
Abstract
Bone marrow (BM) mesenchymal stem cells (MSCs) represent an interesting field of research for their in vitro properties and the in vivo therapeutic applications. In the present study, we compared the clonogenic and differentiation capacity of MSCs present in three BM-derived populations—namely, the CD105+/CD45− cells, the glycophorin A (GlycoA)−/CD45− cells, and the BM mononuclear cells (BMMCs)—by growing/expanding clones from single colony-forming unit fibroblasts (CFU-F). We also quantified the Oct-4 and Nanog mRNA in the CD105+/CD45− and GlycoA−/CD45− cells to define the fraction containing more immature MSCs. We found that basic-fibroblast growth factor (bFGF) favors the long-term survival and growth of the more immature MSCs but has no significant effect on MSC clonogenic potential. CFU-F number and clone recovery were higher in CD105+/CD45− compared to GlycoA−/CD45− (p < 0.0001 and p = 0.0364, respectively) cells or BMMCs (p < 0.0001 and p = 0.0007, respectively). The relative mRNA expression of Oct-4 and Nanog was significantly increased in CD105+/CD45− compared to GlycoA−/CD45− cells (p < 0.0001 and p < 0.0001, respectively). No significant difference was found in the immunophenotypic characteristics and differentiation potential of clones derived from all three cellular sources. These data suggest that the CD105+/CD45− BM cell fraction is enriched in immature MSCs and, accordingly, represents an appropriate source for MSC culture initiation.
Introduction
MSCs represent a minor fraction of the total nucleated cell population in the BM with an approximate frequency of 1 MSC per 5 × 103 BM mononuclear cells (BMMCs). 9 It is therefore necessary to cultivate MSCs in vitro to obtain adequate cell numbers for clinical purposes. MSC expansion, however, under culture conditions may result in alteration of the original stem cell characteristics and loss of MSC multipotentiality, and may also drive cells to senescence or even to spontaneous transformation. 10 It is therefore essential to develop improved isolation techniques to obtain fresh noncultured MSCs with intact stemness properties. MSC enrichment has been proposed by improving plastic adherence cell properties in the presence of selected batches of fetal calf serum (FCS)1,11 and also by isolating cells with positive and negative purification techniques based on a variety of cell surface antigens.12–15 Identification of the more immature fraction of MSCs is based on the expression of genes known to be associated with maintenance of MSC multipotentiality. These stemness markers include the Pit/Oct/Unc (POU) domain–containing octamer-binding protein 4 (Oct-4/POU5F1) 16 and the homeoprotein Nanog.17,18 They both represent transcriptional factors originally described in embryonic stem cells (ESCs). Specifically, they have been identified in the pluripotent cells of the inner cell mass where, in association with the Sox-2 transcription factor, they regulate the pluripotent and self-renewal potential of ESCs by controlling the expression of a variety genes during development.19,20 Downregulation of these three transcription factors has been associated with induction of differentiation process. 18 Interestingly, Oct-4 and Nanog have been also identified in human MSCs possibly acting through similar regulatory mechanisms as in ESCs. 21
The aim of the current study was to compare the properties of MSCs in three different BM cell populations, namely, the CD105+/CD45− cells, the glycophorin A (GlycoA)−/CD45− cells, and the BMMCs. The type III receptor of the transforming growth factor (TGF)-β receptor family cell-surface antigen CD105 (endoglin) was selected on the basis of its wide expression on MSCs. 22 Negative selection for GlycoA and the common leukocyte antigen CD45 aimed to exclude hematopoietic cells, whereas the BMMCs were also assessed as the most common population for initiating MSC cultures. 23 We specifically compared MSC clonogenic capacity and differentiation potential in the three populations and also quantified the expression of the Oct-4 and Nanog stemness markers in the CD105+/CD45− and GlycoA−/CD45− cells to define the cell fraction containing the more immature MSC reserves.
