
Abstract
The microenvironment of bone marrow-derived mesenchymal stem cells (MSCs) strictly regulates their differentiation. In this study, we have developed a new suspension induction method for myocardial differentiation of bone marrow-derived rat MSCs (rMSCs) in vitro on various extracellular matrix (ECM) proteins. Myocardial differentiation of rMSCs was induced with a conventional monolayer method and our suspension method. In our suspension induction, a cell suspension was treated with the medium in the presence of an inducer, incubated for 2 h under a suspension conditions, and moved to a monolayer culture on gelatin-coated, collagen type I-coated, fibronectin-coated, or polystyrene dishes until the total induction time was 24 h. We evaluated the myocardial differentiation by counting the number of colonies of beating cells, performing immunohistochemical staining, and measuring the expression of cardiac-specific gene mRNA using real-time quantitative polymerase chain reaction. We found that rMSCs induced with the conventional monolayer method did not differentiate efficiently, whereas beating cell colonies were found on ECM-coated dishes of suspension-induced cells, after 3 weeks of culture, especially on gelatin-coated dishes. The beating cells were positively stained with anti-troponin T-C antibody and expressed specific cardiac markers. In conclusion, these results demonstrated that the suspension induction followed by subsequent culture on gelatin ECM substrates is a promising method for differentiating rMSCs into cardiomyocytes in vitro.
Introduction
An implantable left ventricular assist device has been proposed as a bridge to transplant for many patients who are on a waiting list for donor organs. 2 Left ventricular assist device can improve organ perfusion, reduce wall stress, and improve functional capacity and quality of life, but it is not an option for the majority of people with heart failure.3,4 Thus, the ultimate goal is to repair the injured myocardium by cell transplantation.
Some fundamental studies and clinical trials suggest that cell-based therapies can improve cardiac function.5–8 The isolation of cardiomyocytes from a patient's heart is unrealistic at present. In general, three types of potential cell sources have been proposed, but the search for these sources and types of cells are still under investigation. 9 One potential source is allogeneic cells, including human embryonic stem cells or fetal allogeneic cardiomyocytes, but there remain ethical issues in their use. Another option is transgenic sources. Genetically engineered animal cardiomyocytes have been studied in an attempt to reduce the rejection reaction in vivo, which is still a long-term problem in recipients. 9
To deal with this problem, autograft bone marrow-derived mesenchymal stem cells (MSCs) are foreseen to be the most promising candidate for transplantation, because they are easy to obtain and less immunogenic than other stem cells. The differentiation of MSCs into cardiomyocytes in vivo has been observed, but it occurs at an extremely low rate and its efficiency is under debate.7,10
The production of autologous beating cardiomyocytes is thus an attractive goal for cell-based therapy. For this purpose, it is preferable to differentiate MSCs into cardiomyocytes in vitro before transplantation, and it is crucial to understand how best to achieve this.
Based on traditional isolation of MSCs and monolayer culture, Wakitani et al. reported that rat MSCs (rMSCs) were differentiated to myogenic cells after 24 h of exposure to DNA-demethylating agent 5-azacytidine, 10 and Makino et al. reported that the repeated treatment of murine MSCs with 5-azacytidine differentiated the cells into cardiomyocytes with high cardiac marker expression in vitro. 11 These findings are in contrast with a report that functional cardiac cells and gene expression were not obtained after treatment with 5-azacytidine. 12 Xu Wang et al. also reported cardiac marker expression in 5-azacytidine-treated MSCs, but they did not observe any beating cells. 13 The differences in these observations might be related to the efficiency of the inducer and the timing of induction.
Clemmons et al. reported that fibroblasts in the suspension did not undergo DNA synthesis and division. 14 Griffin and Houstan reported that cells in monolayer cultures are in a static environment and have a relatively small surface area for diffusion, in contrast to suspension cultures in which the entire surface area is exposed to the drug. 15 In addition, in suspensions, efflux transporters are not retained because of the loss of cell polarity and redistribution of canalicular membranes 16 ; therefore, the compound remains in the cell. Hence, we assumed that by treating the cells with the inducer in suspension culture, the treated cells were more likely to proceed toward the differentiation phase instead of the division phase.
