
Abstract
In parallel to the active search for therapeutic and industrial applications of human embryonic stem cells (hESCs), designing automated means of producing those cells is a timely goal. Slow-turning lateral vessels (STLVs) with low shear stress have shown promise for expanding the cells at the embryoid body stage. We have improved this technology by developing two complementary systems, allowing continuous optimization of the culture conditions. First, perfused STLV bioreactors were set up, to provide continuous delivery of culture medium to the cells growing in the rotating chamber. This allowed the external control of the culture medium, and consequently optimized oxygenation, pH, nutrient supply, and waste elimination. Second, a dialysis chamber was adapted. This led to a further enhanced controlled environment and a decrease in the quantity of adjunct products (e.g., growth factors) necessary to the cells inside the bioreactor chamber. hESC aggregation and initial differentiation—taking neural induction as an example—were compared between the perfused and dialyzed STLV system and static cultures. Perfused and dialyzed STLV bioreactors promoted formation of embryoid bodies that were differentiated more rapidly and were homogeneously synchronized in a statistically significant manner.
Introduction
Fulfilling these promises requires, however, making full usage of both capacities of hESCs for unlimited expansion and pluripotency. For that purpose, these two abilities have to be translated into technological platforms for cell production and controlled differentiation. Conventional stirrer vessels have the disadvantage of generating shear forces, and although manageable, these forces still damage the cells. Low-shear-stress bioreactors exist, and pioneer demonstration has been given of mass production of embryoid bodies derived from human embryoid bodies (hEBs). 4 This success relied on specific techniques used to ensure dynamic, yet mild, suspension conditions to control the aggregation processes of differentiating hESCs.
Building upon these recent developments, the present study was undertaken to promote optimizations required for therapeutic or industrial implementation of hESC progeny production, focusing on the precise control and reproducibility of cell culture conditions. Fine tuning and maintenance of cell culture conditions is a requisite for traceability of industrial processes and safety of clinical-grade cell therapy products.5,6 Our first goal was therefore to make the closed chambers of the bioreactors fully accessible to external analysis and to addition of exogenous product, in order to provide a workable platform for implementation of any requested control and medium adaptation, without any disruption of dynamic cell suspension. In addition, our study addressed the complementary issue of the renewal of large quantities of culture medium over time. Cell production in bioreactors requires a large volume of culture medium and, in parallel, increases considerably the amount of adjunct product, some of which—particularly cytokines—are expensive.7,8 Accordingly, our second goal in this study was the development of a system that would allow costs to be reduced by limiting the amount of adjunct product to the desired concentration in the strictly limited, necessary volume of medium.
We propose here technological developments that have been tested on a rotating slow- turning lateral vessel (STLV) and evaluated on the neural differentiation path but may be adapted to any type of low-shear-stress bioreactor or initial steps of hESC differentiation.
Methods
hESC culture
The hESC lines VUB01 derived at the Vrije Universiteit Brussels, 9 SA01 distributed by Cellartis (Sweden), H9 (WA09, WiCell Research Institue), and HUES-9 octamer (OCT)-4GFP, in which green fluorescent protein (GFP) is under the control of the full-length POU5F1 (OCT-4) promoter, kindly given by Chad Cowan (Harvard Stem Cell Institute), were used during this study. Cells were maintained on a feeder layer of mitomycin C-inactivated murine Sim's Thioguanine Ouabaine–resistant fibroblasts in knock-out (KO)–Dulbeccos' modified Eagle medium (DMEM) supplemented with 20% KO serum replacement, 1 mM L-glutamine, 0.1% penicillin/strepto-mycin, 1% non-essential amino acids, and 4 ng/mL fibroblast growth factor-2 (FGF2) (all from Invitrogen, Cergy, France). Culture medium was changed by half daily, supplemented with 8 ng/mL FGF2. For passaging, cells were harvested using collagenase type IV (1 mg/mL, 5 min) and gently scraped with a plastic pipette. To separate isolated cells from feeder cells and clumps a passive sedimentation was performed and hESCs were then filtered through a 70 μm cell strainer (Beckton Dickinson, Le-Pont-de-Claix, France) was performed to calibrate the aggregate's size. hESCs were re-suspended and seeded at a 1:5 ratio in a 300-cm2 flask.
