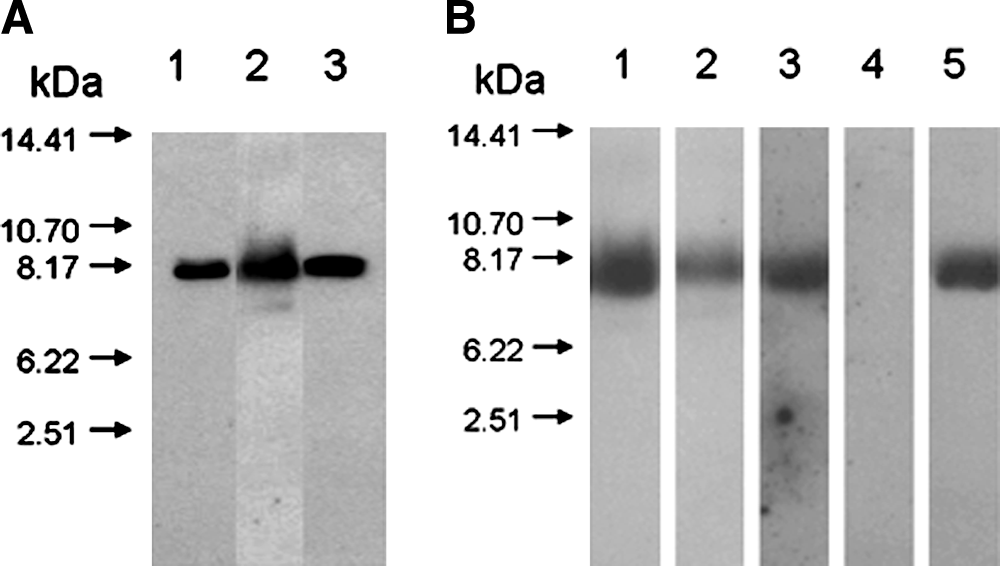
Western blot analysis of de novo–synthesized osteocalcin (OC) in the conditioned medium and extracellular matrix (ECM) of cultured mesenchymal stem cells (MSCs). Conditioned medium and ECM were prepared from MSCs cultured under osteoinductive conditions for 16 days. Aliquots (20 μg) of total protein were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis followed by western blot analysis using a specific anti-rat OC monoclonal antibody and a chemiluminescence detection kit. (
Osteogenic phenotype gene expression in cultured cells
Based on the above results, we explored osteogenic phenotype gene expression in cultured MSCs (Fig. 5). During culture on PS dishes in osteoinductive medium, ALP and OC transcripts were detected at days 6 and 8, respectively, and their time-dependent gene expression patterns were similar to those of their protein expressions (ALP activity in Fig. 2B and secretory OC contents in Fig. 3A). Because BSP is considered to play prominent roles in cell adhesion and initiation of mineralization, its gene expression was investigated. In the osteoinductive medium, BSP mRNA began to be significantly expressed on day 6 of culture and was maintained at high levels until day 16. Transcription factors involved in promoting OC transcriptional activity (Cbfa1 and osterix) were also expressed at the early stages of cell culture in the osteoinductive medium. Although OP is considered to be expressed earlier than OC to promote osteoblastic adhesion, our results revealed that OP mRNA expression peaked on day 8 of cell culture. BMP-2 was expressed during the initial stage of cell culture (days 4–6), and its expression increased after the mid-stage of the cell culture (Fig. 5).

Temporal changes in the messenger RNA expressions of alkaline phosphatase (ALP), osteocalcin (OC), bone sialoprotein (BSP,) osteopontin (OP), core-binding factor (Cbfa)1 type 1, Cbfa1 type 2, bone morphogenetic protein-2 (BMP-2), and osterix in rat mesenchymal stem cells (MSCs) cultured under osteoinductive (solid squares) or non-osteoinductive (open squares) conditions. Aliquots (1–2 μg) of total RNA extracted from cultured cells from days 4 to 16 were transcribed into complementary DNA and subjected to real-time quantitative polymerase chain reaction analysis. The relative messenger RNA (mRNA) levels are expressed as arbitrary units normalized according to the corresponding levels of rat glyceraldehyde-3-phosphate dehydrogenase mRNA. Values are shown as means ± standard deviations (n = 4). **p < 0.01.
