
Abstract
A novel nerve guide tube (poly (L-lactic) acid (PLLA)/ polyglycolic acid (PGA)-c-tube) capable of repairing long peripheral nerve injuries in a canine model has been developed. The tube was created by braiding together PLLA and PGA and then coating it with collagen. PLLA was newly added to the formulation to achieve higher sustainability. The tube was compared with a PGA-collagen tube in clinical use since 2002 having the same structure with a collagen coating but composed of PGA alone (PGA-c-tube). When tested for repair of a 40-mm gap in the left peroneal nerve, using PLLA/PGA-c-tube (n = 15), PGA-c-tube (n = 15), and a negative control group where the cut stump was capped using a silicone cap (n = 15), the lumen structure essential for securing the space for nerve regeneration was maintained in PLLA/PGA-c-tube for over 12 months with a higher number of axons both within the tube and at the distal nerve end. Electrophysiological evaluation revealed that the amplitude of compound muscle action potentials and sensory nerve action potentials after nerve regeneration with PLLA/PGA-c-tube were significantly higher. When assessed using magnetic resonance imaging (MRI), the volume of the tibialis anterior (TA) muscle in dogs that had undergone nerve repair using PLLA/PGA-c-tube was approximately 80% that of the positive control at 12 months. Functional analysis conducted by assessing the ankle angle revealed faster recovery in the PLLA/PGA-c-tube group. Better regeneration was achieved using a PLLA/PGA-c-tube that contains the slowly decomposing fiber material, PLLA. This indicates potential for repair of even longer nerve gaps or defects located near joints, and also clinical application.
Introduction
When a nerve gap is so long that end-to-end tension-free suturing becomes difficult, employment of another method is necessary to bridge the gap so that the fibers can cross over to reach the distal nerve stump. The most widely used technique is autologous nerve grafting. The use of autologous nerve grafts has yielded many successful results,5–7 although it also has certain disadvantages, such as the need for a second surgical procedure, loss of donor nerve function, the limited supply of donor nerves, and dimensional mismatch between the impaired nerve and the graft.8–10 In addition, because the donor nerve is routinely obtained from a purely cutaneous source to minimize the resulting deficit, repaired motor axons must by necessity grow under less than favorable conditions.11,12 These disadvantages have led to the development of several alternative techniques, such as those utilizing artificial nerve guides.13,14
Likewise, for repair of nerves, biodegradable artificial nerve tubes such as the polyglycolic acid (PGA) hollow tube (Neurotube; Neuroregen, LLC, Bel Air, MD) 15 or collagen hollow tube (Neura Gen; Integra Life Sciences, Plainsboro, NJ) 16 are commercially available. However, their use is indicated only for sensory nerve defects with gaps of less than 30 mm. These tubes made of thin-walled PGA or collagen do not have a supporting structure within them that provides sufficient mechanical strength for reconstruction of longer nerve gaps. 17
Nakamura et al. (2004) have developed a PGA-collagen tube that uses a collagen scaffold as the support structure (PGA-c-tube), and shown that it was able to allow regeneration across a short gap of approximately 15 mm with significantly higher levels of reinnervation than when using autografts in a canine nerve defect model. This PGA-c-tube has been in clinical use since 2002 for human patients with peripheral nerve injuries, and some promising results have been reported. 18 However, there are still several problems to overcome. First, this tube is not designed for nerve defects in certain locations, such as those near joints (i.e., the elbow, hip, or knee). Considering its material characteristics, it would probably not withstand the outside pressure caused by bending and stretching of muscles in such locations. Second, it cannot be used to reconstruct longer nerve defects such as obstetrical brachial plexus paralysis caused by complications sustained at birth. Further, we have observed that PGA and collagen degrade rapidly in vivo. 19
To overcome these problems, we have developed a stronger biodegradable nerve tube by braiding together poly (L-lactic) acid (PLLA) and PGA with collagen (PLLA/PGA-c-tube), which allows the lumen structure to be preserved for a longer period than in the case of the PGA-c-tube, thus allowing regeneration across longer nerve gaps and recovery of nerves located around joints. PLLA has long been used clinically for internal fixation of bone fractures. These screws fix the fractured section in place for over 4 months to allow rigid bone union, and the screws themselves are reabsorbed in 8 months. 20 This slowly decomposing fiber, PLLA, was braided together with a rapidly decomposing fiber, PGA, to create the PLLA/PGA-c-tube, and this allowed maintenance of the lumen structure for over 24 weeks after implantation into the back of rabbits, whereas the lumen structure of the PGA-c-tube was lost by 12 weeks (unpublished observation). Both the PGA-c-tube and PLLA/PGA-c-tube had a coating of collagen, which is known to have three major roles as a nerve insertion. One is sealing: by coating the outside wall repeatedly, neurotrophic factors or Schwann cells originating from both of the nerve stumps will be retained within the tube lumen. The second role is mechanical reinforcement: the amorphous collagen within and outside the tube maintains the initial rigidity of the tube for 2 weeks after immersion in PBS. 19 The third role is induction of axonal elongation: collagen forms a membranous structure. Placement of the collagen scaffold with a piled thin-film structure inside the nerve conduit continues to provide an appropriate environment for damaged axons, enabling regenerating axons to reach the distal nerve stump. 21 For longer nerve defects, a resorbable guide achieves better levels of regeneration than a nonbiodegradable one. 22 Among the few previous reports on the use of biodegradable nerve tubes for repair of longer nerve defects in canine models, none have provided details of functional recovery. 23
The aim of this study was to evaluate the use of PLLA/PGA-c-tube for reconstruction of a long nerve gap (approximately 40 mm) in canine models. Histological and morphological assessment using light microscopy, immunohistochemistry, and transmission electron microscopy was performed to assess the speed and quality of nerve regeneration at 3, 6, and 12 months after surgery. Functional recovery was evaluated using electrophysiology, determination of the volume of the tibialis anterior (TA) muscle using magnetic resonance imaging (MRI), and walking analysis using a high-speed camera.