Materials and Methods
Isolation of BM subpopulations
BM aspirates were obtained from posterior iliac crest from healthy volunteers (n = 10) after informed consent according to the Helsinki declaration, and the BMMCs were isolated following centrifugation on Histopaque-1077 (Sigma-Aldrich, St. Louis, MO). Using an immunomagnetic cell separation system (Miltenyi Biotec, Bergisch Gladbach, Germany), we subsequently isolated the CD105+/CD45− and GlycoA−/CD45− cell subpopulations according to the manufacturer's instructions. More precisely, isolated BMMCs were labeled for CD45, and the negatively selected cells were equally divided and further labeled for (a) CD105 and (b) GlycoA according to the manufacturer's instructions. Positive selection was then used for CD105, and negative for GlycoA. All three study cell populations—the CD105+/CD45−, GlycoA−/CD45− cells, and BMMCs—were expanded in vitro in Dulbecco's modified Eagle's medium–low glucose (DMEM-LG; Gibco Invitrogen, Paisley, Scotland)/10% FCS (Hyclone, Logan, UT)/100 IU per mL penicillin–streptomycin (MSC medium) as previously described, 9 either with or without 1 ng/mL bFGF (R&D Systems, Minneapolis, MN).
Clonogenic potential of BM subpopulations
A colony-forming unit fibroblast (CFU-F) assay was used to evaluate the clonogenic potential of MSCs within the three cell populations. In brief, day 0 cells were seeded at three different concentrations in 60-mm plates and cultured for 14 days in MSC medium in the presence or absence of 1 ng/mL bFGF. A CFU-F was defined as a discrete colony containing 50 or more cells. CFU-F frequency was estimated using a linear regression analysis from the three different initial cell concentrations. CFU-F size (expressed in mm) was measured using a graduated ruler. 24
Generation of MSC clones
It has been reported that each CFU-F derives from a single MSC. 25 We have therefore generated clones from CFU-F to indirectly investigate the properties of MSCs present in the initial cell subpopulations. In brief, day 0 cells were initially seeded in 60-mm plates at a concentration of 5 × 104 CD105+/CD45− cells/plate, 5 × 104 GlycoA−/CD45− cells/plate, and 10 × 104 BMMCs/plate. Following 14-day culture in MSC medium supplemented with 1 ng/mL bFGF, generated colonies, presumably arising from single MSCs, were individually picked using cloning rings, and plated in separate wells of 24-well plates. Subsequently, trypsinized cells were expanded to 25-cm2 flasks and finally to 75-cm2 flasks.
Phenotypic characteristics of MSC clones
Trypsinized cells derived from MSC clones were phenotypically characterized by flow cytometry, using monoclonal antibodies against CD29 (4B4; Cyto-Stat/Beckman-Coulter, Fullerton, CA), CD44 (J173; Immunotech/Coulter, Marseille, France), CD73 (AD2; Becton Dickinson-Pharmingen, San Diego, CA), CD90 (F15.42; Immunotech/Coulter), CD105 (SN6; Caltag, Burlingame, CA), CD146 (P1H12; Becton Dickinson-Pharmingen), CD45 (IMMU19.2; Immunotech/ Coulter), CD14 (RMO52; Immunotech/Coulter), and CD34 (QBend10; Beckman-Coulter).
Differentiation potential of MSC clones
The differentiation potential of MSCs in the initial cell populations was evaluated by exploring the capacity of the clones to generate cells of the adipogenic, chondrogenic, and osteogenic lineages as previously described. 9
Adipogenic differentiation was induced in MSC medium supplemented with 10% FCS/0.5 mM 1-methyl-3-butylisoxanthine/1 μM dexamethasone/0.2 μM indomethacin/10 μg/mL insulin. Differentiation was assessed by Oil red O stain. 11
Osteogenic differentiation was induced in MSC medium supplemented with 0.1 μM dexamethasone/0.15 mM ascorbate-2-phosphate/3 mM NaH2PO4. Differentiation was assessed by alkaline phosphatase (ALP)/Von Kossa stain. 11 .
For chondrogenic induction, cells were pelleted in 15 mL tubes and cultured in DMEM–high glucose (Gibco Invitrogen), supplemented with 6.25 μg/mL insulin/6.25 μg/mL transferrin/1.33 μg/mL linoleic acid/1.25 mg/mL bovine serum albumin/1 mM sodium pyruvate/0.17 mM ascorbate-2-phosphate/0.1 μM dexamethasone/0.35 mM L-proline/6.25 ng/mL selenous acid/0.01 μg/mL TGF-β3 (R&D Systems). 26 Differentiation was identified with Alcian blue and Masson's trichrome stains. 11 All reagents were purchased from Sigma (St. Louis, MO) unless otherwise indicated.