After 7 days, approximately 50 million undifferentiated hESCs per flask were harvested using 5 mL of collagenase type IV and dispase I (at 1 and 0.3 mg/mL, respectively) at 37°C, 5% carbon dioxide (CO2) for 20 min. After elimination of remaining feeder cells using 100-μm filtration, hESC colonies were broken into small clumps and filtered again onto a 70-μm cell strainer before size control under the microscope.
hESCs were seeded in the bioreactor chamber at approximately 0.5 million cells per mL in hESC medium without FGF2. The bioreactor was set to rotate at 12 rpm, and the speed was increased daily by 1 rpm to a plateau at 20 rpm. The perfusion flow was set to renew all medium in the chamber within 24 h (10 rpm). Control hEBs formed under static culture conditions (SCC) were seeded at the same concentration to obtain the same rate of aggregation; medium was replaced by half daily.
STLV bioreactors: set-up and function
The perfused STLV system (Fig. 1) included an autoclaved 55-mL-wide chamber, variable speed motor drives with tachometers, a culture tank, a peristaltic pump, and a silicone membrane oxygenator (all from Synthecon, Cellon SA, Bereldange, Luxembourg). All components were connected using flexible silicone tubing. The medium outlet used was covered using a dialysis membrane with a 100-kDa–molecular weight cut-off at the inner cylinder of the perfused STLV to keep cells out.

Diagram of the perfused and dialyzed slow-turning lateral vessel (STLV) bioreactor. The dialysis loop (in grey) comprises the medium tank, a pump, and the outer part of the dialysis chamber equipped with a semi-permeable membrane. The culture loop (in black) comprises the STLV bioreactor, a pump, an oxygenator, a bubble trap, a bioanalyzer, and the inner part of the dialysis chamber. Only the STLV chamber rotates.
Culture conditions were controlled online by connection of the medium used at the outlet of the STLV to a Bioprofile 400 (Nova Biomedical, les Ulis, France), allowing non-invasive follow-up of the culture parameters. Online analysis was performed for pH, partial pressure of oxygen (O2), partial pressure of CO2, osmolarity, concentration of glutamine, glutamate, glucose, lactate, sodium, potassium, and ammonium. The analyzer was programmed to initiate calibration cycles at regular intervals every 6 h.
For EB sampling, the rotating vessels were stopped and placed on a clean bench to allow the cell aggregates to settle and were taken away by pipetting. hEBs were individually and randomly retrieved from the chamber of the STLV bioreactor or control SCC after 2, 4, and 8 days. Mixed samples were also retrieved on the same days. For retrieval from the STLV chamber, 500 μL of medium was retrieved and the hEBs filtered out using a 70-μm-pore nylon cell strainer (Becton Dickinson, Le Pont de Claix, France). The strainer was rinsed with phosphate buffered saline (PBS) to deliver the EBs to a Petri dish. Resulting aggregates were individually collected under a stereomicroscope and each placed in 100 μL of RNeasy Lysis buffer (RLT) (Qiagen, Courtaboeuf, France). Mixed samples were formed using the remaining non-individually collected filtered aggregates; they were centrifuged (900 rpm, 1 min) and collected in 1 mL RLT.
Neural differentiation protocols
Differentiation of the H9 cell line along the neural lineage was performed in two different manners. First, hEBs produced in STLV conditions were plated after 6 days of aggregation onto poly-ornithine/laminin-coated (POL) culture dishes in DMEM/F12 with N2 supplement. Second, neural progenitors were obtained from undifferentiated hESCs by co-culture with MS5 stromal cells, as described previously. 10 In both protocols, morphologically identified neural rosettes were isolated mechanically for analysis (n = 4).
Real-time reverse transcriptase polymerase chain reaction
Total RNA was isolated from hESCs (undifferentiated) and hEBs (differentiated) using RNeasy extraction kits (Qiagen) according to the manufacturer's protocol. Quality and concentration of RNA were checked using the Nanodrop technology. Reverse transcription was performed using the Superscript II reverse transcription kit (Invitrogen).
For all samples, including isolated hEBs, real time reverse transcriptase polymerase chain reaction (PCR) was performed using a Chromo4 real-time system (Bio-Rad, Marne la Coquette, France) and SYBR Green PCR Master Mix (Applied Biosystsem, Courtaboeuf, France) following the manufacturer's instructions. Quantification of gene expression was based on the cycle threshold value calculated using Opticon Monitor software (Bio-Rad, Marnes-la-Coquette, France). Melting curve and electrophoresis analysis were performed to control PCR product specificities and exclude non-specific amplification. PCR primers are listed in Supplemental Table 1 (available online at
Fluorescence-activated cell sorting analysis
Hues-9 POU5F1/GFP EBs obtained in SCC or perfused and dialyzed STLV were enzymatically dissociated with TrypleSelect (Invitrogen) for 15 min at 37°C, washed, and resuspended in 1 mL fluorescence-activated cell sorting (FACS) buffer (2% FBS in PBS). Cells were probed for 30 min at 4°C with monoclonal r-phycoerythrin-mouse anti-human CD 56 (N-CAM) Clone B159 or r-phycoerythrin-isotype control (R&D systems, Lille, France). Stained cells were then analyzed in duplicate on a FACScalibur flowcytometer using CellQuest software (BD Biosciences, France).