Langer and Vacanti reported that three important components of tissue-engineered constructs were the cell source, soluble chemical factor, and extracellular matrix (ECM). 17 ECM proteins and the cooperation between signaling pathways triggered by soluble factors such as growth and differentiation factors were found to determine cell proliferation and cell differentiation. 18 In our previous study, ECM components were seen to affect the beating behavior of primary neonatal cardiomyocytes and cardiac differentiated P19.CL6 cells in which enhanced beating behavior and cardiac differentiation on gelatin-coated dishes were observed. 19
The aim of our studies was to produce spontaneously beating cardiac cells from rMSCs by our new induction method on different substrates. Optimal substrates for stem cell attachment, proliferation, and differentiation have been reported for various types of stem cells. 20 In this study, treated rMSCs were cultured on gelatin-coated, fibronectin-coated, collagen type I-coated, and polystyrene dishes. We treated rMSCs using a newly established suspension method, and the differentiation tendency was compared with those treated by the conventional monolayer method.
Materials and Methods
Bone marrow cell preparation
Femora and tibiae of 4-week-old, male Sprague Dawley rats with average body weight of 80 g were collected and adherent soft tissues were removed. Institutional guidelines for the care and use of laboratory animals were observed. The rMSCs were obtained from collected femora and tibiae by flushing the marrow cavities. Isolated cells were cultured in high-glucose Dulbecco's modified Eagle's medium (DMEM-HG; Gibco, Grand Island, NY) supplemented with 10% fetal bovine serum (lot no. 7297H; MP Biomedicals, Eschwege, Germany), 5% heat-inactivated horse serum (lot no. 076K8430; Sigma-Aldrich, St. Louis, MO), and penicillin (100 U/mL)/streptomycin (100 μg/mL) (Wako, Osaka, Japan).
The cells were seeded on 10 mm fibronectin-coated dishes (BD Falcon, BD BioCoat, BD Biosciences, Bedford, MA) and incubated in a 5% carbon dioxide (CO2)/air atmosphere at 37°C. At 24 h after plating, nonadherent cells were removed, and the medium was changed every 3 days until the adherent cells reached 80% confluence. The cells in one dish were harvested with 0.25 mg/mL trypsin (Lonza, Walkersville, MD), washed with phosphate-buffered saline (PBS), and seeded onto three new dishes.
Isolation of neonatal heart
Cardiomyocytes were isolated from neonatal (2-day-old) Sprague Dawley rat hearts by the collagenase digestion method with modifications.21,22 Institutional guidelines for the care and use of laboratory animals were followed. The hearts were removed and carefully minced with a scalpel blade into fragments and rinsed several times with Hanks' balanced salt solution (Sigma-Aldrich) to remove blood and cellular debris. The minced hearts were gently stirred in 50 mL collagenase solution (0.15 M NaCl, 5.63 mM KCl, 0.02 M HEPES, 0.02 M NaHCO3, 3.74 mM CaCl2 · 2H2O, and 6.5 × 104 U collagenase [lot no. 06032W; Wako]) at 37°C for 30 min. The resulting cell suspension was filtered through a 40 μm pore-sized nylon cell strainer (BD Falcon, BD BioCoat, BD Bioscience) and centrifuged at 78 g for 3 min.
Isolated cardiomyocytes were cultured in minimum essential medium alpha (Gibco) supplemented with 10% (v/v) fetal bovine serum (lot no. 7297H; MP Biomedicals) and 100 IU/L penicillin–streptomycin (Wako) on 60 mm gelatin-coated dishes (Iwaki; Asahi Glass, Tokyo, Japan). Three days after isolation the mRNA levels of the cardiac marker genes were evaluated.
Cardiomyocyte differentiation
Monolayer induction
The rMSCs at fourth passage were seeded on 60 mm gelatin-coated dishes (Iwaki; Asahi Glass), fibronectin-coated dishes (BD Falcon, BD BioCoat, BD Biosciences), collagen type I-coated dishes, and noncoated polystyrene dishes (Iwaki; Asahi Glass) at a density of 1.0 × 105 cells/dish. The cells were cultured at 37°C in humidified air with 5% CO2, reaching 80% confluence within 3 days. Afterward the cells were exposed to the inducers, 10 μM 5-azacytidine (Nacalai Tesque, Kyoto, Japan), 300 μM
Suspension induction
The suspension of 1.0 × 105 rMSCs was treated with and without 10 μM 5-azacytidine (Nacalai Tesque), 300 μM
The experiments were repeated to determine the expression of troponin C type-2 after suspension induction.