Total RNA isolated from differentiated MSC clones (RNeasy mini kit; QIAGEN, Hilden, Germany) was reverse transcribed (SUPERSCRIPT II; Gibco) and amplified by PCR for the evaluation of specific, differentiation-associated gene expression: adipose fatty acid–binding protein (aP2) and peroxisome proliferator activated receptor-γ (PPARG) for adipocytes, ALP and runt-related transcription factor 2 (RUNX2) for osteocytes, and collagen type II (COL2A1) and aggrecan (AGC1) for chondrocytes. Products were normalized according to β2-microglobulin (β2m) expression, using the ImageJ densitometry analysis system. Primer sequences are listed in Table 1.
Analysis has been performed by RT-PCR in clones derived from single CFU-Fs following a differentiation procedure toward the adipogenic (aP2 and PPARG), osteogenic (ALP and RUNX2), and chondrogenic (COL2A1 and AGC1) pathways.
aP2, adipose fatty acid–binding protein; PPARG, peroxisome proliferator activated receptor-γ; ALP, alkaline phosphatase; RUNX2, runt-related transcription factor 2; COL2A1, collagen type II; AGC1, aggrecan; β2m, β2-microglobulin.
Real-time quantitative RT-PCR
Total RNA isolated from day 0 CD105+/CD45− and GlycoA−/CD45− cell subpopulations was reverse transcribed. Platinum®qPCR SuperMix-UDG (Invitrogen, Paisley, Scotland) and TaqMan® Gene Expression Assays (Applied Biosystems, Foster City, CA) were used for Nanog, Oct-4, and GAPDH identification. Measurements were performed using the ABI Prism 7000 Sequence Detection System software (Applied Biosystems). Results were normalized by dividing the mean quantity of target gene by mean quantity of GAPDH.
Statistical analysis
Data were analyzed using the Student's t-test and the one-way analysis of variance (ANOVA). Grouped data are expressed as mean ± 1 SD.
Results
bFGF favors the expansion of MSCs
Final clone expansion was not reached by all originally generated clones. By defining as clone recovery, the number of finally expanded clones divided by the number of initially plated clones, we found that bFGF favors clone recovery independently of the initial MSC source. Specifically, in the presence of bFGF, clone recovery increased significantly compared to the untreated cultures in all three populations, namely, the BMMCs, the CD105+/CD45− cells, and GlycoA−/CD45− cells (38.34% ± 19.77% vs. 21.99% ± 10.04%, 70.31% ± 14.81% vs. 35.71% ± 7.53%, and 49.89% ± 24.41% vs. 31.51% ± 1.58%, respectively; p = 0.0315, p < 0.0001, and p =0.0288, respectively) (Fig. 1A). We have therefore used bFGF in all clone-expanding experiments. Interestingly, although the presence of bFGF did not increase significantly the CFU-F number obtained by the three populations at day 0 in comparison to the untreated cultures (p = 0.4764 for BMMCs, p = 0.4850 for CD105+/CD45−, and p = 0.2772 for GlycoA−/CD45− cells), it dramatically increased the colony size. Specifically, the mean size of CFU-F derived from BMMCs was 6.24 mm ± 1.95 mm in cultures treated with bFGF and 3.64 mm ± 1.44 mm in untreated cultures (p < 0.0001) (results correspond to the mean size of total CFU-F number derived from BMMCs of four healthy controls in the presence or absence of bFGF). A typical example is shown in Figure 1B, whereas similar results were obtained from cultures derived from GlycoA−/CD45− and CD105+/CD45− cells (data not shown). Based on the aforementioned data, we might suggest that bFGF favors the long-term survival and growth of an immature fraction of MSCs but has not any significant effect on the clonogenic potential of MSCs.
