
Abstract
Whole organ engineering would benefit from a three-dimensional scaffold produced from intact organ-specific extracellular matrix (ECM). The microenvironment and architecture provided by such a scaffold would likely support site-appropriate cell differentiation and spatial organization. The methods to produce such scaffolds from intact organs require customized decellularization protocols. In the present study, intact adult porcine hearts were successfully decellularized in less than 10 h using pulsatile retrograde aortic perfusion. Serial perfusion of an enzymatic, nonionic detergent, ionic detergent, and acid solution with hypotonic and hypertonic rinses was used to systematically remove cellular content. The resultant cardiac ECM retained collagen, elastin, and glycosaminoglycans, and mechanical integrity. Cardiac ECM supported the formation of organized chicken cardiomyocyte sarcomere structure in vitro. The intact decellularized porcine heart provides a tissue engineering template that may be beneficial for future preclinical studies and eventual clinical applications.
Introduction
Most commercially available biologic scaffold materials are manufactured as thin sheets. The source tissues are typically decellularized by immersion and agitation in a combination of salt solutions, detergents, and enzymatic solutions. Due to the density, mass, and 3D architecture of most whole organs such as the heart, liver, and kidney, these approaches are ineffective at removing cellular material.1,23 Recently, Ott et al. described the decellularization of rat hearts by vascular perfusion. 19 Decellularization of a porcine heart was shown to be possible, but a comprehensive, reproducible, and time-effective decellularization technique was not provided.
The purpose of this article is to describe a decellularization method for a complex organ, specifically the porcine heart, by pulsatile retrograde aortic perfusion to generate cardiac extracellular matrix (C-ECM). Decellularization was confirmed by immunohistochemical (IHC) methods and DNA quantification. The C-ECM was characterized by immunohistochemical (IHC) analysis, scanning electron microscopy (SEM), and mechanical testing.
Materials and Methods
Preparation of C-ECM
Porcine hearts weighing approximately 300 g were obtained immediately after euthanasia of adult pigs. Excess fat and connective tissue were removed, and the ventricles were rinsed with water to remove coagulated blood. Each heart was frozen at −80°C for at least 16 h for storage and to aid in cell lysis. The hearts were then thawed in type 1 reagent–grade water at room temperature. The aorta was cannulated with a ½″ to ¼″ straight barbed reducer and connected to ¼″ internal diameter (ID) silicone tubing. Each heart was placed in a 4 L beaker containing 3 L of hypotonic type 1 water that was recirculated using a peristaltic pump (L/S® Drive EW-07550-30; Cole-Parmer, Vernon Hills, IL) for 15 min at 1 L/min. The type 1 water was replaced with 2 × phosphate-buffered saline (PBS) at 1 L/min each for 15 min. Three liters of 0.02% trypsin/0.05% ethylenediaminetetraacetic acid/0.05% NaN3 solution was warmed to 37°C using a digital hotplate and then perfused through the myocardial vasculature at 1 L/min for 2 h. A 3% Triton X-100/0.05% ethylenediaminetetraacetic acid/0.05% NaN3 was then used for perfusion followed by a 4% deoxycholic acid solution at 1.3 L/min each for 2 h at room temperature. After each chemical solution was used as a perfusate, type 1 reagent–grade water was perfused through the heart for approximately 5 min with no recirculation followed by recirculating 2 × PBS for 15 min to aid in cell lysis and removal of cellular debris and chemical residues. Disinfection was accomplished by perfusion of 0.1% peracetic acid/4% ethanol at 1.7 L/min for 1 h. The acid was neutralized and removed from the ECM by perfusing the intact matrix with PBS (pH 7.4) two times and type 1 water three times for 15 min each at 1.7 L/min. Fluid pressure was measured at the aorta during the entire decellularization process. The free walls of the left ventricle (LV) and right ventricle (RV) were excised, and were either used immediately for mechanical properties testing in the hydrated state or laid flat on nonstick aluminum foil, frozen at −80°C for at least 2 h, and then lyophilized until dry for biochemical analysis.