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde for 10 min at room temperature before blocking and permeabilizing with 2% PBS, 0,1% bovine serum albumin, and Triton X-100. Primary antibodies were incubated overnight at 4°C in blocking buffer, including rabbit polyclonal antibodies raised against PAX6 (Covance, Eurogenetec, Angers, France), mouse monoclonal antibodies raised against OCT-4 (Chemicon, St. Quentin en Yuelines, France), N-CAM clone Eric1 (Santa Cruz Biotechnologies, Santa Cruz, CA), and class III–tubulin (Tuj1, Covance). Cells were then stained with the appropriate fluorophore-conjugated secondary antibody and 4',6-diamidino-2-phenylindole.
Statistical analyses
Statistical analysis was performed using Analyse-it General Statistics (Analyse-it Software, Ltd, Leeds, UK). Values are presented as means ± standard deviations.
A Fisher test was performed to compare variance in samples. The statistical significance level of a difference between data sets was determined using the Student t-test when the Fisher test was positive; Welch t-test was applied (indicated in the text by Welch's approximation) when it was not.
Results
The optimized perfused and dialyzed STLV consists of two loops of medium perfusion. The first cell-perfusion loop feeds the bioreactor. Protein complementation of the medium is performed at this level. The perfusate goes though the STLV chamber equipped with a 100-kDa-molecular-weight-cut-off (MWCO) semi-permeable membrane. The second loop consists of a dialysis chamber (200 mL) with an inner cylinder covered by a dialysis membrane with a 12-kDa MWCO connected to a large medium tank. The function of the dialysis chamber is to dilute the dialysate with fresh medium. It enhances waste elimination and nutrient supply by exchange between the two compartments of the dialysis chamber through the 12 kDA MWCO semi-permeable membrane. Two pumps control the cross-flow of the two perfusion loops independently to provide full medium renewal within 24 h. Because of on-line analysis, a bubble trap has been added on the first loop to avoid pump failure while retrieving medium for analysis (Fig. 1).
Effectiveness of the dialysis chamber on the maintenance of culture conditions
To validate the effectiveness of this system, we first measured physicochemical parameters over 10 days of EB culture (Table 1), namely pH, osmolarity, and partial pressure of CO2 and O2. All these parameters are indirect indicators of homeostasis, glucide, and respiratory and ionic metabolism and together reveal the mass transfer capacity of the system. The results obtained indicated a robust stability for all parameters over time in the culture chamber.
Four physicochemical parameters were measured twice a day between day 0 and day 10. Variations of the physical parameters are presented as means ± standard deviations.
Figure 2 summarizes the results of successive analyses in the culture medium of concentrations of glucose, glutamate, ammonium (NH4), and lactate.
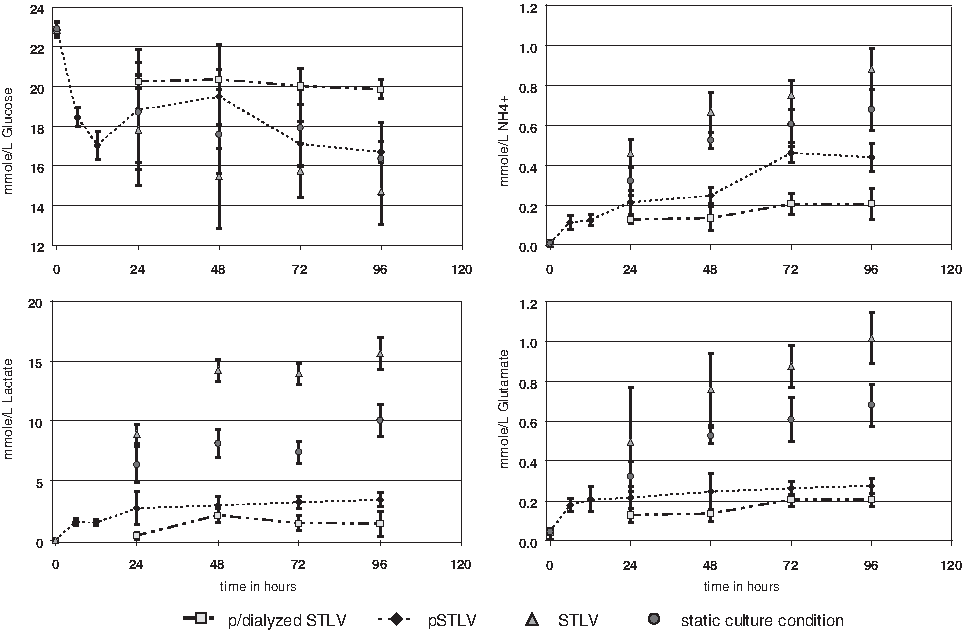
Concentrations of glucose, ammonium (NH4+), lactate, and glutamate in the culture medium between day 0 and day 4 of differentiation. Analyses were performed on perfused and dialyzed slow-turning lateral vessels (STLVs) (▪), perfused STLVs (♦), STLVs (▴), and under static culture conditions (•). The data are presented as means ± standard deviations (n = 3, 3, 6, and 6, respectively). Dotted lines indicate analyses performed by the bioanalyzer on samples directly retrieved on-line. For other results, samples were retrieved manually and analysis performed secondarily using the bioanalyzer.