Total RNA isolation and reverse transcription
Total cellular RNAs from both noninduced and induced rMSCs with monolayer induction and suspension induction were extracted by QuickGene RNA cultured cell kit S (Fujifilm Life Science, Tokyo, Japan) after 1, 2, and 3 weeks of culture. In another experiment, total cellular RNAs from induced rMSCs with suspension induction were extracted after 1, 2, 3, 4, and 5 weeks. The cellular RNAs from neonatal cardiomyocytes were also extracted with the same protocol after 3 days of culture as a positive control for real-time quantitative polymerase chain reaction (PCR). Total cellular RNAs were calculated as follows: [RNA] = A260 (nm) × Dilution × 40 μg/mL. The RNAs from beating and nonbeating colonies were extracted separately.
First-strand cDNAs were synthesized using a mixture of oligo(dT)18 primer. Total cellular RNAs (200 ng) were incubated with 2.5 μM oligo(dT)18 primer at 70°C for 10 min to denature RNA secondary structure and then incubated at 4°C to let the primer anneal to the RNA. A given amount of 5 × reverse transcriptase (RT) buffer (Toyobo, Osaka, Japan) and 2.5 mM dNTP mixture (Takara Bio, Shiga, Japan) (4 μL) were added and incubated at 37°C for 5 min. Reverse transcriptase (100 units; Toyobo) was added into the mixture and the RT reaction was extended at 37°C for 1 h. Then the reaction was heated at 94°C for 5 min to inactivate the enzyme and cooled at 4°C for 15 min. RNase (DNase-free, 0.5 μg; Roche Diagnostics GmbH, Mannheim, Germany) was added into the mixture and incubated at 37°C to remove the template RNA. To confirm that the beating cells were cardiomyocytes, an immunochemical study was conducted, in which the expressions of cardiac-specific markers troponin C type 1 (slow, TNNC 1), troponin T type 2 (cardiac, TNNT 2), troponin I type 3 (cardiac, TNNI 3), GATA binding protein 4 (GATA 4), and myocyte enhancer factor 2D (MEF2D).
Real-time quantitative PCR
Real-time quantitative PCR was conducted with SYBR Green. Primers for PCR analysis of troponin T type-2 (cardiac, TNNT 2), troponin C type-1 (slow, TNNC 1), troponin I type-3 (cardiac, TNNI 3), GATA4, MEF2D, and troponin C type-2 (fast, TNNC 2) were designed using Primer Express software (Perkin-Elmer Applied Biosystems, Warrington, UK). Primer sequences are shown in Table 1. The reaction mixtures contained 23.74 μL distilled water, 25 μL SYBR Green Real-Time PCR master mix (Toyobo), 100 nM of each primer, and 0.26 μL cDNA. The thermal profile for PCR was 50°C for 2 min, followed by 95°C for 10 min, and then 40 cycles of 15 s at 95°C and 1 min at 60°C. We also performed a negative control PCR reaction using 0.26 μL distilled water to ensure the absence of template contamination in PCR reagents. The cycle number at which the reaction crossed an arbitrarily placed threshold (Cτ) was determined for each gene. The average Cτ values of triplicate measurements were used for all subsequent calculations on the basis of the delta Ct method (ΔCτ). The amount of mRNA levels was determined by 2ΔCτ. To correct any variation in mRNA content, the quantities of the genes of interest were normalized by the quantity of glyceraldehyde-3-phosphate dehydrogenase and expressed as relative values of mRNA.
nt, nucleotide.