Immunohistochemistry and immunofluorescence studies
Full-thickness samples of C-ECM and native (nondecellularized) ventricles were fixed in 10% formalin and then paraffin embedded. Eight-micron-thick sections were cut and deparaffinized. Hematoxylin and eosin and 4′,6-diamidino-2-phenylindole were used to evaluate the presence of nuclear material. Movat's Pentachrome stain was used to allow observation of the distribution of nuclei, elastic fibers, collagen, glycosaminoglycans (GAGs), fibrin/fibrous structures, and muscle. Herovici stain was used to discriminate and observe collagen I and III in the ECM scaffolds. In addition, collagen IV, a basement membrane component, was observed using a mouse anti-human collagen IV antibody. Slides were imaged using a Nikon™ E600 microscope with 4 × and 20 × objectives and captured using MetaVue™ Software package (Molecular Devices, Sunnyvale, CA).
Scanning electron microscopy
C-ECM ventricle and native ventricle were fixed with 2.5% glutaraldehyde followed by dehydration by 1% osmium tetroxide. The samples were then dehydrated in a graded series of ethanol concentrations in PBS. The samples were sputter coated with 3.5 μm of gold and observed using a Jeol 9335 field emission gun SEM (Jeol, Tokyo, Japan) to capture standard scanning electron digitized images at 1000 × and 30 ×.
DNA quantification
Approximately 10 mg of native and ventricular C-ECM was digested with 0.1 mg Proteinase K (Sigma-Aldrich, St. Louis, MO) in 1 mL of PBS at 37°C on a rocker overnight. The digest was then purified using phenol/chloroform/isoamyl alcohol. 24 The Quant-iT™ PicoGreen® dsDNA assay (Molecular Probes, Eugene, OR) was used for quantification of the amount of DNA using the manufacturer's instructions. Samples were evaluated in triplicate. Equal volumes of digest were separated by gel electrophoresis in a 1% agarose gel with ethidium bromide at 60 V for approximately 1 h, and the gel was observed under ultraviolet transillumination to determine the fragment size of residual DNA.
GAGs and elastin quantification
Total sulfated GAGs, and cross-linked elastin within the C-ECM were determined using the manufacturer's instructions (Biocolor, Carrickfergus, United Kingdom). One sample of three lots each of lyophilized C-ECM LV and RV as well as one sample of LV and RV from native ventricle were digested with papain for the GAG and elastin assay. Samples were evaluated in duplicate, and all values were normalized to 1 mg dry sample for comparison. A two-tailed Student's t-test was performed to determine whether differences existed between the GAG and elastin contents of native ventricle versus C-ECM with the p-value set at 0.05 (Minitab® version 15.1.1.0; Minitab, State College, PA).
Ball burst mechanical testing
An MTS Insight with a 2000 N MTS load cell model 569327-03 (MTS, Eden Prairie, MN) was used with a ball burst compression cage (Instron, Norwood, MA) to measure the biaxial burst strength of native LV and RV and the C-ECM derived from each ventricle. The test was performed in accordance with ASTM D3787-07 Test Method for Bursting Strength of Textiles-Constant-Rate-of-Traverse Ball Burst with deviations for sample geometry. Similar methodology has been used in multiple published studies to describe the biaxial strength of synthetic and biologic scaffolds.23,25–27 The native ventricles were excised and tested the same day as euthanasia. The C-ECM was tested within 48 h of the completion of decellularization. For all groups, the specimen was clamped in the fixture such that the polished ball contacted the endocardium. A 25.4 mm polished steel ball was advanced at a constant rate (25.4 mm/min) through the test material. Each experiment was conducted three separate times for each ventricle in the native and decellularized form. A two-tailed Student's t-test was performed to determine whether differences existed between the maximum force to failure and extension at maximum force for the LV and RV C-ECM compared with the values for native LV and RV with the p-value set at 0.05 (Minitab version 15.1.1.0; Minitab).