Results obtained under SCCs and non-perfused STLV were similar, showing on the second day a progressive decrease in glucose concentration and a rapid parallel increase in cell waste, with lactate reaching close to 7.5 and 14mmol/L for SCC and STLV, respectively, and NH4 doubling up to 0.6 mmol/L. Daily change of half of the medium after the second day of culture under SCCs stabilized those parameters at the levels of the second day.
In sharp contrast, addition of continuous perfusion to the STLV better stabilized concentrations over time at levels compatible with cell culture. 11 Glucose concentration followed the same evolution as under SCCs and non-perfused STLV. By contrast, most parameters indicative of cell waste increased in the medium only over the first 24 h. Glutamate and lactate then remained stable for up to 4 days, at 0.2 and 4 mmol/L, respectively. NH4 doubled again within 48 h to reach a plateau at 0.4 mmol/L, Addition of a dialysis chamber to the perfused STLV systematically improved all culture parameters. Glucose concentration remained at approximately 20 mmol/L up to 4 days (i.e., ∼25% than in the non-dialyzed system). In parallel, all indicators of cell waste were half those of the non-dialyzed system. Lactate was in the mmol range at all times, and glutamate and NH4 were at approximately 0.2 mmol/L. Cell viability was similar at day 14 in perfused and in perfused and dialyzed STLV, better than in non-perfused STLV and under SCCs (Table 2).
Cell viability assays were performed using the live/dead viability cell kit from Invitrogen on day 7 and day 14 for each condition (static culture condition (SCC), slow-turning lateral vessel (STLV), perfused STLV, and perfused and dialyzed STLV) and following the manufacturer's protocol. Analyses were performed in triplicate. For the SCC and non-perfused STLV condition, the medium was changed by half every other day.
Effect of simulated microgravity on hESC differentiation
Two main features of the differentiation of hEBs were analyzed to compare results obtained using SCCs and the perfused and dialyzed STLV, namely the decrease in expression of markers of the undifferentiated stage and the increase in expression of markers of differentiation, specifically focusing on the neural lineage. For the undifferentiated stage, we chose NANOG and OCT-4, two transcription factors associated with the undifferentiated hESC stage, and the controls TDGF1 and LEFTY A that also participate specifically to this stage.12,13 Analysis of neural differentiation roughly distinguished two stages: an earlier one characterized by expression of FGF5, SIX3, SOX1, and PAX6, and later Nestin, CD56/N-CAM, MAP2, and TUJ-1.
In addition, we determined the course of expression of E-cadherin, because it has been shown to peak during the phase of aggregation and then to decrease transiently before increasing again during formation of three germ layers.14,15 This specific time course of expression was used to mark the effectiveness of induction and the completeness of differentiation.