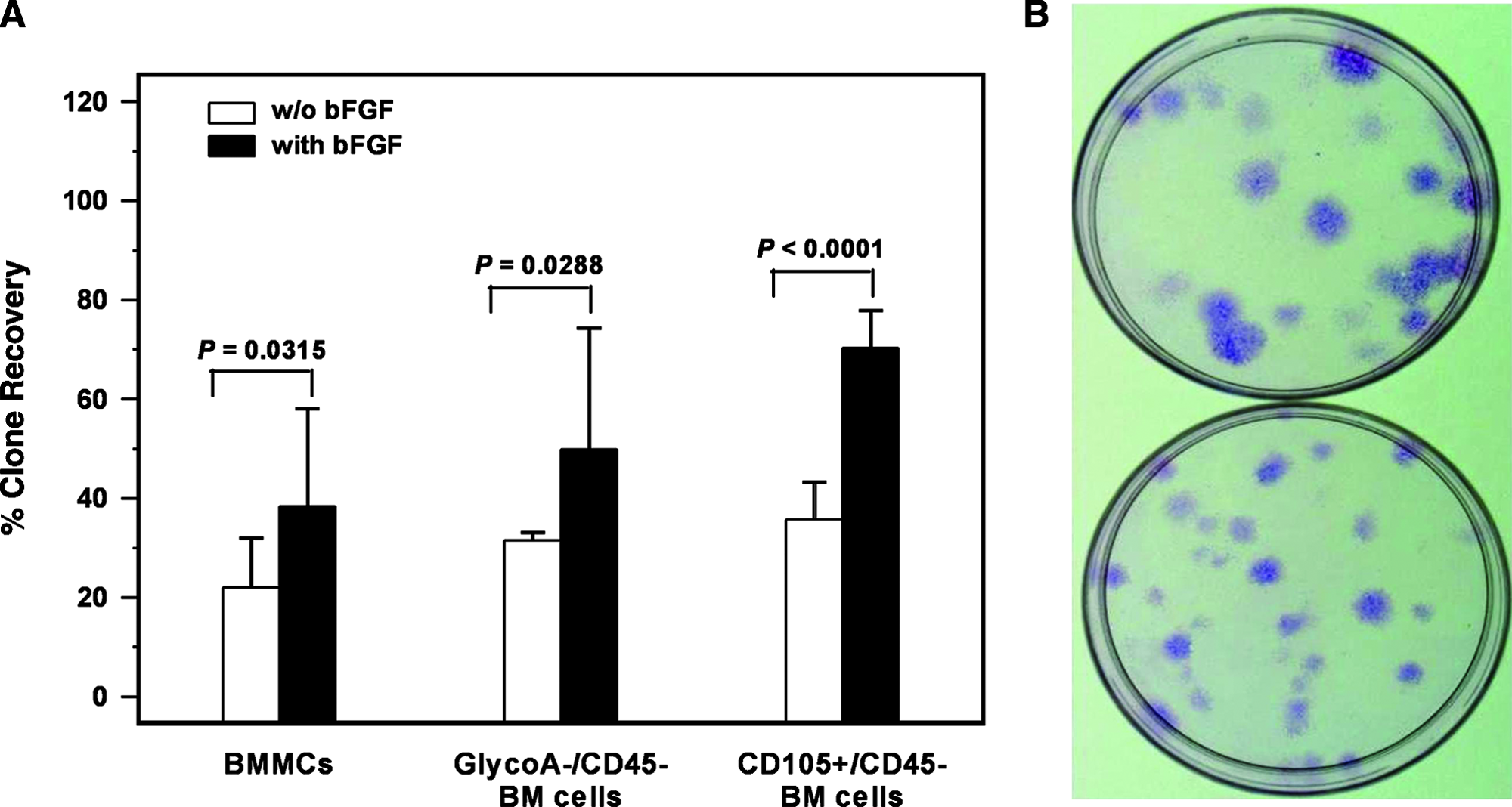
The effect bFGF in CFU-F colony growth and clone recovery. (
CD105+/CD45− cells give higher CFU-F number compared to GlycoA−/CD45− or BMMCs
The immunomagnetically sorted CD105+/CD45− cells (1.46% ± 0.87% of the original BMMC population) and GlycoA−/CD45− cells (9.63% ± 5.00% of the original BMMC population) were further evaluated for their clonogenic potential. In the standard CFU-F assay, the colony number obtained by CD105+/CD45− cells (152.41 ± 66.70 CFU-F/105 cells) was significantly higher compared to GlycoA−/CD45− cells (30.29 ± 21.99 CFU-F/105 cells) or BMMCs (12.55 ± 12.45 CFU-F/105 cells) (p < 0.0001 and p < 0.0001, respectively) (Fig. 2). Given that each CFU-F derives from a single MSC, these data suggest that the CD105+/CD45− cell fraction is enriched in MSCs and accordingly represents a more appropriate source for MSC culture initiation compared to GlycoA−/CD45− cells or BMMCs. As anticipated, CFU-F frequency differed significantly between the GlycoA−/CD45− cells and BMMCs (p = 0.0395), as the latter cell population is highly contaminated by hemopoietic cells.