Immunostaining analysis
To confirm the protein expression in addition to the mRNA expression, cells generated by the monolayer method and beating cells generated by suspension induction were stained with anti-troponin T-C antibody. After 4 weeks of culture, the cells were fixed with 10% formalin in PBS and washed with PBS three times. Next, the cells were incubated for 5 min in 0.1% hydrogen peroxide in PBS to quench endogenous peroxide activity and washed in PBS twice for 5 min each. Then the cells were incubated with 10% Block-Ace™ (Dainippon Sumitomo Pharma, Osaka, Japan) in PBS for 20 min to suppress nonspecific binding of IgG. After three cycles of washing with PBS for 5 min each, the cells were incubated with 2.5 μL/mL primary antibody (troponin T-C(C-19), sc-8121; Santa Cruz Biotechnology, Santa Cruz, CA) for 60 min in PBS with 1.5% Block-Ace™, washed three times in PBS for 5 min, and incubated with 2 μL/mL secondary antibody (donkey anti-goat IgG-FITC, sc-8121; Cosmo Bio, Tokyo, Japan) for 45 min in PBS with 1.5% Block-Ace™. The cells were washed with PBS four times and mounted with aqueous mounting medium.
Stained cells were observed using Nikon Eclipse TE 300 (Nikon) fluorescence microscope. An image was taken using Image Pro 4.5 software (Media Cybernetics) with the following parameters: for bright file, an exposure time of 20 ms and gain of 7; for fluorescence, an exposure time of 2 s and gain of 7.
Statistical analysis
All data are presented as means ± standard deviations. Statistical analysis was performed using Student's t-test. A p-value of less than 0.05 was considered significant.
Results
rMSCs form myotubes
After 3 weeks of cultivation, the shape of the cells induced with suspension induction was very different from that with monolayer induction, as shown in Figure 1. The shape of the suspension-induced cells appeared to be myotubular and seemed to correlate closely to beating colony formation. The phenotypic difference in these shapes was confirmed by measuring TNNT 2, TNNC 1, and TNNC 2 expression.
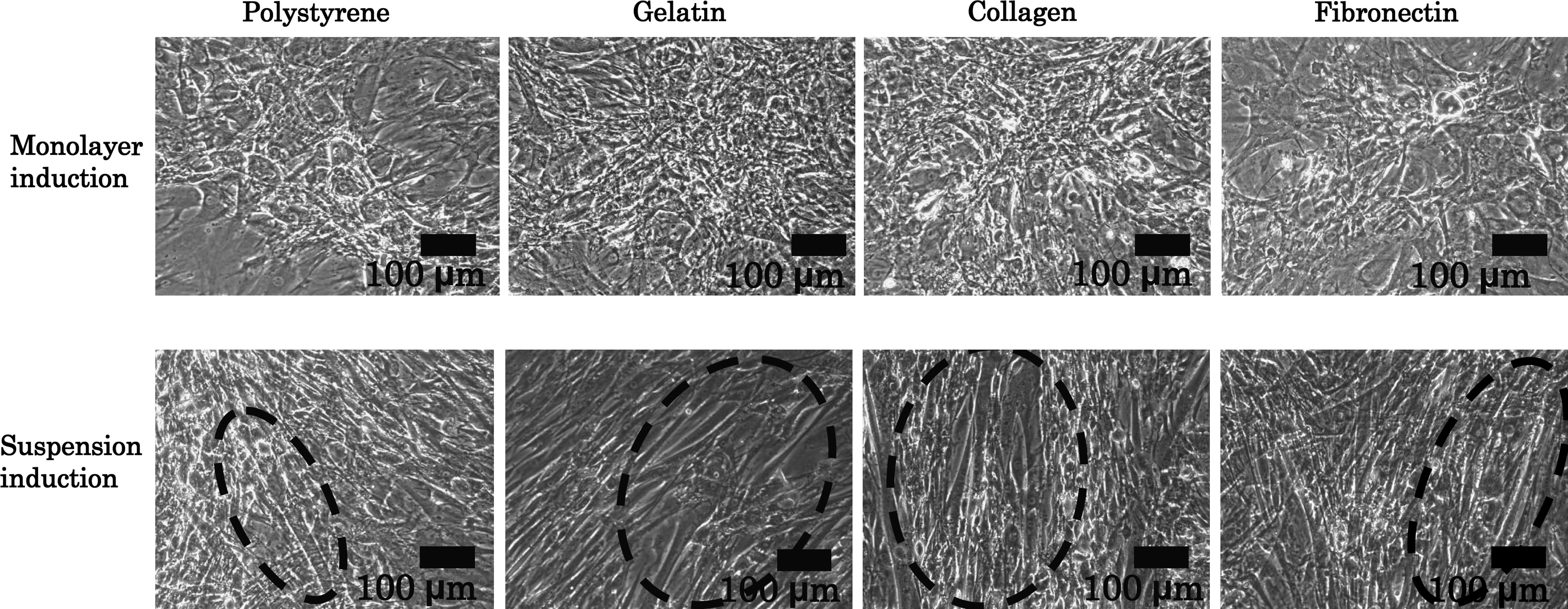
Microscopic image of rMSCs after monolayer or suspension induction and 3 weeks of culture on several types of dishes. Dashed regions represent the regions of cells with myotube-like shape. rMSC, rat mesenchymal stem cell.