In the analysis of 32 randomly retrieved individual hEBs, expression of NANOG and OCT-4 statistically significantly decreased more rapidly in perfused and dialyzed STLV than under SCCs at 48 and 96 h (Fig. 3A, B). This enhancement was also significantly more homogeneous in perfused and dialyzed STLV, and no undifferentiated stage patterning resulting in defective aggregation contrary to SCCs was seen (black arrows in Fig. 3B). These observations were confirmed with another cell line, SA-01 (Supplemental Fig. 1, available online at
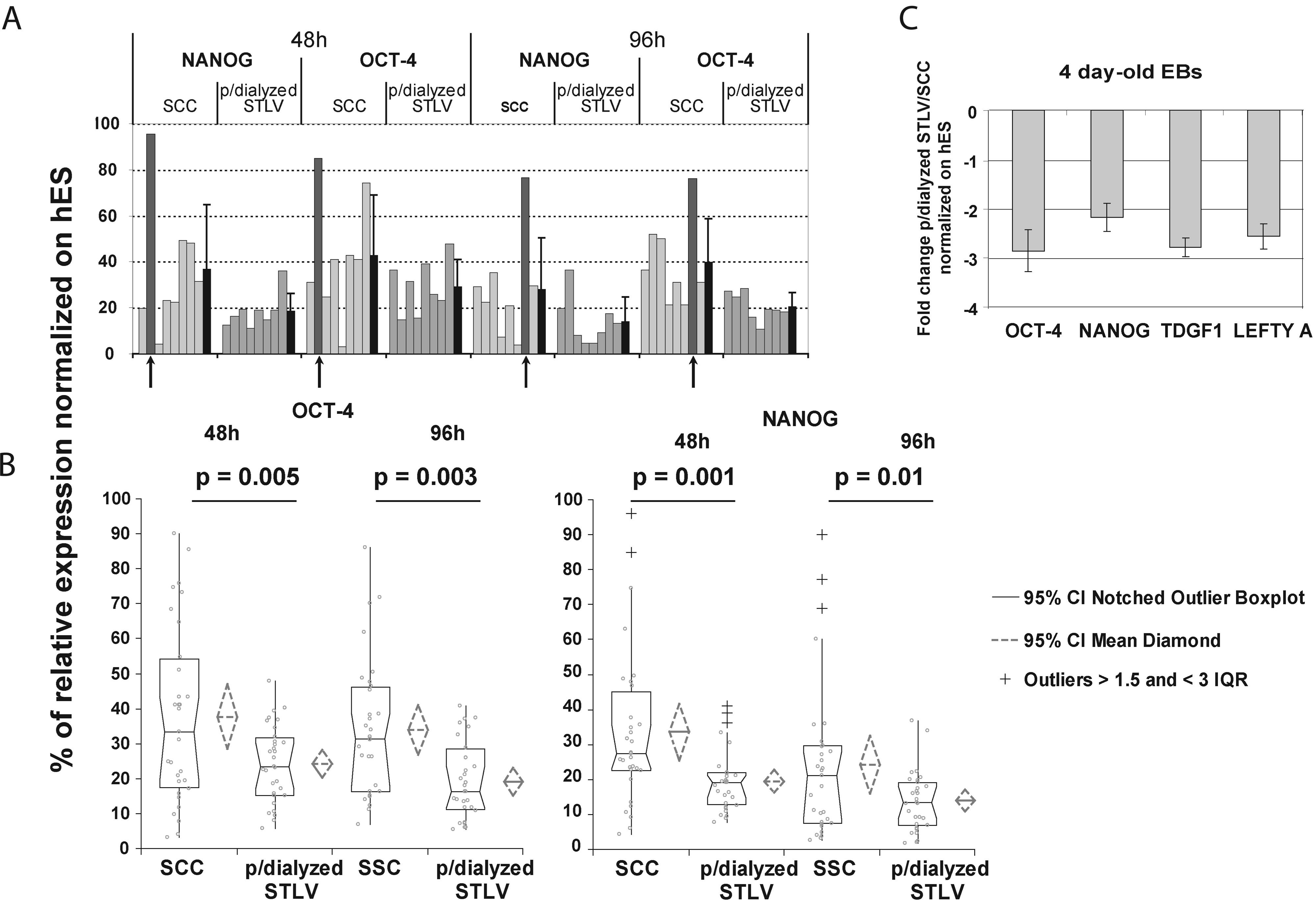
Quantitative polymerase chain reaction of relative octamer-4 and NANOG gene expression in embryoid bodies (Ebs) at 48 and 96 h (VUB-01 line). (
Comparison of mixes of 4-day-old hEBS showed that all markers of the undifferentiated stage were at least half those in perfused and dialyzed STLV than under SCCs (one-way Student t-test perfused and dialyzed STLV/ SCCs with p < 0.01, n = 3) (Fig. 3C). Those results were confirmed using FACS analysis on cells expressing GFP under the control of the promoter of OCT-4, which showed a 24-h delay in the differentiation kinetics at the earliest time-points for the perfused and dialyzed STLV conditions compared with SCCs (Fig. 4).