Cell seeding
Lypophilized C-ECM sheets were packaged in sterilization pouches and terminally sterilized using ethylene oxide. As previously described, Hamburger-Hamilton Stage 31 (day 7) White leghorn chicken embryonic cardiomyocytes (CMs) were isolated using collagenase and trypsin digests and preplated for 1 h to selectively isolate CMs.28,29 CMs were seeded on the luminal side of the C-ECM sheet. Approximately 500,000 cells/cm2 were cultured on the ECM scaffolds for 4 days in Dulbecco's modified Eagle's medium with 10% fetal bovine serum, 1% chick embryo extract, and 1% penicillin and streptomycin. The scaffolds were then fixed in 4% paraformaldehyde for 20 min followed by rinsing with PBS. Monoclonal anti-α-actinin antibody (clone EA-53; Sigma-Aldrich) and anti-β-tubulin antibody (TU-06; Abcam, Cambridge, MA) were used to distinguish CMs from other cell types. 28 The scaffolds were placed between two cover slips and imaged using an inverted Olympus Fluoview 1000 confocal microscope (Olympus, Center Valley, PA).
Results
Preparation of C-ECM
At the beginning of the decellularization process, the mean perfusion pressure was recorded as approximately 150 mm Hg. During the trypsin step, the hearts lost some of their red–brown coloration and became more flaccid, and the reagent became pinkish in appearance. With each subsequent reagent change, the hearts became whitish in appearance and expanded to approximately twice the original volume (Fig. 1). Although the flow rate was increased throughout the decellularization process, the mean perfusion pressure dropped to approximately 50 mm Hg by the time of the final rinses. Sixteen porcine hearts have been successfully decellularized using the method described.
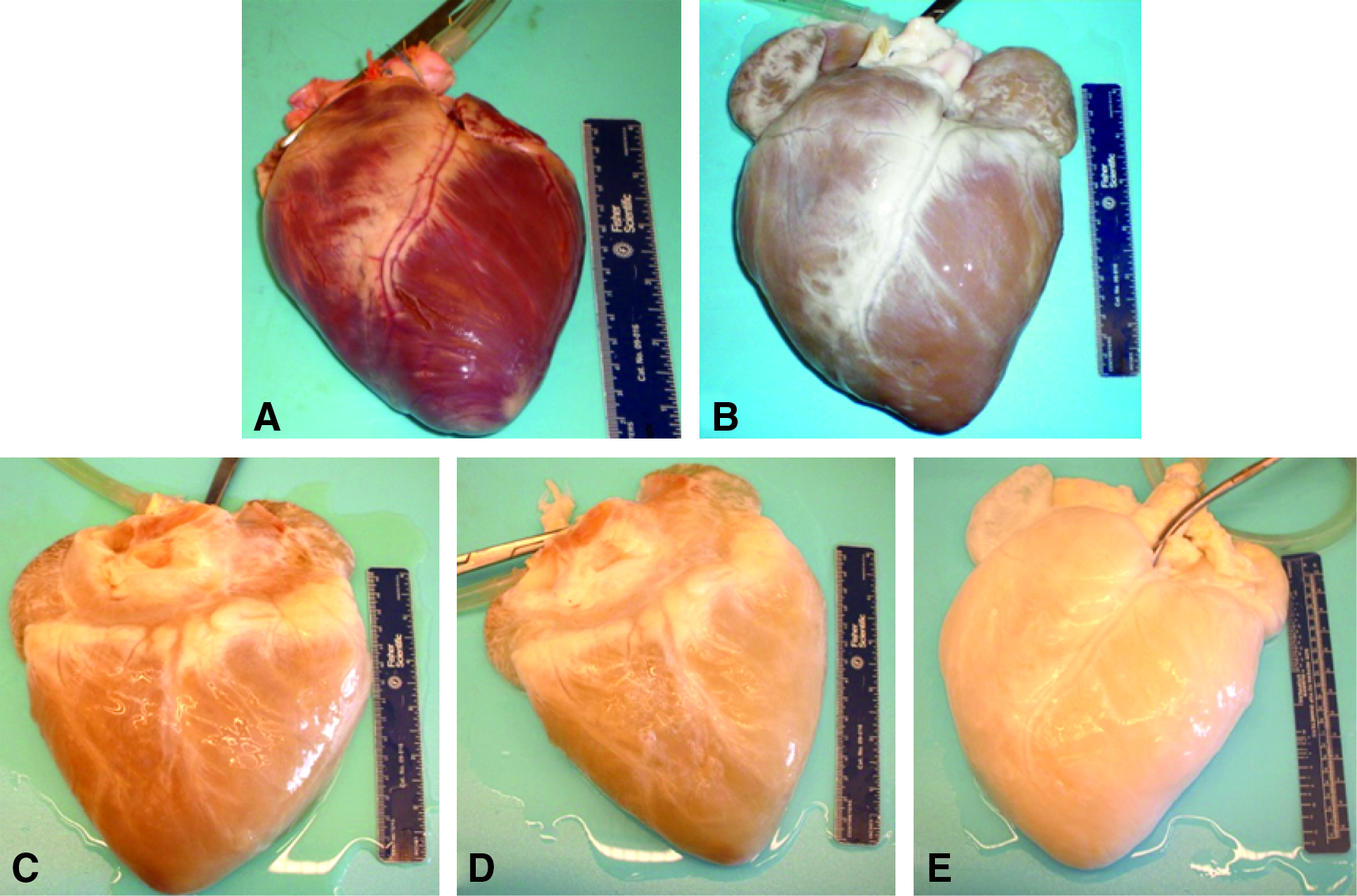
Representative images of the gross appearance of intact porcine hearts subjected decellularization by retrograde perfusion. (
Immunohistochemistry