Materials and Methods
Preparation of nerve conduit
The nerve conduit was prepared by braiding together PLLA and PGA in one-to-one ratio. The PLLA fiber was a monofilament (average molecular weight, 130,000; crystallinity, 50–60%; diameter, 200 μm), and the PGA fiber was a multifilament consisting of 26 PGA monofilaments (DEGRIX™; average molecular weight, 150,000; crystallinity, 56%; diameter, 17.4 μm; Mitsui Fine Chemical, Tokyo, Japan). The PLLA and PGA fibers were placed in a 48 spindle tubular braiding machine (Kokubun, Shizuoka, Japan). Each spindle held one PLLA fiber or four PGA fibers. A teflon tube with an outer diameter of 4 mm (equivalent to the inner diameter of the nerve guide tube) was inserted as a mandrel at the center of the spindle, where the fibers converged to braid. One end of the fibers was held at the top of the mandrel and pulled upward. The other end of the fibers from the spindles was then pulled to the center of the braiding machine, with half of the spindles circulating clockwise around the mandrel and the remaining spindles circulating anticlockwise. After braiding, the tube was cut to the required length with a heated blade to prevent the fibers from unwinding. It was then coated with amorphous collagen layers by repeatedly dipping into a 1% v/w collagen hydrochloride solution and then allowing it to dry. This coating process was repeated 20 times. 19 The inner side of this tube was then filled with a collagen scaffold with a piled thin film structure to induce nerve regeneration (Fig. 1B, D). 21 All collagen used in this study was atelocollagen that had been extracted from young porcine skin by enzyme treatment (Nippon Meat Packers, Ibaraki, Japan), and consisted of mainly type I collagen (70–80%), the rest being mainly type III collagen. The collagen had low antigenicity as a result of removal of the telopeptides from both ends of the collagen molecules 27 and was subjected to a virus check before use. To adjust the rate of bioabsorption, PLLA/PGA-c-tubes were subjected to dehydrothermal (140°C, 24 h) treatment in vacuo, which resulted in cross linking of the collagen molecules. The final length of the tubes was 45 mm, with an inner diameter of 4 mm and a tube wall thickness of 5.0 × 10−5 m (Fig. 1A, C). The tube was sterilized with ethylene oxide gas before surgery.

(
Animals and surgical procedure
Forty-five adult beagle dogs weighing 9.0–11.0 kg were used. The dogs were anesthetized by intramuscular injection of ketamine hydrochloride (15 mg/kg) and xyladine (7 mg/kg). After intratracheal intubation, mechanical ventilation was provided (50% oxygen and 50% nitrous oxide mixed with 1% halothane) to allow the maintenance of anesthesia by inhalation during surgery. A 10-cm skin incision was then made in the left lateral knee along the length of the thigh 1 cm outside the condylus lateralis. The tractus ilio tibialis was cut 5 cm longitudinally, and the left peroneal nerve was exposed. The nerve was then transected with a surgical scalpel at a point 50 mm from its entry point into the TA muscle, and a segment of the distal stump was resected, leaving a gap of approximately 40 mm due to retraction of the nerve stumps. In the PLLA/PGA-c-tube group, the cut nerve was re-established by interposing the 44-mm PLLA/PGA-c-tube (n = 15) (Fig. 2A). Both the proximal and distal cut ends of the peroneal nerve were telescoped into the ends of the PLLA/PGA-c-tube to a depth of 1–2 mm and fixed with epineural 8–0 nylon monofilament suture (Ethicon, Somerville, NJ) with the aid of a surgical microscope. In the PGA-c-tube group, the 40-mm nerve gap was bridged with the 44-mm PGA-c-tube using 8–0 suture to secure each nerve stump inside the nerve guide (n = 15) (Fig. 2B). In the negative control group, after cutting the left peroneal nerve, the cut stump was capped using a silicone cap (As ONE, Osaka, Japan) (n = 15). In dogs of all three groups, neurolysis of the contralateral peroneal nerve was performed as a positive control for comparison (n = 45). Each animal was given a daily intramuscular injection of 100 mg isepamicine sulfate for 1 week. Despite the sensory and related motor deficits, the animals showed normal behavior such as feeding and grooming.

Intraoperative view after implantation of (
Surgical procedures, electrophysiological assessments, animal housing, and enrichment programs were approved by the Kyoto University Animal Care and Use Committee (1989). The veterinary staff monitored the animals on a daily basis, and no animal displayed distress or any secondary complications resulting from peroneal nerve resection, such as self-mutilation, pressure ulcers, sensory neglect, or infection.
Functional analysis
Functional analyses were performed before surgery to obtain baseline control values, and again at 3, 6, and 12 months after the operation. An additional examination was conducted for walking analysis 1 month after surgery. For normalization, values obtained after the operation were expressed as a percentage of positive control values for each dog and were plotted against time.
Walking analysis
To measure articular movement, markers were attached to the sides of the lateral epicondyle of the femur (knee), lateral malleous of the fibula (ankle), and on the basis of the fifth metatarsal bone, and angles made at the ankle were measured with a high-speed camera (160 shots/s, Digital Handy Cam DCR-TRV30; SONY, Tokyo, Japan). Ankle angles were determined from the movement of individual markers. The angles were identified by calculating the mean of three shots taken in the swing phase of walking. As the ankle angle reaches a minimum in the midswing of normal gait, foot-drop deformity such as that caused by peroneal nerve injury is best viewed in the midswing phase. Hence, calculation of this angle allowed us to quantify the degree of improvement in motor function.24,25 When peroneal nerve injury occurs in dogs, the TA muscle is denervated and dorsiflexion of the ankles will not occur. In normal control dogs, the ankle angle of the dogs while walking will be approximately 90°. If the TA muscle is denervated, as in negative control dogs, the angle will be approximately 140° or more. Hence, if the ankle angle becomes smaller with time, functional recovery is evidenced. The final goal of the reconstructed groups was to obtain function that was statistically indistinguishable from the normal control group.