Progression toward three-germ-layer stage from static culture conditions (♦) and perfused and dialyzed slow-turning lateral vessel (▪) cultures in embryoid bodies (octamer (OCT)-4/green fluorescent protein (GFP) Hues-9 line). The determination of induction and progression of differentiation was performed by control of percentage of total population expressing GFP under OCT-4 full-length-promoter expression (dotted lines) and neural cell adhesion molecule (dark lines) monitoring usng flow cytometry over the first 5 days. Data are shown as means ± standard deviations (n = 3).
Analysis of differentiation markers in mixed samples of EBs at 4 and 8 days confirmed the overall acceleration of the differentiation (Fig. 5). At 4 days, E-cadherin expression in STL-derived hEBs was one-fifth that under static conditions. At 8 days, the reverse was observed, with E-cadherin expressed three times more in the former than in the latter, indicating a faster progression toward gastrulation. Early neural differentiation markers (FGF5 and SOX-1) showed a similar acceleration in the STLV conditions compared with SCCs (Fig. 5A). At that 8-day time point, expression of genes associated with later stages of neural differentiation was also 5 to 10 time more in the perfused and dialyzed STLV than under SCCs (Fig. 5B).

Characterization of embryoid bodies (Ebs) differentiated in perfused and dialyzed slow-turning lateral vessel (STLV) conditions versus static culture conditions (SCCs) after 4 days (black bars) and 8 days (grey bars) (VUB01 line). Upper panel: (
Neural differentiation of EBs produced in simulated microgravity
To analyze the effect of a rotary bioreactor on the specific neural differentiation, we compared this differentiation for hEBs derived from an H9 hESC line in the perfused and dialyzed STLV, perfused STLV, and non-perfused STLV; under SCCs; and using co-culture with stromal cells.
As shown in Figure 6A, the mean time delay to “neural rosette” formation grown was significantly shorter under all three STLV conditions than under SSCs (1 to 2 days), and all were more than 1 week shorter than after induction of stromal cells. Under STLV conditions, they were collected after only 13 to 14 days (6 days in the bioreactor and 7–8 days after plating hEBs), whereas early neural rosettes were observed only after 23 days using co-culture with MS5 feeder cells, in agreement with previous data. 10
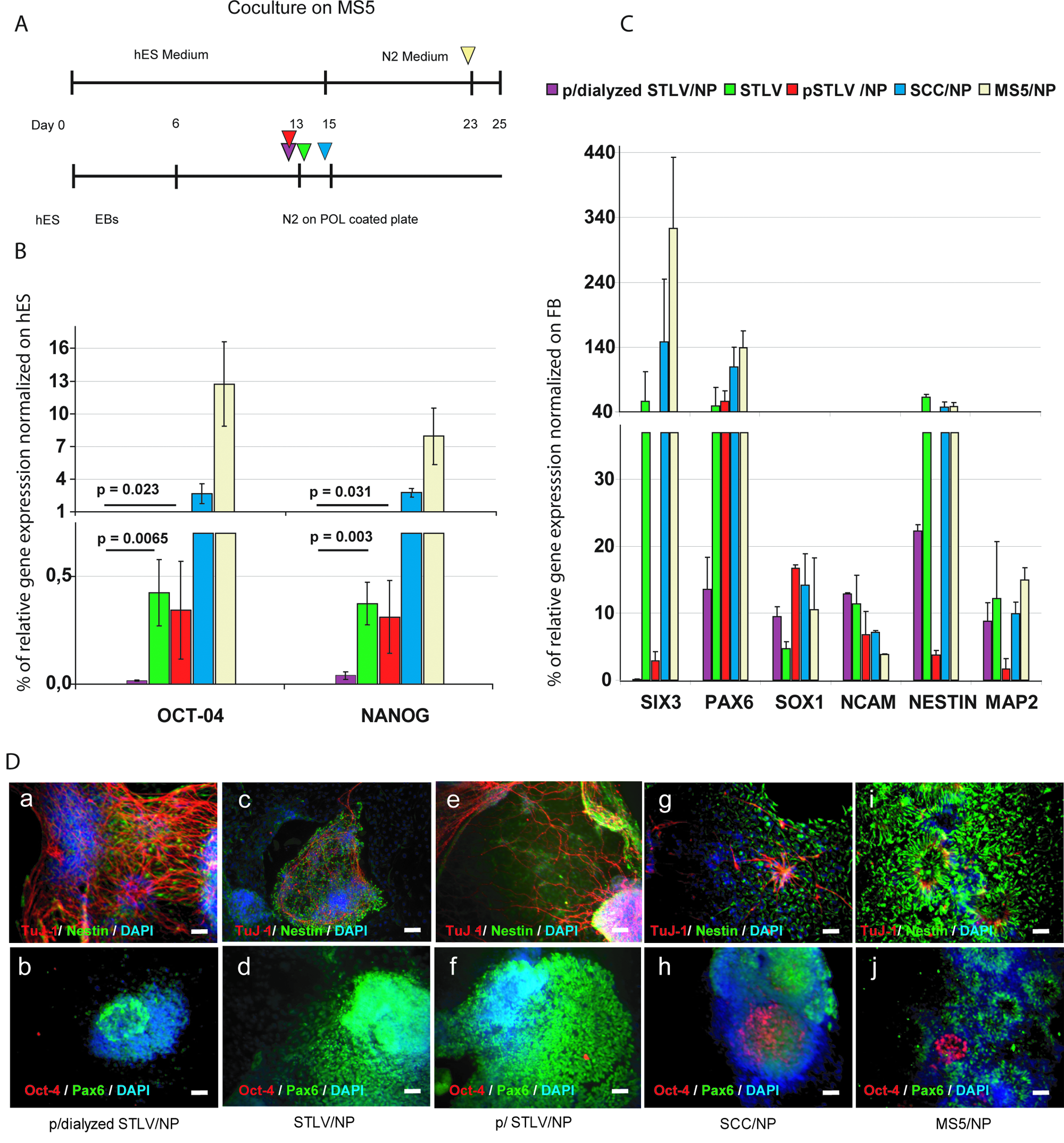
Comparison of neural progenitors (NPs) generated under perfused and dialyzed slow-turning lateral vessel (STLV), perfused STLV, non-perfused STLV, static control conditions (SCCs) and after co-culture with MS5 stromal cells (H9 line). (
Undifferentiated leftover cells in neural rosettes were analyzed using real-time PCR of OCT-4 and NANOG. The expression of these markers of the undifferentiated stage was dramatically lower STLV and perfused STLV until undetectable in the perfused and dialyzed STLV-derived rosettes, whereas both remained expressed in co-culture– and SCC-derived rosettes, at lower but significant levels to the undifferentiated stage (Fig. 6B). All others markers of differentiation increased as expected under all three conditions. Once normalized on gene expression recorded in the brain of a 21- to 41-week-old human