Expression of cardiomyocyte-associated genes in monolayer and suspension induction
Troponin T type-2 (cardiac, TNNT 2) and troponin C type-1 (slow, TNNC 1) are known to be markers of cardiomyocytes,23,24 and troponin C type-2 (fast, TNNC 2) is reported to be expressed at the early stage of the cardiac development.
23
In this study, the expression of TNNT 2 was higher in the suspension induction than in the monolayer induction, as shown in Figure 2A. On the other hand, TNNC 1 expression was generally lower in the suspension induction than in the monolayer induction (Figure 2B). However, the gene expression of TNNC 2 was detected only in the suspension induction and not in the monolayer induction (Figure 2C). These results indicate that the suppression of TNNC 1 may have been affected by the expression of TNNC 2 and signify the initial stage of cardiac differentiation, as suggested by Stoutamyer and Dhoot.
23
In addition, the gene expression of TNNT 2 and TNNC 2 were detected only in rMSCs treated with inducers and not in the rMSCs treated without inducers (Supplemental Fig. S1 available online at
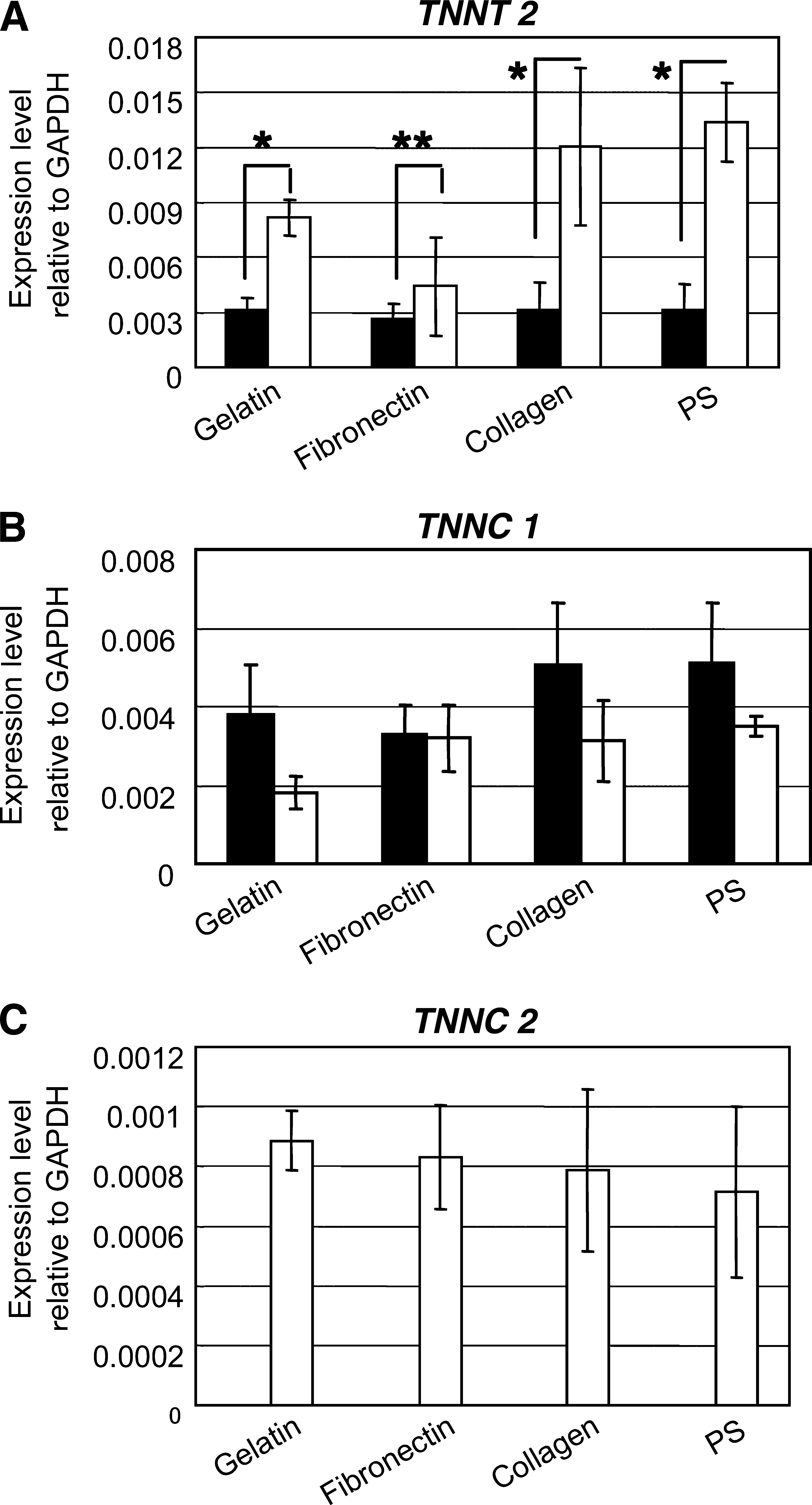
Expression levels of (
In other experiments, the expression of TNN2 was detected after 2 weeks of differentiation and decreased by culture time as shown in Figure 3. This observation is possibly related to the cardiomyocyte differentiation. Besides, this result shows similarity with that during quail heart development in ovo. 23

The expression of TNNC 2 decreased with culture period. Data are means ± standard deviation; n = 3 for each sample.
Myotube-like cells on ECM substrates show spontaneous contraction
In general, about 3 weeks are needed to observe spontaneous beating of the cells without the addition of any chemical reagent, such as acetylcholine.10,25 Once the beatings are detected, it takes about another 1 week to enter the synchronous stage.