In vitro histological expression of OC during MSCs differentiation
To ascertain whether osteoblastic differentiation could be evaluated according to early secretion of OC into the culture medium, OC mRNA expression was analyzed according to in situ hybridization over an extended culture period from days 6 to 15. The specificity of the hybridization was determined using DIG-labeled antisense and sense RNA probes. Under osteoinductive conditions, signals for OC mRNA began to be detected at day 9 in the periphery of early multilayered nodules where the ECM was not mineralized (Fig. 6, day 9). The number of OC mRNA-expressing cells increased markedly thereafter until day 15, when mineralized nodules associated with intense signals for OC mRNA covered most of the tissue culture dish (Fig. 6, days 12 and 15). In contrast, no signals were detected in the cultured cells at day 15, when the sense RNA probe was used for hybridization (data not shown). Therefore, the bone marrow–derived MSCs clearly exhibited the appearance of early osteoblasts at day 9, followed by an increase in the number of cells. In situ hybridization of cell layers from cultured rat MSCs revealed that OC mRNA could be detected in the osteoblastic cells before obvious biomineralization. This temporal expression pattern of OC mRNA detected according to in situ hybridization was clearly reflected in the real-time quantitative PCR analysis of OC mRNA expression in cells cultured for a period of 16 days.

Osteocalcin (OC) messenger RNA (mRNA) expression in rat mesenchymal stem cell cultures evaluated using in situ hybridization. Representative photomicrographs of cell layers grown under osteoinductive conditions for 6 to 15 days are shown. Fixed cell layers after 6, 9, 12, and 15 days of culture were hybridized using a digoxigenin (DIG)-labeled antisense RNA probe for OC. Nuclei were stained with methylene blue after the hybridization. Positive signals (dense dot) for OC mRNA appear at the periphery of unmineralized nodules (colony area) at day 9 of culture and increase in intensity with increasing numbers of mineralized nodules during the longer culture periods. No signals were observed when a fixed cell layer from day 15 was hybridized using a DIG-labeled sense RNA probe for OC (data not shown). Bars, 100 μm. Color images available online at
Secretion and deposition profiles of OC in cultured HA/MSC composites
Figure 7 shows the differences in OC secretion between HA/MSC composites cultured in the presence and absence of Dex. The composite was made using primary cultured MSCs. OC secretion from the HA/MSC composites cultured in the osteoinductive medium was much higher than that from HA/MSC composites cultured in the non-osteoinductive medium and further increased after day 6 (Fig. 7A). Furthermore, the timing of OC secretion into the conditioned medium was earlier than that from MSCs cultured on plastic dishes. OC deposition on the cultured HA/MSC composites increased after day 8 (Fig. 7B).
Amounts of osteocalcin (OC) secreted and deposited by hydroxyapatite/mesenchymal stem cell (HA/MSC) composites cultured under osteoinductive (solid squares) or non-osteoinductive (open squares) conditions. (
In vitro and in vivo osteogenic potential of MSCs at P-1 and P-3
To confirm whether OC secretion can reflect in vivo osteogenic potential, we used P-1 and P-3 cells for in vitro and in vivo bone-formation experiments. As shown in Figure 8A, cultured P-1 MSCs secreted large amounts of OC, whereas cultured P-3 MSCs exhibited very low levels of OC secretion. Next, P-1 or P-3 cells were combined with porous HA ceramics (HA/MSC composites), and the composites were implanted at subcutaneous sites in male rats. Four weeks after implantation, the composites containing P-1 cells showed strong in vivo bone formation (Fig. 8B), as well as high ALP activity and OC contents (Fig. 8C). In contrast, the composites containing P-3 cells showed fibroblast-like cells without any bone formation and did not exhibit any osteoblastic activities.