Electrophysiological study
CMAP, SNAP, and motor nerve conduction velocity (MNCV), were measured on both the experimental and positive control sides for each dog in all three groups. All recordings were taken with the use of a Neuropack Σ (Nihon Kohden, Tokyo, Japan). During the electrophysiological studies, the beagle dogs were lightly anesthetized with ketamine hydrochloride (15 mg/kg) and xyladine (7 mg/kg) and placed on a cotton sheet in a warm environment. The temperature of the hind limbs was kept at 37°C by an automatically controlled heating element. To record the CMAP, the peroneal nerve was stimulated with a supramaximal 0.2-mA current pulse through a pair of needle electrodes inserted in an area proximal to the nerve conduit. The evoked muscle response from the TA muscle was recorded through micro needle electrodes placed in the center of the muscle belly from knee and ankle joint. To record the SNAP of the peroneal nerve, a pair of needles was inserted in an area distal to the nerve conduit and microneedle electrodes were placed in an area proximal to the conduit. The latencies of CMAP and SNAP were measured from the point of stimulation to the first deflection of the curve from the baseline to assess the fastest conducting motor or sensory axons. The distance from the site of stimulation to the muscle was kept approximately constant in the repeated studies. The amplitudes of CMAP and SNAP were measured peak to peak. Nerve conduction studies were performed on 45 peroneal nerves on the unlesioned side in the 45 beagle dogs, and the values obtained were used as controls for the amplitude and conduction velocities of CMAP and SNAP. Recovery index was calculated by dividing the peak amplitude on the experimental side by that on the positive control side (Recovery index = Peak amplitude on the experimental side/Peak amplitude on the positive control side) 26 for all three groups.
Evaluation of TA muscle volume using magnetic resonance imaging
Using a magnetic resonance (MR) scanner (40 mT/m maximum amplitude, 200 mT/m/ms slew rate; SIEMENS Sonata 1.5 T; Siemens Medical System, Erlangen, Germany) with a head coil, we measured the volume of the TA muscle, which is innervated by the peroneal nerve. We optimized the sequence for constructive interference in steady-state three dimensions (CISS 3D). The details of the imaging sequences were as follows: repetition time/echo time 7.64/3.82 ms; flip angle 70°; field of view 192 × 256 mm; slice thickness 1 mm; bandwidth 200 Hz/pixel; image matrix 192 × 256; 160 slices. MR images were acquired from the knee to the ankle.
The region of interest (ROI) of the TA muscle for MRI image data was extracted, and the volume of the TA muscle was calculated from three-dimensional reconstructions of the region of interest.
Histological studies
Morphological examination of regenerated nerve tissues
At the end of functional follow-up 3, 6, and 12 months after nerve tube implantation, the dogs were re-anesthetized, and the regenerated nerves were harvested (n = 15 at each follow-up). The nerve tissue was dissected from the surrounding tissues, removed, and fixed in glutaraldehyde (2.5%) in 0.1 M cacodylate buffer (pH 7.4, 4 h, 4°C). The nerve samples were examined at three different positions along the nerve: 5 mm proximal to and 5 mm distal from the stump, and at the midpoint of the nerve conduit. Samples were postfixed for 4 h at 4°C in a solution of 2% osmium tetroxide and 1.5% potassium ferrocyanide in 0.1 M phosphate buffer (pH 7.4, 2 h). The samples were then dehydrated through a graded ethanol series, followed by butyl-2, 3-epoxypropylether, and embedded in Epon 812. For light microscopy, 10-μm-thick longitudinal sections were cut and stained with hematoxylin–eosin. The rest of the nerve samples were sliced transversely to allow us to determine the morphology of the regenerated nerve fibers. After nonspecific immunoreaction was blocked, the samples were incubated in solution of anti-neurofilament (NF) antibody (1:200) (2F11 M0762; Dako Japan, Kyoto, Japan) and anti-S-100 protein antibody (1:100) (Z0311, Dako Japan) overnight at room temperature. They were then washed three times with PBS and incubated in biotinylated anti-mouse rabbit IgG solution (1:200) for 1 h. They were washed three times with PBS, and then horseradish-peroxidase labeled secondary antibody was developed by the diaminobensidine method. Semi-thin sections (0.5–1.0 μm) were cut and stained with toluidine blue for 45 s and examined by light microscopy. Photomicrographs were made at a final magnification of × 400 for measuring the cross-sectional area of the whole nerve, and at × 1000 for morphological analysis. Morphological analysis was conducted on 8–10 randomly selected fields containing at least 500 myelinated fibers. If the whole nerve section had less than 500 fibers, all the regenerated fibers were measured. The total number of myelinated fibers in the reconstructed nerve was estimated from the area occupied by the fibers in the photographs. Morphological evaluation was carried out using image analysis software (Lenaraf 200.xls [version 2.0 for Windows]), and axon, fiber perimeters, and the G ratio (myelinated axon diameter/total myelinated fiber diameter) were obtained. When there was no nerve regeneration at the cross-sectional level, zero values were entered for nerve area and number of myelinated fibers, but the sample was not used for morphometrical measurements. Areas of interest were selected for ultrathin sectioning. The ultrathin sections (50 nm) were stained with uranyl acetate and lead citrate and examined using a Philips EM 201 electron microscope operating at 60 kV. These samples were evaluated to examine the regeneration of the nonmyelinated nerve fibers and the number of Schwann cell nuclei.