Interestingly, the beating cells and colonies were detected only after they were induced with suspension induction on ECM protein-coated dishes, but not in monolayer induction. We carried out these induction experiments 14 times and found a beating colony only once in monolayer induction on gelatin-coated dishes. As five dishes were used for each experiment, the average number of beating colonies in one dish was calculated as 0.75 ± 1.5 (Table 2). However, the real probability of beating colony appearance was much lower than this value. A large number of beating colonies (4.5 ± 0.6) with sizes ranging from 400 to 500 μm were found in the five gelatin-coated dishes, and 1.3 ± 1.5 beating colonies with a similar size were found in the five fibronectin-coated and collagen type-I-coated dishes. No beating cells were detected in noncoated polystyrene dishes in either form of induction. Table 2 summarizes the colonies of beating cells. Supplementary Video S1 (available online at
Five 60 mm culture dishes were used for calculation of average beating colony number.
In some cases, monolayer-treated and suspension-treated rMSCs were detached from the dishes after 3 weeks of culture. The nondetached cells proliferated and only suspension-treated rMSCs became beating cells after 6 weeks of culture (data not shown).
Immunostaining
Immunofluorescence examination clearly showed that when the dishes were stained with anti-troponin T-C antibody, the beating areas were positively stained as shown in Figure 4, but cells treated by monolayer induction remained at the background level of staining.

rMSCs induced by monolayer or suspension method were stained with anti-troponin T-C antibody.
Expression of cardiomyocyte-associated genes in beating and nonbeating cells
To define the phenotype of the beating cells, the expression levels of cardiac-specific genes TNNC 1, TNNT 2, TNNC 2, TNNI 3, GATA 4, and MEF2D were evaluated. Expression levels of given genes were assessed using RNAs from neonatal hearts as positive qualitative controls. The expression levels of TNNC 1, TNNT 2, and MEF2D were higher in beating cells than in nonbeating cells, as shown in Figure 5A and B. The expressions of TNNI 3 and GATA 4 were detected in only one of four isolated colonies of beating cells and were not detected in any nonbeating colonies. The high expression of TNNC2 in Figure 5A is possibly because either skeletal muscle cells or initial cardiomyocytes are also present in the beating colonies.