Hematoxylin and eosin and 4′,6-diamidino-2-phenylindole showed no visible cell nuclei or double-stranded DNA at 40 × and 200 × magnification, respectively, whereas the native heart showed dense cellularity (Fig. 2). Movat's Pentachrome staining of the C-ECM showed the absence of muscle cells after decellularization (Fig. 3). Movat's and Herovici's staining showed the presence of collagen type I and III as well as elastin, with particularly dense collagen structure localized at the epicardium and endocardium (Figs. 3 and 4). The basement membrane structures present within native ventricle tissue and C-ECM were identified by the positive staining for collagen IV, a basement membrane component, on the endocardium, myocardium, and coronary arteries (Fig. 5).

Representative photomicrographs showing no nuclear staining after perfusion decellularization. (

Movat's Pentachrome photomicrographs of (

Herovici stain of C-ECM. (

Collagen IV staining of (
Scanning electron microscopy
The native and C-ECM epicardium and endocardium both showed a dense collagen layer with topographic variances. The native ventricle has a densely cellular myocardium, whereas the C-ECM shows a more open configuration within the myocardium. Intact vascular matrix was evident throughout the C-ECM. Figure 6 shows a 0.5-mm-diameter coronary artery in the cross-sectional view. No cells were visible in any of the SEM samples for C-ECM, but they were apparent in the native samples (Fig. 6).

Scanning electron microscopy of native and lyophilized C-ECM from the LV. (
DNA quantification
Quantitative analysis of DNA content within the C-ECM with the Pico Green assay showed a significant decrease in the amount of DNA compared to the DNA present in the native ventricles (0.66 ng DNA/mg sample vs. 8.48 ng DNA/mg sample [p = 0.014]). This value represents a 92% decrease in the amount of the double-stranded DNA found in the tissue as a result of decellularization (Table 1). The ethidium bromide gel showed no DNA bands associated with the decellularized C-ECM, whereas the native ventricle showed a large band above 1500 bp (Fig. 7).