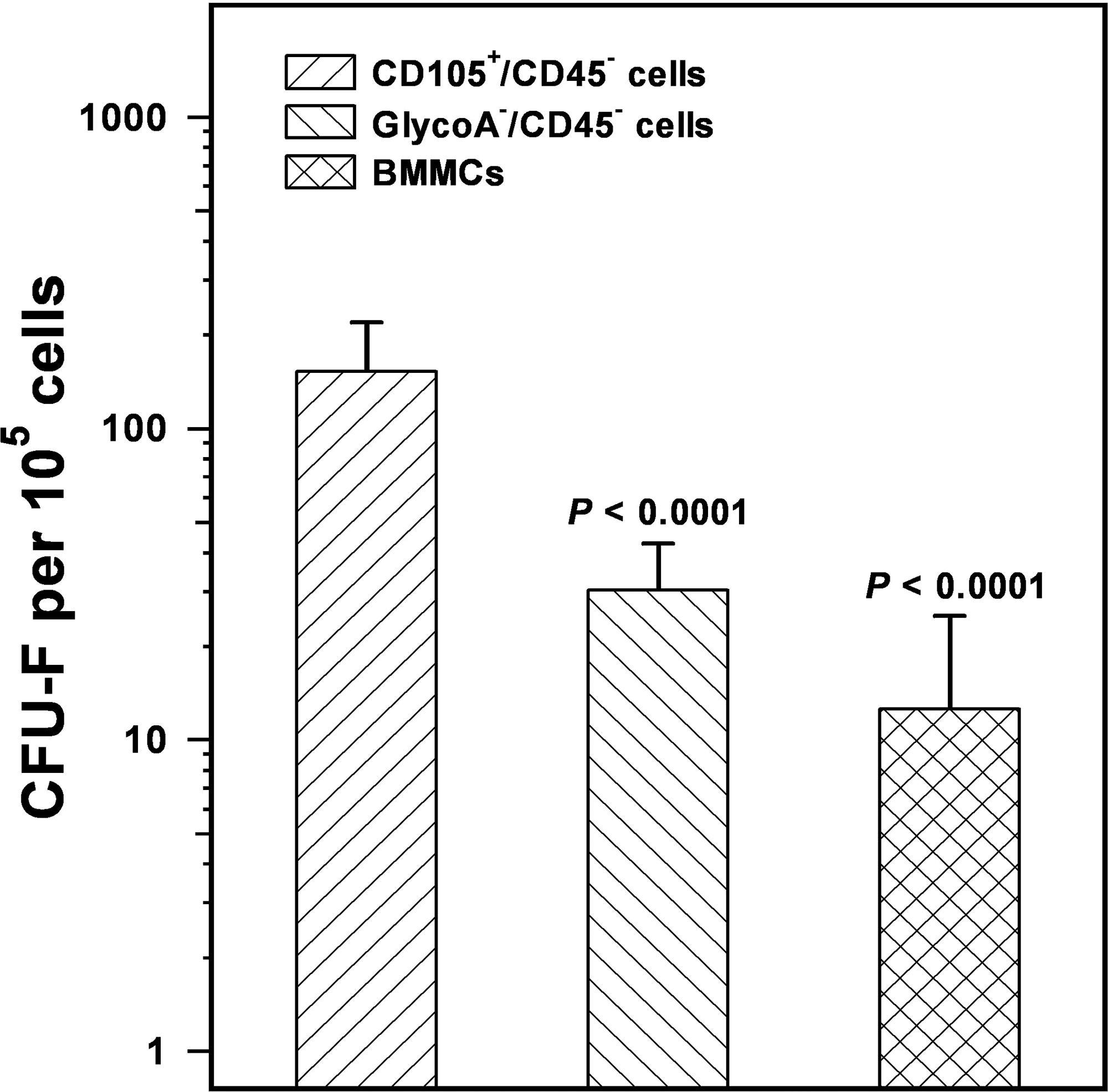
Frequency of CFU-F in the CD105+/CD45− cells, GlycoA−/CD45− cells, and BMMCs. The bars represent the mean ± SD frequency of CFU-F obtained by the three study populations using a clonogenic assay. Statistical analysis has been performed by the Student's t-test, and p-values indicate the comparison with the CD105+/CD45− cells.