fetus, results were not significantly different for SOX-1, N-CAM, Nestin, and MAP 2 (Fig. 6C). SIX3, a precocious and transient marker gene of neural specification, appeared down-regulated only in perfused and perfused and/dialyzed STLV conditions. The typical marker of early (rosette-associated) neural precursors, PAX6, was less expressed in in perfused and dialyzed STLV than under all other conditions, a result confirmed using immunocytochemical staining of cells in culture (Fig. 6D). In contrast, the cells expressing the marker of more-advanced neuronal progenitors TUJ-1 were more numerous in perfused and dialyzed STLV culture than under any other condition.
Discussion
The main result of this study is the demonstration that optimizing low-shear-stress STLV bioreactors through control of a continuously perfused culture medium allows for long-term production of hEBs that differentiate more rapidly and more homogeneously than under classic culture conditions. Addition of a regenerating dialysis loop permitted the further optimization of the medium. It also restricted the need for renewal of the medium.
Non-perfused STLV bioreactors have been shown to be a useful tool for bulk production of EBs derived from hESCs. 4 We have built upon those data in the search for technical improvements that would allow optimization of the culture medium content through adaptation to the continuously changing needs of the cells in the bioreactor chamber. Using a perfusion system allowed us to address this issue in three different and complementary ways. First, it allowed for continuously diluting cell waste while maintaining concentrations of nutriments, because the culture medium in the chamber was in permanent equilibrium with an external source. Second, it allowed the medium in the chamber to be changed whenever needed without altering the growth process. In bioreactors, interrupting stirring—and microgravity in STLV—leads to agglomeration of EBs and formation of aggregates that are too large for optimal growth and differentiation and thus disturb the evolution of the early stages of the gastrulation.16,17 Continuous diffusion of the medium reduces the variations of free-steam concentration, because it results in a nearly constant value at the surface of the aggregates that rotate in the bioreactor. 18 Third, a perfusion system creates a derivation of the culture medium that can then be analyzed directly, allowing for precise control of the microenvironment in which the cells are growing. It is of most importance that adequate oxygen and nutrient mass transfer are essential parameters to maintain growth and differentiation of aggregated cells in suspension, 19 and they are continuously evolving over time in bioreactors. Accordingly, cell viability was better under perfused STLV conditions, dialyzed or not, than under non-perfused STLV conditions or SCCs. Microgravity culture condition has been successfully used for organic tissue growth and differentiation,20,21 with for example, low shear stress adapted with sensitivity of the construct–liquid interface, microenvironment around the structure that include nutrient and O2 supply, and preservation of paracrine autocrine pathways. 22 The third conditions could explain maintenance of growth and differentiation.