Evaluation of muscle atrophy
Atrophy of the TA muscle was evaluated in terms of mean transverse cross-sectional area. The mid portions of the TA muscles from both the treated and positive control sides of each dog in all three experimental groups were cut out and fixed in 10% formaldehyde. Each sample was then dehydrated through a graded ethanol series, followed by dimethylbenzene hyalination, mounted in paraffin, and cut into 4-μm-thick sections. The sections were stained with Masson trichrome, and used to measure the mean cross-sectional areas of the fibers. Two hundred myocytes from 10 fields in muscle transverse sections were measured, and their mean muscle fiber cross-sectional area was calculated.
Statistical analysis
All the data were calculated and expressed as mean values ± SD. The data were analyzed using one-way ANOVA. If there was a significant over all difference between groups, pairwise comparisons were conducted using Scheffe's post hoc test. Differences at p < 0.05 were considered to be statistically significant.
Results
Functional analyses
Walking analysis
All the experiments were completed uneventfully after the surgery. After reconstruction, none of the dogs in any of the experimental groups showed obvious signs of inflammation or infection. The locomotor function of the treated limbs recovered gradually; the dogs in the PLLA/PGA-c-tube and PGA-c-tube groups began to stand with their paws on the experimental side touching the ground at 1 week after the operation and limped while they walked. By 1 month after surgery, they were able to walk without any problem. Six months after surgery, the PLLA/PGA-c-tube and PGA-c-tube groups were able to walk without limping, and coordinated the experimental-side limb well with the other hind limb used as the positive control (Fig. 3A–C). The TA muscle on the experimental side showed no marked atrophy in the PLLA/PGA-c-tube group. In the negative control, however, the animals limped on the treated hind limb, and the limb exhibited TA muscle atrophy, abnormality of metatarsophalangeal joint plantar flexion (known as drop foot), and motion that was uncoordinated with the positive control hind limb even at 12 months after surgery (Fig. 3D–F).

Sequential video frames of walking dogs 12 months after surgery (left to right). Note the dorsiflexion of the left ankle joint. The dorsiflexion appears to be the same as that in the positive control on the opposite side in the PLLA/PGA-c-tube group (
At 1 month, the ankle angle in the negative control group was significantly larger in comparison with the other two experimental groups (p < 0.05) (Fig. 4). From 1 to 3 months, while there were no marked changes in the ankle angle in the negative control, the angle increased in both experimental groups. The ankle angle of the experimental limb in the PLLA/PGA-c-tube and PGA-c-tube groups was significantly smaller than in the negative control group at 3 months (p < 0.05). From 3 to 6 months, while the experimental groups showed recovery indicated by a decrease in the ankle angle, the negative control group did not show any marked change in this respect. At 6 months, the ankle angle of the experimental limb in the PLLA/PGA-c-tube group was significantly smaller (107 ± 3) than that in the PGA-c-tube group (117 ± 4) (p < 0.05). There was no statistically significant difference in the ankle angles in two reconstructed groups at 0, 1, 3, and 12 months after surgery except for at 6 months. The ankle angle on the positive control side and on the experimental side in the negative control group was 92 ± 2 and 143 ± 5, respectively.

Changes in the angle of the ankle joint during the swing phase over 12 months. Locomotor function recovered almost fully in the PLLA/PGA-c-tube group 6 months after surgery.
Electrophysiological evaluation
At 6 months after surgery, CMAP were detected in all dogs in both the PLLA/PGA-c-tube and PGA-c-tube groups. At 12 months, CMAP and SNAP were detected in all dogs in both groups (Fig. 5 and Table 1). Pinching the distal nerves evoked a reflex response (plantar flexion) in anesthetized animals, whereas pinching and cutting the fibrous cover of the tube did not. The average recovery index of CMAP amplitude on the experimental side in the PLLA/PGA-c-tube and PGA-c-tube groups was 0.87 and 0.62, respectively. The differences in CMAP and SNAP amplitude on the experimental side between the PLLA/PGA-c-tube and PGA-c-tube groups were statistically significant (p < 0.05 and p < 0.01). The MNCV on the experimental side in the PLLA/PGA-c-tube and PGA-c-tube groups was 30.7 ± 1.3 and 24.2 ± 2.7 m/s (mean ± SD), respectively. While there was a statistically significant difference between the two groups (p < 0.05), the average MNCV on the experimental side in the PLLA/PGA-c-tube group was 51% of the positive control side value (60.2 ± 3.8 m/s). On the experimental side in the negative control group, no CMAP or SNAP was recorded throughout the experimental period.