Day 0 cells from the three isolated subpopulations were plated at very low concentrations (4000 BMMCs/cm2 and 2000 cells/cm2 for CD105+/CD45− and GlycoA−/CD45−), and after 14 days of culture, clones were generated and further expanded. Clone recovery by CD105+/CD45− cells (70.31% ±14.81%) was statistically significantly higher compared to the respective from GlycoA−/CD45− (49.89% ±24.41%; p =0.0364) cells or BMMCs (38.34% ± 19.77%, p = 0.0007). No statistically significant difference was documented in clone recovery between the GlycoA−/CD45− cells and BMMCs (p = 0.2605). These data suggest that the CD105+/CD45− cells are enriched to a more immature MSC fraction, capable of prolonged cell divisions, compared to the GlycoA−/CD45− cells or BMMCs.
CD105+/CD45− cells highly express stemness markers
To investigate further the hypothesis that the CD105+/CD45− cell fraction is enriched in immature MSCs, we quantified the mRNA expression of the stemness markers Oct-4 and Nanog by means of real-time RT-PCR. Because the BMMC fraction contains hemopoietic cells, mRNA expression was compared with the GlycoA−/CD45− cell population, which is also enriched in MSCs. The relative expression of Oct-4 and Nanog mRNA was statistically significantly increased in CD105+/CD45− cells (13.71 ± 0.05 and 75.28 ±2.75, respectively) compared to GlycoA−/CD45− cells (0.90 ± 0.07 and 1.67 ± 0.23, respectively) (p < 0.0001 and p < 0.0001, respectively), suggesting a 15-fold and 45-fold difference, respectively (Fig. 3). These data corroborate further the assumption that the CD105+/CD45− cell fraction contains an immature MSC population.
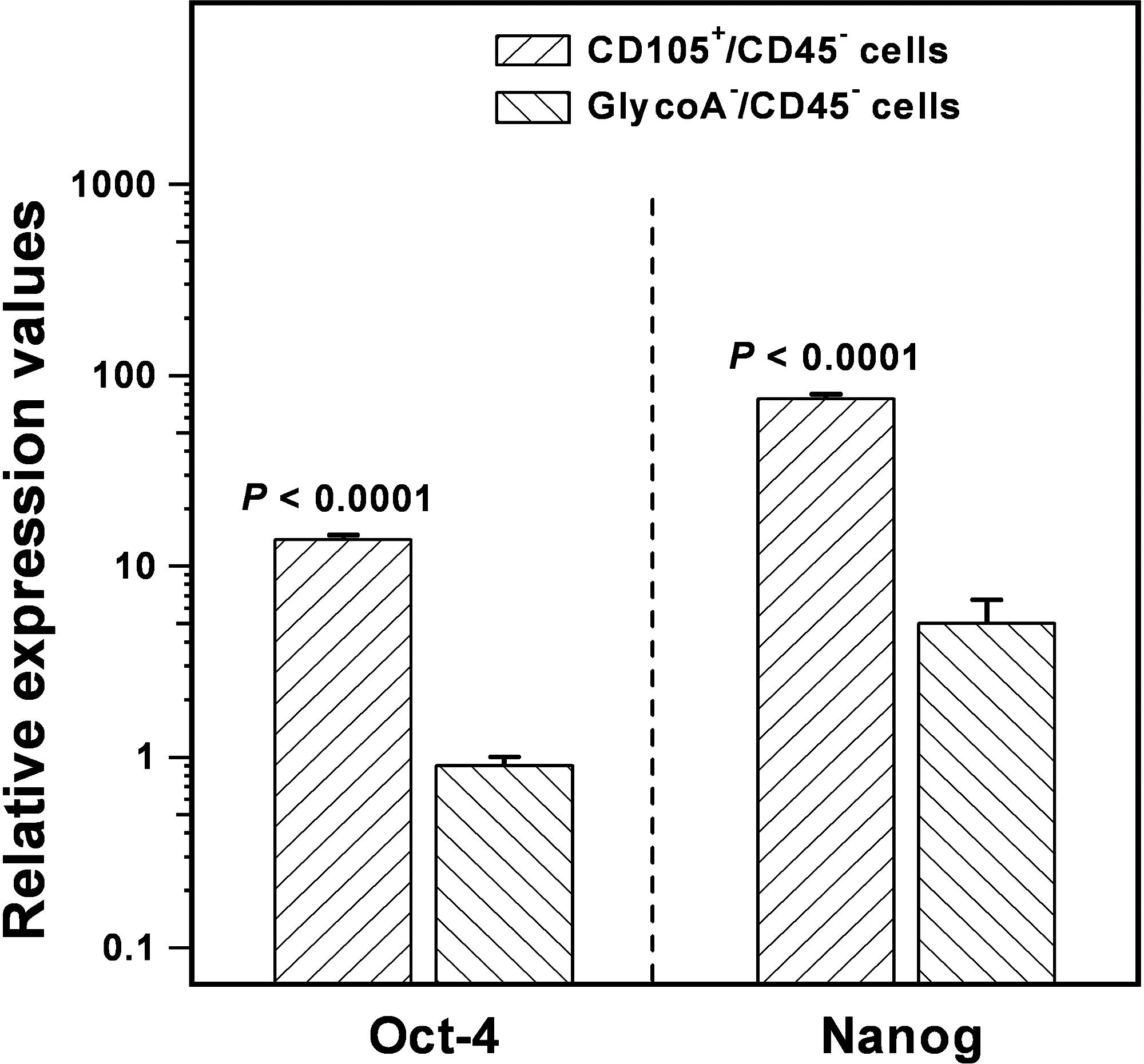
Expression of Oct-4 and Nanog genes in immunomagnetically sorted CD105+/CD45− and GlycoA−/CD45− cells by means of real-time RT-PCR. Results are normalized to GAPDH expression, as the quotient of mean quantity of target gene to mean quantity of reference gene. Statistical comparison was performed by Student's t-test, and p-values are indicated.
Differentiation potential of MSC clones