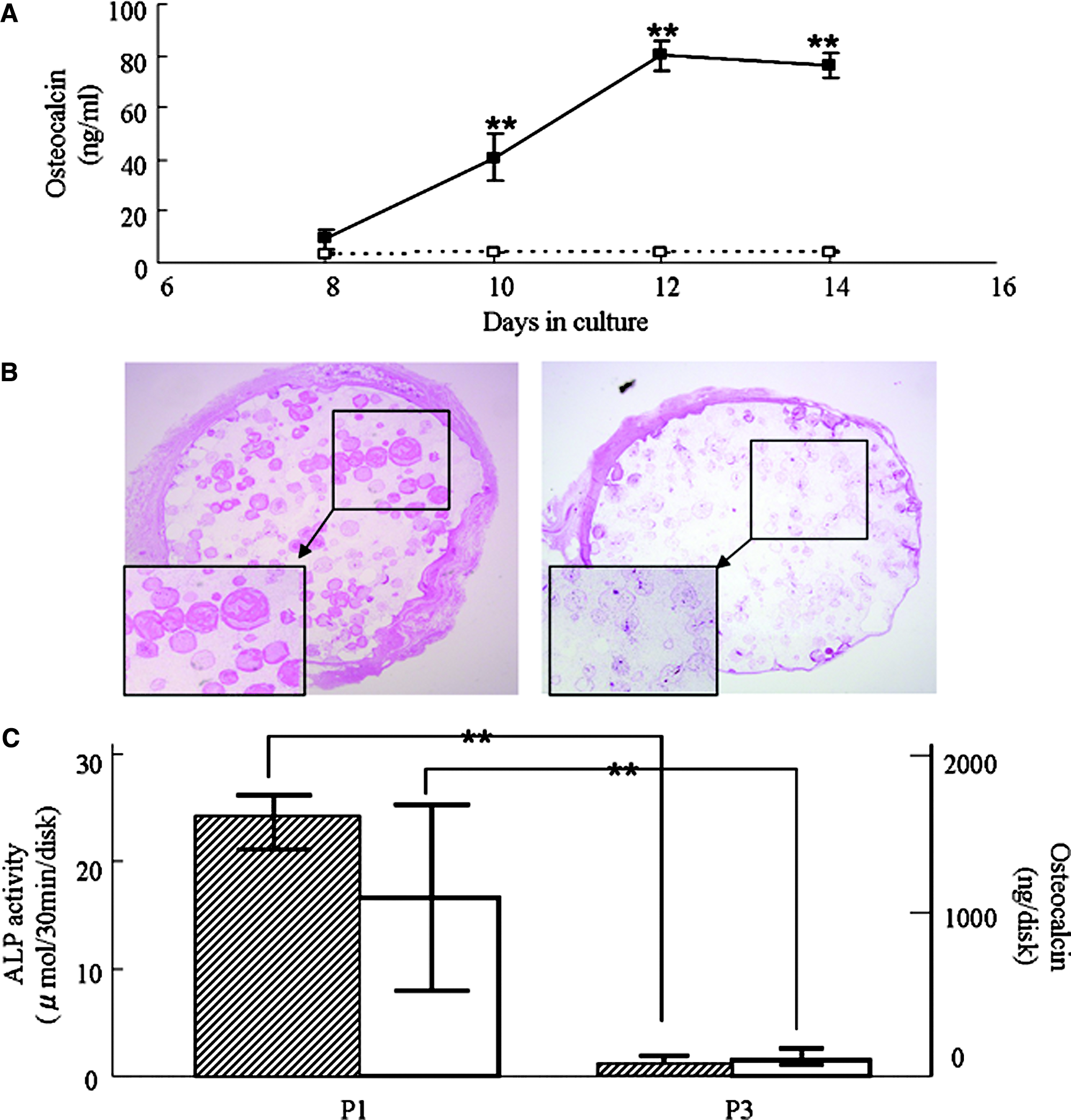
(
Discussion
Recently, bone marrow–derived MSCs have been widely employed for bone tissue regeneration because of their bone-forming capability.13,14 In vivo formation of bone by implanting not only proliferated MSCs, but also MSCs subcultured in scaffolds, such as porous HA ceramics, in osteoinductive medium has previously been demonstrated.15,16 Therefore, there are two types of the tissue-engineering approaches for bone regeneration; one uses cultured MSCs, and the other further cultures the MSCs to show osteoblastic phenotype in the scaffolds (fabrication of cultured bone). The clinical applications of the MSCs has been previously reported, andthe usefulness of the cultured bone for various patients has also been reported.17,18 In contrast to the MSCs' application, bone-forming osteoblasts already exist in the cultured bone; therefore, the cultured bone can show immediate new bone-forming capability after in vivo implantation. However, a drawback of using the cultured bone is how to assess the osteogenic activity to ascertain the existence of viable and active osteoblasts in the scaffolds before their implantation. The present article demonstrates a non-invasive method of assessing osteogenic activity by measuring OC in the culture medium; the measured OC is a genuine osteoblastic marker, as seen below.