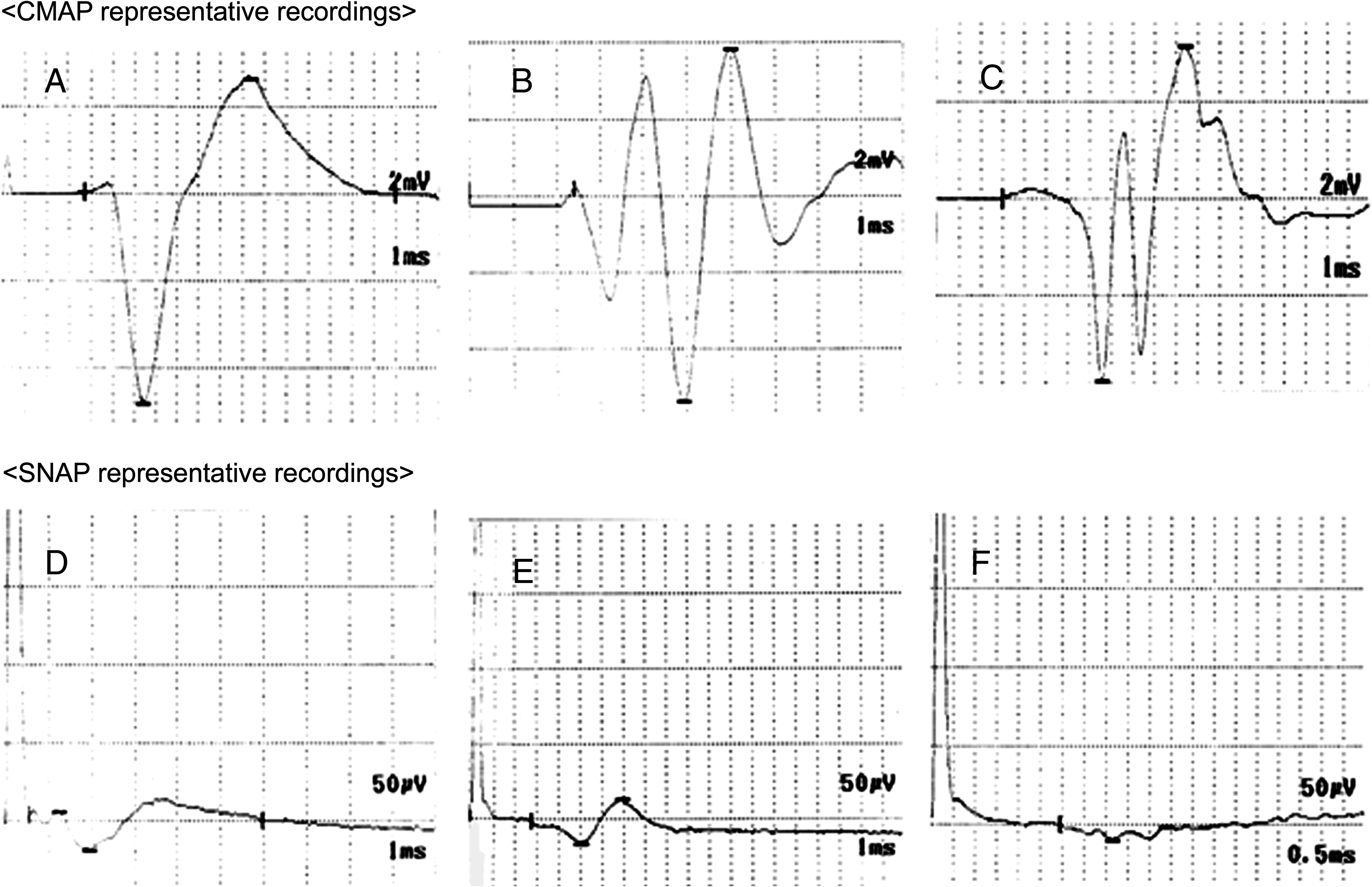
Representative CMAP recordings in the PLLA/PGA-c-tube (
p < 0.01.
p < 0.05.
ns, not significant.
Evaluation of TA muscle volume using MR imaging
At 3 months, all three groups showed a decline in the volume of the TA muscle, although the volume in the negative control group was significantly smaller than in the two experimental groups (p < 0.05) (Fig. 6). While the negative control group continued to show a decline until 12 months, the two experimental groups began to show an increase after 6 months. At 12 months after surgery, the ratio of TA muscle volume (= the muscle volume on the treated side/the muscle volume on the positive control side) differed significantly between the PLLA/PGA-c-tube group (0.80) and the PGA-c-tube group (0.63) (p < 0.05), and the ratio for the negative control group was also only 0.13. This was significantly lower than the average volume ratio of 0.80 for the PLLA/PGA-c-tube group (p < 0.05) (Fig. 7).

Changes in the volume of the tibialis anterior (TA) muscle over 12 months. At 3 and 6 months after surgery, the volumes of the TA muscle in the PLLA/PGA-c-tube and PGA-c-tube groups were significantly better maintained than the negative control group (p < 0.05). At 12 months after surgery, recovery of TA muscle volume in the PLLA/PGA-c-tube group was significantly superior to that in the PGA-c-tube group (p < 0.05).