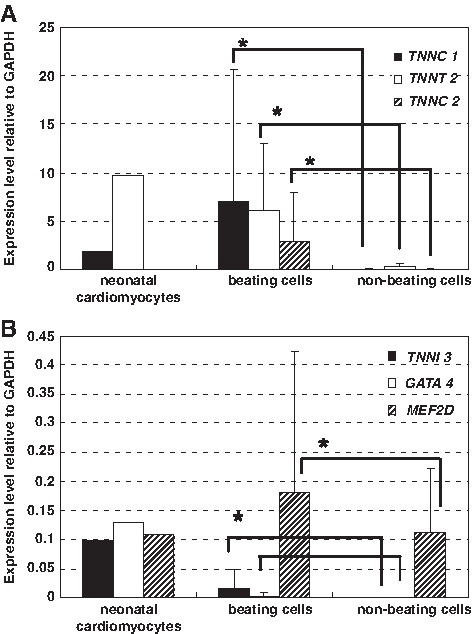
(
In neonatal cardiomyocytes (3 days of cultivation), TNNC 1, TNNT 2, TNNI 3, and GATA 4 were expressed (Fig. 5A, B). However, TNNC 2 was not expressed. These data suggested that 25% of the beating cells were cardiomyocytes and 75% were late-maturing cardiomyocytes.
Discussion
MSCs derived from bone marrow are useful cells because they can be isolated from patients and can differentiate into many types of cells. The production of autologous beating cardiomyocytes is an attractive goal for cell-based therapy. However, in previous studies, differentiation into cardiomyocytes occurred at extremely low rates.7,10 Therefore, it is essential to establish a new, more effective system for differentiating MSCs into beating cardiomyocytes in vitro before being transplanted into patients. Other reports have demonstrated that rat and mouse bone marrow cells can differentiate into cardiomyocytes in vitro.10,11 On the other hand, Liu et al. reported that 5-azacytidine could not expand rMSCs or induce their differentiation into cardiomyocytes. 12 In our experience from 14 experiments in suspension and monolayer induction, beating cells were obtained in only 7 experiments with suspension induction and in 1 with monolayer induction. These results suggested that beating cardiomyocytes were not easily obtained after exposure to 5-azacytidine. In this regard, the induction method and the substrate are important to obtain beating cardiomyocytes.
Some reports have described manipulating microenvironmental factors, such as cell dimensions, controlled delivery of soluble factors, chemical cues, mechanical cues, and culture substrates, for the controlled differentiation of stem cells.26,27 Cells in monolayer culture are in a static environment and have a relatively small surface area for diffusion in contrast to cells in suspension culture.
15
In this study, more than 90% of cells induced with suspension method were adhered onto the dishes at 24 h after the cells were inoculated into monolayer cultures (Supplemental Fig. S2 available online at
In this study, the expression of TNNT 2 was higher in suspension induction than in monolayer induction, as shown in Figure 2A. On the other hand, TNNC 1 expression was generally lower in suspension induction than in monolayer induction (Figure 2B). However, the gene expression of TNNC 2 was detected only with the suspension induction and not with the monolayer induction (Figure 2C). These results indicated that differentiation efficiency was affected by suspension induction. The enhancement mechanism is unclear but may be related to the proliferation activity, which had not yet started in the suspension condition.
The expression levels of cardiac-specific genes TNNT 2, TNNC 1, and MEF2D of the beating cells were higher than those of nonbeating cells and almost the same level as those of neonatal cardiomyocytes. The beating of neonatal cardiomyocytes weakens with culture time. In these experiments, the gene expression of neonatal cardiomyocytes was measured at 3 days after cultivation, to use the well-beating cardiomyocytes as control cells.
Troponin, a calcium-receptive protein complex involved in the contraction of striated muscles, consists of three components: a calcium-binding component (troponin C), a component that inhibits contractile interaction (troponin I), and a tropomyosin-binding component (troponin T). 28 These contractile proteins are expressed in the developing mammalian heart. 24 Therefore, cardiac troponin T (TNNT2), slow troponin C (TNNC1), and cardiac troponin I (TNNI 3) are known to be markers of cardiomyocytes.23,24 Myocyte enhancer factor (MEF2) genes are recognized to be expressed in differentiated skeletal and cardiac muscle. 29 MEF2D and GATA-4 are regulatory factors for cardiac troponin I. 30 Arceci et al. and Edmondson et al. reported that the GATA-4 gene was expressed in the early stage 31 and MEF2D in the middle stage of heart development. 32 Fukuda also reported that the MEF2D gene was expressed in cardiomyogenic cells after differentiation. 33 Consequently, in this study, we measured the expression of TNNT2, TNNC1, TNNC2, TNNI3, GATA-4, and MEF2D to examine the differentiation of rMSCs to cardiomyocytes.