We previously reported that rat and human MSCs derived from bone marrow could differentiate into active osteoblasts that produced bone matrices on PS dishes or bioactive ceramics under osteoinductive conditions in the presence of Dex, ascorbic acid 2-phosphate, and β-GP.3,15,16,19 We focused on OC in the present study, because it specifically exists in hard tissues, such as bone and teeth, and is regarded as a marker for the differentiation and maturation of osteoblasts.20–22 Physicochemical and biochemical analyses have revealed that OC deposition and mineralization of in vitro–cultured bone matrices represent OC newly synthesized by differentiated osteoblasts and biological accumulation of apatite, which exists in natural bone, respectively. 3 However, these methods are destructive toward cultured cells, and the cells used for the analyses are invariably lost. As a non-destructive method, confocal laser scanning microscopy has been used to analyze and visualize the three-dimensional structure of the calcified bone matrix constructed during the final stage of in vitro osteogenic differentiation of rat MSCs. 23 The degree of calcification during the last stage of bone differentiation has been evaluated by measuring fluorescence intensity after adding calcein, a Ca-chelating agent, to rat MSCs cultured on the surface of titanium alloy culture dishes in osteoinductive medium. 6 These experiments not only revealed a strong positive correlation between the amounts of Ca and OC deposition (main constituents of the bone ECM), but also determined that the titanium alloy used represented an excellent and efficient material for bone differentiation. The methods in both of the above-mentioned reports allow successful assessment of mineralization by viable cells, because the addition of calcein does not adversely affect the viability of cultured cells. Therefore, methods using calcein represent non-destructive ways of measuring the amount of mineralization. However, this method simply measures the ECM contents and does not evaluate the cellular activity of osteoblasts. In the present study, we have proposed a valuable assessment method for selecting active osteoblastic cells that is simple, reliable, and quantitative for osteoblastic activity, without any procedures that might damage the cells. We achieved this by culturing expanded rat MSCs and MSCs in scaffolds under osteoinductive conditions and quantitatively monitoring the amounts of secretory OC in the conditioned medium throughout the culture period using a specific anti-rat OC monoclonal antibody-based ELISA system.
In accordance with previous studies on in vitro MSC differentiation, our MSCs cultured under osteoinductive conditions for 16 days showed high expression of differentiation markers, ALP activity, and Ca deposition. OC was detectable in the conditioned medium at day 8, and the amount of OC secretion increased dramatically thereafter. On the other hand, OC deposited in the ECM was not detectable until day 10. Thus, OC deposition had a 2- to 3-day lag time after the onset of OC secretion. Deposited OC was first detected in the late differentiation stage at day 13 and increased as calcification progressed during days 13 to 16. These findings demonstrate that the amount of OC deposition is closely related to biomineralization or hydroxyapatite crystallization. However, the amount of deposited OC within the ECM per well was low even after 13 to 16 days in culture, despite the abundant secretion of OC. According to our recent report, 24 the levels of OC secreted by MSCs cultured in medium without β-GP at day 14 were comparable with or higher than the levels secreted by MSCs cultured in osteoinductive medium, despite the fact that Ca deposition did not occur during the culture of MSCs in the absence of β-GP. These results suggest that the osteoblastic differentiation of MSCs occurs earlier than and independent of calcification.
We compared the molecular features of the secreted and deposited OC using western blot analysis. The molecular sizes of the two proteins were similar and coincided with that of OC purified from rat cortical bone. The proteins were identified as Gla-type OC, which contains three Gla residues with a high affinity for HA, because three anti-Gla type OC monoclonal antibodies that bind to the center of the Gla-containing domain corresponding to amino acid residues 16 to 30 recognized them. These findings lead us to surmise that the differentiated osteoblasts newly synthesize the OC secreted into the conditioned medium and the OC deposited in the cell layer, which reflect their differentiation into mature osteoblasts.