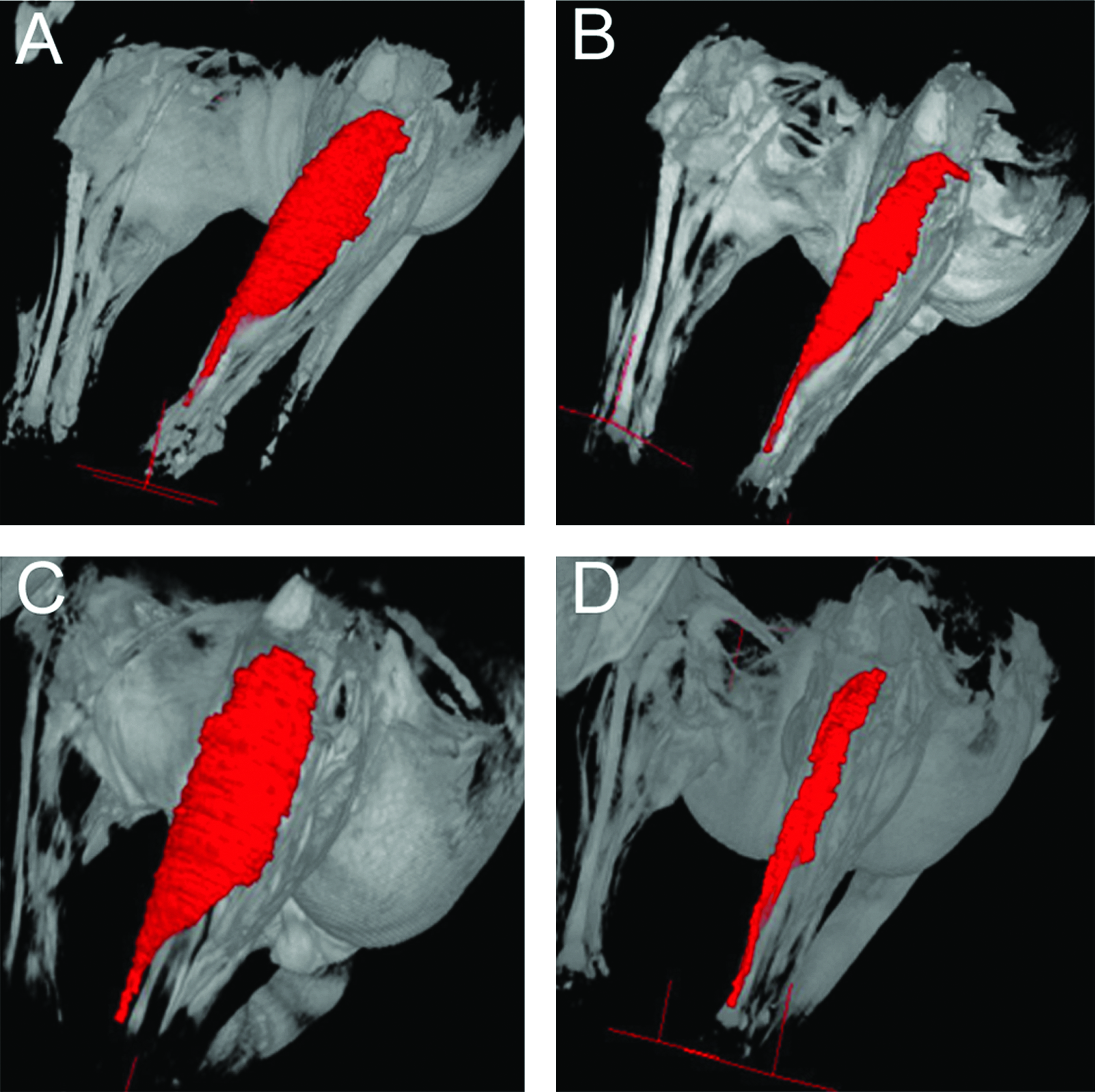
Three-dimensional reconstruction of magnetic resonance imaging volume of the TA muscle 12 months after surgery. (
Histological studies
Morphological examination of the regenerated nerve by light microscopy
At 3 months after surgery, no residual nerve tube was found and only thin fibrous cords remained on the experimental side in the PGA-c-tube group. At 6 months after surgery, the nerve guide in the PLLA/PGA-c-tube group still maintaining its luminal area and a small amount of fibrous tissue was formed around the nerve tube. In the fibrous tissue, newly formed small longitudinally oriented blood vessels were observed in a fibrous capsule. In the PGA-c-tube group, the proximal and distal portions of the regenerated segments were moderately enlarged. The mid portion of the regenerated segments appeared as a thin fibrous cord. At 12 months after surgery, the nerve tube in the PLLA/PGA-c-tube group still retained its lumen, and both the fibrous capsule and the network of blood vessels surrounding the tube had become more extensive (Figs. 2C and 8A). Residual small fragments of PLLA were observed, but in the mid portion of the tube, no infiltration of inflammatory cells was evident. In the mid portion of the tube, typical regenerated nerve cables were surrounded by loose connective tissue, in which a central core of myelinated and unmyelinated axons was arranged in small fascicles. Fibroblasts and blood vessels were also surrounded by loose connective tissues (Fig. 8B). In the PGA-c-tube group, slight to moderate adhesion to the surrounding muscles was observed, and the reconstructed nerves appeared as thin fibrous cords that were well vascularized throughout the full length of the nerve (Fig. 2D). However, neither residual fragments of PGA nor infiltration of inflammatory cells were found in the mid portion of the tube (Fig. 8C). The number of myelinated nerve fibers at the distal end of the PLLA/PGA-c-tube conduit was significantly higher than that at the distal end of the PGA-c-tube (p < 0.05) (Fig. 9(a), (b) and Table 2). The mean diameter of myelinated fibers at the distal nerve on the experimental side in the PLLA/PGA-c-tube and PGA-c-tube groups was 9.0 ± 1.0 μm and 7.4 ± 0.9 μm, respectively, and the difference was statistically significant (p < 0.01). The mean diameter in the PLLA/PGA-c-tube group was the same as that of the normal nerve (9.0 ± 1.5 μm).
(

(
Upper row, PLLA/PGA-c-tube group; lower row, PGA-c-tube group.
G ratio = Myelinated axon diameter/Total myelinated fiber diameter.
p < 0.01.
p < 0.05.
Immunohistochemistry
After immunohistochemical staining with anti-NF and anti-S-100 antibodies, the percentages of NF-immunopositive (neural tissue) areas were measured to compare the PLLA/PGA-c-tube and PGA-c-tube groups. Immunoreactivity for NF was widely observed at the distal end of both conduits 12 months after surgery (Fig. 10A, B). S-100 protein-like immunoreactivity was also observed at the distal end of the conduit in both the PLLA/PGA-c-tube and PGA-c-tube groups (Fig. 10C, D). At the distal nerve in the PLLA/PGA-c-tube group, the percentage of NF-immunopositive (neural tissue) areas was 18.6%, which was significantly higher than that in the PGA-c-tube group (p < 0.01) (Table 2).