Akavia et al. reported that GATA 4 and TNNI 3 were expressed only in cardiac muscle cells and not skeletal muscle cells. 34 However, the expression levels were decreased by culture time and were almost undetectable after 10 days of culture for TNNI 3. Referring to this report, we successfully differentiated the MSCs into beating cardiomyocytes by suspension induction. We concluded that expression of GATA 4 and TNNI 3 was not detected in the other three beating colonies because of the long cultivation period. TNNC 2 was also expressed in beating cells. The high expression of TNNC2, shown in Figure 5A, is possibly because either skeletal muscle cells or initial cardiomyocytes were also present in the beating colonies. As shown in Figure 3, TNNC 2 expression was decreased by the culture period. This observation was similar to that of Stoutamyer and Dhoot, who detected TNNC 2 on day 3 in quail hearts in ovo but could not detect it by day 17 using RT-PCR. 23
Interestingly, our results indicated that the spontaneously beating cells were detected only on ECM protein-coated dishes and were induced with suspension induction. The beating cells or colonies were not detected in noncoated polystyrene dishes. These results indicated that the ECM substrate strongly affected cellular differentiation. The number of beating cell colonies on gelatin-coated dishes was much higher than on other substrate-coated dishes. This was similar to the result in our previous study, in which the beat and the beating period of isolated neonatal rat cardiomyocytes on gelatin-coated dishes were stronger and longer than on fibronectin- or collagen-coated dishes or on noncoated polystyrene dishes. 19 This phenomenon may be related to the cell biological activity and the physical properties of the substrate. It has been reported that during cell culture, large amounts of fibronectin are produced, which associates with collagen in a way that promotes fibrillogenesis. 35 It was also reported that fibronectin binds to gelatin more strongly than to collagen.35,36 Therefore, fibril might be produced at higher levels on gelatin-coated dishes than on fibronectin- or collagen-coated dishes.
A very large difference was observed in the number of beating colonies on gelatin-coated and collagen type I-coated dishes. The dynamic storage modulus of gelatin is higher than that of collagen type I. 37 The high dynamic storage modulus of gelatin, as well as substantial movement and stretching, may have allowed easier contraction of differentiated rMSCs, which might have contributed to the large difference in the number of beating colonies. It was also reported that the elasticity of the substrate affects the differentiation of naive MSCs and mouse myoblast C2C12 cells into myogenic cells.38,39
In this study, cardiac gene expression analysis and immunostaining were performed to verify whether the beating cells derived from treated MSCs were cardiomyocytes or not. The beating cells and colonies were detected at 3 weeks after treatment ended and became synchronous after 1 week. Therefore, the immunostaining was done at 4 weeks after treatment ended. The results of immunofluorescence and cardiac gene expression demonstrated that the beating cells expressed cardiomyocyte-like phenotypes.
Conclusion
In conclusion, this study demonstrated that suspension induction is a promising method for differentiating rMSCs into cardiomyocytes in vitro. In addition, the ECM protein is seen to affect the differentiation of rMSCs into beating cells. These findings indicate a bright future for the production of autologous beating cardiomyocytes for cell-based therapy and myocardial patch scaffold. In this study, a gelatin-based niche was found to be preferable for cardiac differentiation and beating function. Nevertheless, further study is required to characterize the full mechanism of the suspension method for the differentiation of MSCs into cardiomyocytes as well as the effect of differentiated MSCs after transplantation in vivo.
Footnotes
Acknowledgments
This work was supported by a CREST-JST and partly supported by a Grant-in-Aid for Scientific Research (B) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan. The authors thank Dr. Tomo Ehashi for his outstanding technical assistance. The primary author A. Miskon thanks the Ministry of Higher Education of Malaysia and Tun Hussein Onn University of Malaysia for funding his study under the Academic Training Scheme.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.