To confirm osteogenic differentiation, the gene expression patterns of osteoblastic markers were characterized using quantitative reverse transcriptase PCR. Specifically, we evaluated the expression levels of ALP, OC, BSP, Cbfa1, osterix, OP, and BMP-2 mRNAs. BSP mRNA expression was detected at an early stage of culture and maintained at high levels until day 16, whereas the peaks of Cbfa1, osterix, and OP mRNA expression coincided with the time when OC was secreted into the culture medium. BSP is known as a marker of calcification, as well as a cell adhesion factor.25,26 However, it is also expressed in macrophage, lung, mammary gland, kidney, and some cancers. On the other hand, OC has been reported to exist in hard tissues, such as bone and teeth, and is regarded as a marker of the differentiation and maturation of osteoblasts.21–23 Therefore, OC is a more suitable osteoblastic marker than BSP. Thus, in the present study, we focused on OC, and especially its protein level in the culture medium, as an early marker of osteoblastic differentiation.
We examined OC mRNA expression at the single cell level under osteoinductive conditions using in situ hybridization and detected early expression in cuboidal cells around cellular clumps at days 6 to 9, when mineralization had not yet started. Many OC mRNA-positive cells were observed in large clumps (nodules) at day 12, followed by calcification of the nodules at day 15. Therefore, the onset of OC gene expression closely matched that of OC secretion into the medium and occurred before the onset of calcification.
Moreover, to assess the osteogenic activity of cultured bone with porous HA for bone tissue engineering, we monitored the levels of OC secreted into the conditioned medium by cultured HA/MSC composites. OC secretion by HA/MSC composites cultured in the osteoinductive medium was higher than that of MSCs cultured on PS dishes and increased in a time-dependent manner, whereas HA/MSC composites cultured in the non-osteoinductive medium showed low OC secretion during the culture period. Using the same porous HA ceramics, we previously reported that in vivo bone formation capacity increased with increasing levels of OC deposition in cultured HA/MSC composites. 5 Because OC secretion reflects OC deposition, monitoring of OC secretion into the culture medium may predict the in vivo new bone formation capability of HA/MSC composites.
We used dish-adherent cultured bone marrow cells (referred to as MSCs) and found high proliferative and osteogenic differentiation capability of the cells. Nevertheless, definitive markers of MSCs have not been established, and the MSCs used in this study are therefore heterogeneous populations. In this regard, many studies have reported decreasing capability for osteogenic differentiation, as well as low mineralization ability after repeated passages of MSCs.6,27 Therefore, if monitoring of OC secretion in the culture medium can predict the osteogenic capability of MSCs, the low secretory OC levels observed in medium from passaged MSCs may indicate insufficient bone formation capability after in vivo implantation. As shown in our data, when P-1 and P-3 cells were cultured on PS dishes under osteoinductive conditions for the culture period, OC secretion from P-1 MSCs was significantly higher than that from P-3 MSCs. To evaluate in vivo osteogenesis, HA/MSC composites containing P-1 or P-3 cells were implanted subcutaneously into syngeneic rats. High levels of osteogenic markers, such as ALP activity and OC content, and histological osteogenesis were prominent in HA/MSC composites containing P-1 cells but almost absent in HA/MSC composites containing P-3 cells. These results clearly demonstrate the importance of measuring secretory OC in the medium to assess the osteogenic activity of cultured MSCs.
Several previous reports have examined the effectiveness of undifferentiated MSCs as a source of the cultured bone. This approach requires in vitro culturing of MSCs on high-bioaffinity inorganic carriers, such as porous HA and alumina, 15 or on organic carriers, such as collagen, 28 poly (DL-lactide-co-glycolide), 29 and temperature-sensitive gels. When considering hard tissue regeneration using the cultured bone, it is important to perform rapid assessments of the high osteogenic capacity and viability of cells cultured on various scaffolds. As shown in the present article, secretory OC is a good non-invasive marker for assessing osteogenic capacity at early stages, because it can be easily measured without any loss of cells and can be detected before mineralization. Therefore, the present analytical method can be integrated into the process of bone tissue regeneration.
Footnotes
Acknowledgments
We thank Ms. R. Hirano and Ms. M. Yoshimura (Department of Public Health, Health Management and Policy, Nara Medical University School of Medicine) for their technical assistance.
Disclosure Statement
No competing financial interests exist.