We investigated the differentiation capacity of the expanded clones toward the adipogenic, osteogenic, and chondrogenic lineages to indirectly evaluate the differentiation potential of MSCs in the CD105+/CD45− cells, GlycoA−/CD45− cells, and the BMMCs. Prior to induction of differentiation, each clone was characterized by means of flow cytometry for the expression of the anticipated MSC phenotype. In all cases, cells were positive for the MSC markers CD29, CD44, CD73, CD90, CD105, and CD146 and negative for the hemopoietic markers CD45, CD14, and CD34 (Fig. 4). We found that MSC clones derived from the three initial cellular sources display similar potential to differentiate toward the adipogenic, osteogenic, and chondrogenic pathways. This was demonstrated by the respective histochemical stains and quantified by the mean relative mRNA expression of aP2 and PPARG (adipocytes), ALP and RUNX2 (osteocytes), and COL2A1 and AGC1 (chondrocytes) that did not differ significantly among the differentiated cells derived from the three initial cell subpopulations (p = 0.896, p = 0.668, p =0.086, p = 0.827, p = 0.695, and p = 0.998, respectively) (Fig. 4).

Immunophenotypic characteristics and in vitro differentiation potential of MSC clones. Representative plots (panel
Discussion
BM-derived MSCs display the capacity to differentiate toward a variety of cells of the connective tissue lineages and have therefore attracted a great deal of interest for potential therapeutic application in tissue repair and regeneration. Because of their low frequency in the BM and the lack of cell-specific markers, MSC isolation and enrichment requires cell culture techniques. Previous studies, however, have shown that culture expansion, although highly efficient, may alter the original in vivo properties of MSCs.27,28 Accordingly, determination of an initial BM cell population enriched in MSCs for culture expansion, standardization of the culture conditions and growth promoting factors, and identification of markers for detection, selection, and testing of MSC subpopulations are of particular importance.