Moreover, to avoid disturbing this rotation frequently, which must be continuous, we also added an on-line analysis system of culture. That gives us a real-time following of the medium composition without having to intervene inside the culture chamber, thus allowing us to perform real continuous hEB cultivation while limiting direct human technical intervention on cells during the culture processes.
The use of a perfusion system with the bioreactors allowed us to move a step further in the optimization of the culture medium by adapting a dialysis chamber to the STLV bioreactors comparable with those designed for other applications. 23 The value of this addition was demonstrated on all culture medium parameters studied. Our interpretation of these results is summarized in Figure 7 and points to the differential cut-off of the membranes in the bioreactor and the dialysis chambers as one key characteristic of the system. By restricting the cut-off point in the dialysis chamber to 12 kDa, we provoked a differential dilution of the elements contained in the culture medium. Exchanges with the used medium flowing out of the bioreactor chamber will be at equilibrium for all elements that can cross the membrane, provoking movement in both directions according to gradients of concentrations. Nutrients that have been used are replenished in the culture medium. Cell waste metabolites that have increased in concentration in the bioreactor chamber will cross the membrane and dilute in the dialysis chamber. Because of their size, most proteins will not cross the membrane and will remain in the culture medium, resulting in growth factors being more effective than under static conditions. 24

Left: schematic representation of the two flows of medium in the dialysis chamber, from the slow-turning lateral vessel (STLV) chamber (perfusion loop), and of fresh medium (dialysis loop). Right: predicted gradient-driven flows of key molecules of the culture medium across the semi-permeable membrane (12-kDa cut-off ).
The use of low-shear-stress bioreactors to grow hEBs has been first considered for their ability to address the issue of mass cell production. 4 We have considered a complementary issue, namely the ability of a system allowing for controlled culture conditions over time to improve the differentiation process of hEBs. Our results support this hypothesis by demonstrating that EBs grown in the perfused and dialyzed STLV bioreactors underwent differentiation in a more-homogeneous manner and, as an example of the overall ability of the system, reached a characteristic stage of neural differentiation more rapidly than under other conditions tested. Micro-gravity affects expression of numerous genes, cell-to-cell interaction, and paracrine–autocrine cell pathways. 16 The minimal mechanical stress inside the vessel is also favorable for cell aggregation and aggregate culture. 25 As demonstrated, the formation of aggregates in simulated microgravity avoided the maintenance of residual undifferentiated cells, which are believed to be implicated in teratoma formation; the ability to eliminate this risk is essential for regenerative medicine. 26
Similar to our results, Gerecht-Nir and colleagues noted that consumption of nutrients and increased concentration of cell waste metabolites characterized non-perfused STLV culture conditions. In non-perfused bioreactors, similar to what occurs under SCCs, growing cells are thus treated with culture medium, which loses optimal characteristics soon after each exchange. In perfused bioreactors, the stability of the cell culture conditions at a level much closer to the original concentrations is a likely major mechanism underlying the improved homogeneity of the differentiating EBs. Optimization of culture conditions may also be responsible for the faster speed of differentiation recorded in the present study. Indeed, under static conditions, it is likely that cells undergoing differentiation are placed in optimal conditions only for short periods of time after medium renewal. As a consequence, speed of differentiation may be cycling after each medium exchange from normal to sub-normal in static conditions. The perfused and dialyzed STLV may, in contrast, continuously preserve culture conditions closer to optimal ones, allowing molecular mechanisms of differentiation to operate more normally.
Footnotes
Acknowledgments
This study was supported in part by additional grants from the Agence Nationale de la Recherche (CSCelo), the cluster Medicen Paris Region (IngeCELL) and the European Commission (STEM-HD, FP6). The authors thank Dr Karen Sermon (AZ-VUB, Brussels) for the kind gift of VUB-01, Dr Chad Cowan of HUES-9 for the OCT-4GFP hESC line, and Bernard Prum for statistical analyses.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.