Immunohistochemical findings: the nerve in the distal part of the conduit at 12 months after surgery. NF staining (
Morphological examination by electron microscopy
At 12 months after surgery, transmission electron microscopy of the distal stump of the nerve conduit in the PLLA/PGA-c-tube group showed regenerated myelinated fibers dispersed densely in clusters, and also numerous unmyelinated fibers (Fig. 11A). Despite their thinner appearance compared with the normal nerve (Fig. 11C), the myelinated fibers had a compact and uniform structure, including a clear, electron-dense myelin sheath and perfect basal membrane of Schwann cells. In the PGA-c-tube group, the regenerated nerve fibers with thin myelin sheaths were dispersed in clusters in their conduits (Fig. 11B). In addition, the G ratio in the PLLA/PGA-c-tube group was 0.80 ± 0.09. This value was significantly smaller than in the PGA-c-tube group (0.85 ± 0.06) (p < 0.05), but significantly larger than that of the normal nerve (0.70 ±0.02) (p < 0.01).

Transmission electron micrographs of the regenerated nerve in the distal part of the conduit at 12 months after surgery. (
Morphological examination of the TA muscle
At 12 months after surgery, in all three groups, the TA muscle on the experimental side was smaller than on the positive control side to varying degrees, being especially small in the negative control group. Morphological analyses were performed on specimens of TA muscle stained with Masson trichrome. In the PLLA/PGA-c-tube group, TA muscle cells on the experimental side were polygonal and uniform in size with an average cross-sectional area of 2250 ± 227 μm2 (Fig. 12A). Their nuclei were located below the sarcolemma, with no remarkable increase in number. There was a small amount of collagen fibers between the TA muscle cells with no obvious hyperplasia; the average percentage area occupied by collagen fibers was 5.2%. In the PGA-c-tube group, TA muscle cells on the experimental side had various patterns in size and had a rounded appearance when compared with the positive control side, with an average cross-sectional area of 1980 ±230 μm2 (Fig. 12B, C). In the negative control group, however, the overall size of muscle cells was considerably reduced, and the cells were surrounded by extensive collagenous connective tissues. Many large blood vessels were observed (Fig. 12D). However, the average percentage area occupied by collagen fibers was much higher (>20%).

Light microscopy of a transverse section of the TA muscle stained with Masson trichrome at 12 months after surgery. Representative micrographs for the (
Discussion
The development of an artificial nerve tube as an alternative to an autologous nerve graft is a current research focus for peripheral nerve repair. Ideally, nerve guides should be composed of a biocompatible and bioresorbable material that degrades concurrently with new axonal growth 27 and yet maintains some mechanical strength and lumen stability until the regenerated axons reach the distal stump. Tube permeability, durability, and composition strongly influence the capacity for regeneration across long gaps.22,28 With regard to the ideal rate of degradation of a nerve conduit, there is still a degree of controversy among researchers. In general, it has long been believed that a peripheral nerve extends 1 mm/day from the proximal stump, and such descriptions have appeared in many textbooks. However, based on our observations of clinical recovery after surgical reconstructions, the rate seems much slower. We believe that the rate is strongly dependent on the site of the nerve defect, patient age, the blood supply of the surrounding environment, and the time delay between initial injury and reconstruction.
In a previous study, we demonstrated the utility of PGA-c-tubes constructed from PGA and collagen. 29 Both materials are currently in clinical use, 30 but degrade too rapidly in vivo to permit successful repair of longer nerve gaps. In the present study, a bioresorbable nerve guide was created by adding a slowly decomposing fiber, PLLA, to the PGA-c-tube. The PLLA/PGA-c-tube was compared with the PGA-c-tube by implanting both into 40-mm peroneal nerve gaps in a canine model and assessing functional, electrophysiological, histological, and TA muscle recovery at 3, 6, and 12 months after implantation.
Macroscopic and histologic analyses revealed that while the PGA-c-tube rapidly degraded and disappeared 3 months after surgery, PLLA/PGA-c-tube maintained its shape for up to 12 months after surgery. PLLA/PGA-c-tube was gradually reabsorbed over the 12-month experimental period, which was long enough for effective nerve regeneration across the long gap. At 12 months, there were nerve fibers passing though the entire length of the conduit and reaching the distal nerve stump.
Functional ankle angle and MRI analyses showed that implantation of both tubes resulted in TA muscle reinnervation and increased muscle volume, respectively. At 1 month, the ankle angle was much larger in the negative control group than in the two experimental groups. At 3 months, the ankle angle had further decreased in both experimental groups but not the negative control. However, by 6 months, the ankle angle in the PLLA/PGA-c-tube group showed an additional decrease over the PGA-c tube group, although this decrease disappeared by the 12-month study end point. At 3 months, TA muscle volume was reduced in all three groups but was significantly smaller in the negative control group than in the two experimental groups. Additionally, the muscle volume of the negative control group continued to decline up to the 12-month end point, whereas the experimental groups showed increases by 6 months.
The time course of these data suggests a functional response to nerve transection and scaffold-mediated repair that occurs in two phases: degeneration and regeneration. Implantation of the scaffolds appears to provide a comprehensive benefit by both decreasing the amount of early degeneration and subsequently increasing the amount of regeneration. Decreased degeneration of muscle and nerve is probably mediated by reduced atrophy of the TA muscle considering the lengthy degeneration of both walking function and TA muscle volume after injury. We speculate that the reduction of muscle atrophy by scaffold implantation may occur via two mechanisms.
The first is the accelerated extension of a limited subset of axons. Brushart et al. (2002) described that while some regenerated motor axons enter the distal stumps in 4 days, after nerve transaction and direct suturing, others require up to 4 weeks, suggesting the existence of motor neuron populations with different regeneration speeds. 31 It was previously shown that placement of a collagen scaffold with a piled thin film structure inside a nerve conduit enabled fast regenerating axons to reach the distal stump. 21 Trophic factors released by denervated muscles may provide target-derived signals to stimulate neuron survival, axon outgrowth, and maintenance of muscle fiber size, thereby additionally promoting muscle reinnervation.32,33 In the present study, although not enough axons were regenerated to allow detectable electrophysiological activity at early time points, it remains possible that sufficient regeneration occurred to inhibit TA muscle degeneration.