In the current study, we compared the properties of MSCs derived from three different BM cell populations, namely, the BMMCs, representing a common cellular source for initiation of MSC cultures, and the CD105+/CD45− and GlycoA−/CD45− cells, representing fractions enriched in MSCs. We initially evaluated the effect of bFGF in the recovery/growth promotion of MSCs. We next compared the clonogenic and differentiation potential of MSCs from the three sources and quantified the expression of stemness markers Oct-4 and Nanog in the enriched cell subpopulations to identify the fraction containing the more immature MSCs.
Our data have shown that bFGF does not exert a significant effect on the clonogenic potential of MSCs; however, the cytokine probably favors the survival and growth of an immature MSC fraction as was indicated by the CFU-F size and clone recovery. These data are in accordance with previous studies demonstrating that the addition of bFGF in MSC cultures is associated with the selection of pluripotent cells with increased telomere length. 29 Based on these data, we postulate that bFGF might extend the in vitro maintenance of MSCs in culture conditions.
To investigate the properties of MSCs in the three original cell populations, we generated and expanded MSC clones from single CFU-Fs and studied their characteristics in terms of immunophenotype, viability, and differentiation potential. Clones from all three origins displayed the anticipated MSC phenotype; that is, they were positive for CD29, CD44, CD73, CD90, CD105, and CD146 and negative for CD45, CD14, and CD34. Clones displayed also similar differentiation potential toward the adipogenic, osteogenic, and chondrogenic lineages, irrespective of the initial cellular source, as was shown by the histochemical and lineage-specific gene expression profile.
As anticipated, CFU-F recovery by MSCs at day 0 was reduced in the BMMCs compared to CD105+/CD45− and GlycoA−/CD45− cells that represent MSC-enriched BM subpopulations. Clone recovery, however, although lower in MSCs originated from the BMMC fraction, was statistically different when compared with the CD105+/CD45− cells but not with the GlycoA−/CD45− cells. These data suggest that the CD105+/CD45− cells contain a more immature fraction of MSCs. To investigate further this hypothesis, we evaluated the relative mRNA expression of the Oct-4 and Nanog stemness markers in the two MSC-enriched BM cell populations. These transcription factors, originally described in pluripotent ESCs,16,17 display a prominent role in the induction/maintenance of multipotentiality in adult stem cells, including MSCs,21,30 through similar regulatory mechanisms. Oct-4 and Nanog mRNA were markedly expressed in the immunomagnetically sorted CD105+/CD45− compared to GlycoA−/CD45− cells, corroborating further the more immature MSC content of the former cell subpopulation.
A limited number of studies using CD105+-enriched MSC population have been reported so far. In the original study of Majumdar et al., 14 the CD105+ cells, isolated with a dual antibody or single antibody procedure, were compared to BMMCs. The finding that the CD105+ cell yield was 3% ± 0.9% and 1.76% ± 0.33%, respectively, is in accordance with our findings. The CFU-F recovery using the two procedures was ninefold and threefold higher, respectively, compared to the BMMCs, a finding that is also in accordance with our data. Finally, the investigators showed that CD105+ cells were able to differentiate in vitro toward the chondrogenic lineage. In a more recent study by Aslan et al., 15 fresh CD105+ cells were engrafted and differentiated into cartilage and bone in vivo.
In conclusion, we have used three different BM cell populations to initiate MSC cultures. We have shown that all three isolation approaches—namely, the BMMCs, CD105+/CD45− cells, and GlycoA−/CD45− cells—may provide MSCs with similar differentiation capacity toward the adipogenic, osteogenic, and chondrogenic lineages. The CD105+/CD45− cell fraction, however, is enriched in immature MSCs and, accordingly, represents a better source for MSC studies compared to BMMCs or GlycoA−/CD45− cells as they will probably display a survival and proliferative advantage when used for clinical purposes.
Footnotes
Acknowledgments
This study was supported by the 6th European Union Framework Program (FP6, GENOSTEM #503161). The authors wish to thank Mrs. Athina Damianaki (University of Crete, Greece) for her technical assistance.