The second proposed antidegenerative mechanism is the enhanced activation of Schwann cells that secrete neurotrophic factors, which in turn could preserve muscle degeneration after nerve transection. After injury and during regeneration, Schwann cells upregulate their synthesis and secretion of several neurotrophic factors, including NGF, brain-derived neurotrophic factor (BDNF), and neurotrophin-3 (NT-3).34–36 NGF stimulates the production and presentation of the neurotrophins BDNF and NT-3 to the outgrowing axons. 37 BDNF prevents the death of axotomized neonatal motor neurons, possibly through promotion of the expression of p75NGFR (low-affinity nerve growth factor receptor). 38 NT-3 enhanced nerve regeneration via increased reinnervation of target muscles and prevention of muscle atrophy after nerve injury. 39 Although the individual roles and effects of these factors vary, they appear to complement each other to reduce muscle atrophy during peripheral nerve regeneration.
In the reconstructed groups, the initial regeneration was followed by functional recovery as demonstrated by a reduction in the stepping angle. This recovery was probably mediated by regeneration of the motor neurons innervating the TA muscle. It is well known that if the gaps are long (>3 cm in human or 1–2 cm in rats), nerve regeneration is impossible without a guide that can bridge the distal and proximal ends. These guides direct the nerve regeneration and concentrate regenerative Schwann cells and growth factors, 40 as well as preventing the formation of fibrous tissue between the distal and proximal ends. ECM molecules such as collagen have been shown to enhance Schwann cell proliferation and induce them to release neurite extension-promoting factors. 41 It is therefore possible that the use of collagen in our nerve guide tubes may have played a role in the observed nerve regeneration.
The number of myelinated fibers, the nerve fiber diameter, and the thickness of myelin sheath are the most reliable parameters to define nerve regeneration. At 12 months after surgery, the MNCV, CMAP, and SNAP amplitudes in the PLLA/PGA-c-tube group were significantly higher than in the PGA-c-tube group, indicating that more myelinated axons had regenerated. 42 Histology showed that the mean diameter of distal nerve myelinated fibers in the PLLA/PGA-c-tube was significantly greater than in the PGA-c-tube that the cross-sectional area occupied by myelinated axons was larger in the PLLA/PGA-c-tube. More myelinated axons were found in the PLLA/PGA-c-tube. These findings suggest that regeneration began earlier in the PLLA/PGA-c-tube than in the PGA-c-tube and are consistent with the electrophysiological and functional analyses.
Full recovery of motor function is the ultimate goal of peripheral nerve regeneration. In the present study, the significant enhancement of motor function at 6 months suggests that regeneration was accelerated in PLLA/PGA-c-tube when compared to PGA-c-tube. We speculate that the rigid mechanical characteristics of PLLA may be responsible for this effect; enhanced rigidity over a longer period of time may provide a wider pathway for Schwann cell and neurotrophic factor migration, in addition to further preventing fibrous tissue formation. Longer maintenance of luminal stability could also provide a more defined guide that could permit extended neuronal regeneration; it has been shown that the time available for proper reinnervation is the main factor determining the final motor and sensory outcome. 3 The number of motor axons that reinnervate muscle fibers is profoundly reduced with prolonged denervation. In addition, long-term denervated muscle fibers fail to fully recover from denervation atrophy, possibly due to exhaustion of the satellite cell population. 40
At 12 months after surgery, transmission electron microscopy of the distal stump of the nerve conduit in the both reconstructed groups showed many clusters of regenerating axons in different stages of myelination, indicating that at this point, the regenerative process was still incomplete. It is therefore possible that additional functional recovery could be observed if the experimental period were increased. However, it is also possible that functional recovery plateaued by 12 months and was prevented from reaching 100% by the long time required for muscle reinnervation; 3 however, this issue will require further investigation.
One notable feature of the present study was its use of MRI to examine muscle atrophy. The MRI data were consistent with data obtained using more conventional methods, indicating that three-dimensional reconstruction using MRI is a promising method for noninvasive evaluation of muscle volume at different stages of nerve reconstruction. Although a few previous studies have assessed functional recovery after nerve reconstruction, to our knowledge no published report has described the use of MRI to assess the volumes; it could therefore be particularly useful for making objective comparisons in clinical settings.
Future research could focus on optimizing the properties of the tube by adjusting the diameter of the PLLA fibers or the method of weaving so that the tube would biodegrade smoothly at an appropriate speed after sustaining the luminal area for a sufficient period of time until regeneration has been accomplished.
Conclusion
Both the PLLA/PGA-c-tube and the PGA-c-tube permitted reinnervation of a 40-mm nerve gap in a canine model; however, functional recovery was accelerated with the PLLA/PGA-c-tube. The PLLA/PGA-c-tube also allowed the development of normal TA muscle cells with a uniform polygonal shape and size. In contrast, regenerating muscle cells in PGA-c-tube-implanted animals exhibited abnormal morphology with different sizes and a rounder form. Both histological and electrophysiological studies indicated that the degree of reinnervation in the PLLA/PGA-c-tube was superior to that in the PGA-c-tube. The PLLA/PGA-c-tube was able to maintain its lumen structure for over 12 months, introducing the possibility of repair of even wider nerve gaps.
Footnotes
Acknowledgments
This work was supported by a Naito Foundation grant, and a grant for young scientists. We would like to thank Satoshi Urushiyama for assistance with preparation of the nerve tubes and Mayuko Fukui for constant support during the research.
Disclosure Statement
No competing financial interests exist.