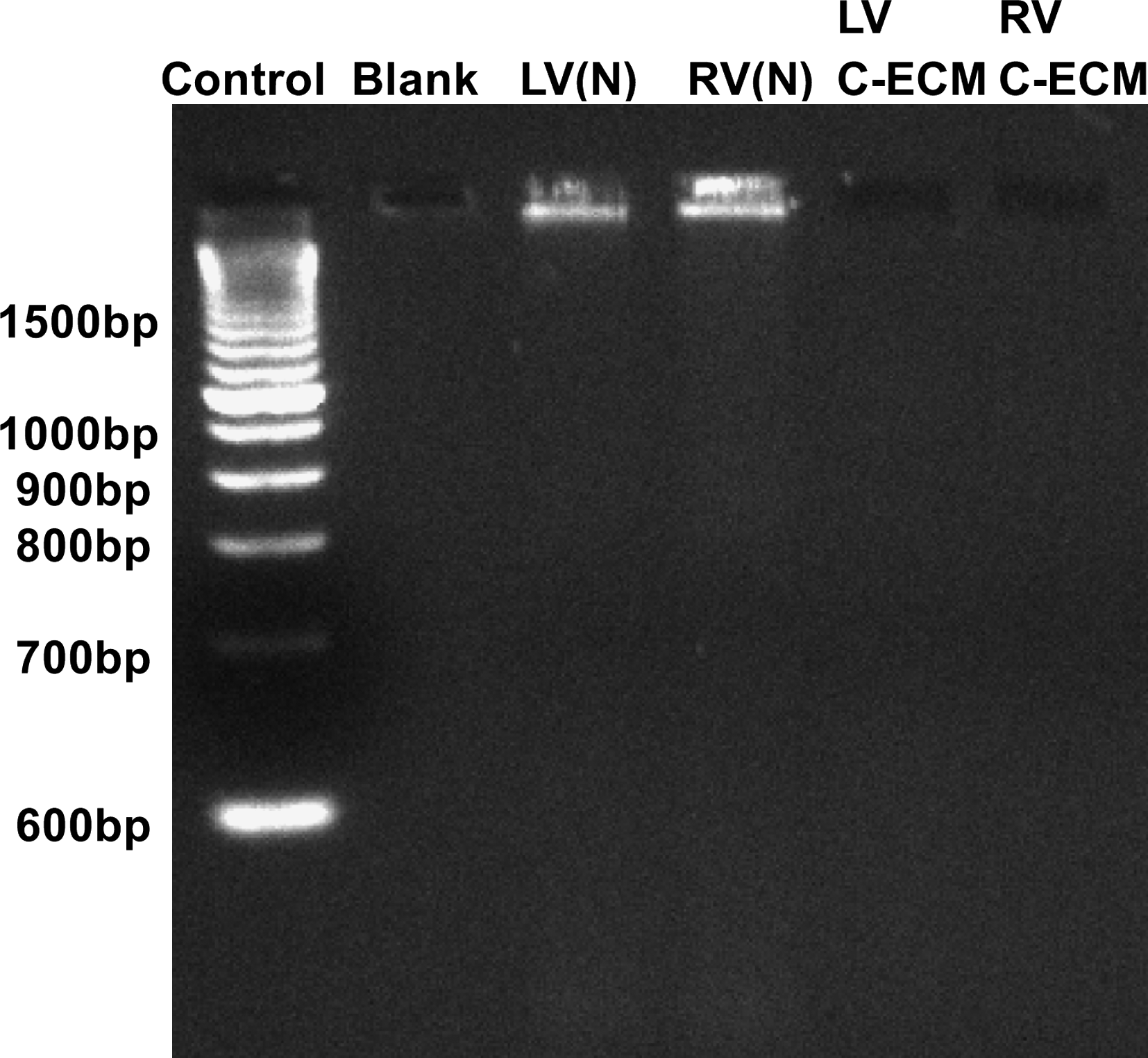
DNA fragment size as determined by ethidium bromide gel. (N), native. RV, right ventricle.
Values are normalized to 1 mg lyophilized sample.
GAG, glycosaminoglycan; C-ECM, cardiac extracellular matrix; STDV, standard deviation.
GAG and elastin quantification
The amount of GAGs and elastin in C-ECM was not different from that measured in the native ventricle tissue (p < 0.05) (Table 1).
Ball burst mechanical testing
The maximum force or extension at maximum force of C-ECM for the respective ventricle was not different from the native ventricle (p < 0.05), although the average extension at maximum force was greater for the native ventricles (Table 2). The native ventricles showed multiple subfailure peaks that were associated with failure of layers within the tissue, whereas the C-ECM showed a smoother curve consistent with rotation and extension of fibers that likely occurred as a result of dilation of the heart during decellularization (Fig. 8). Failure was observed as the ball penetrated through the epicardium in all groups (Fig. 8).
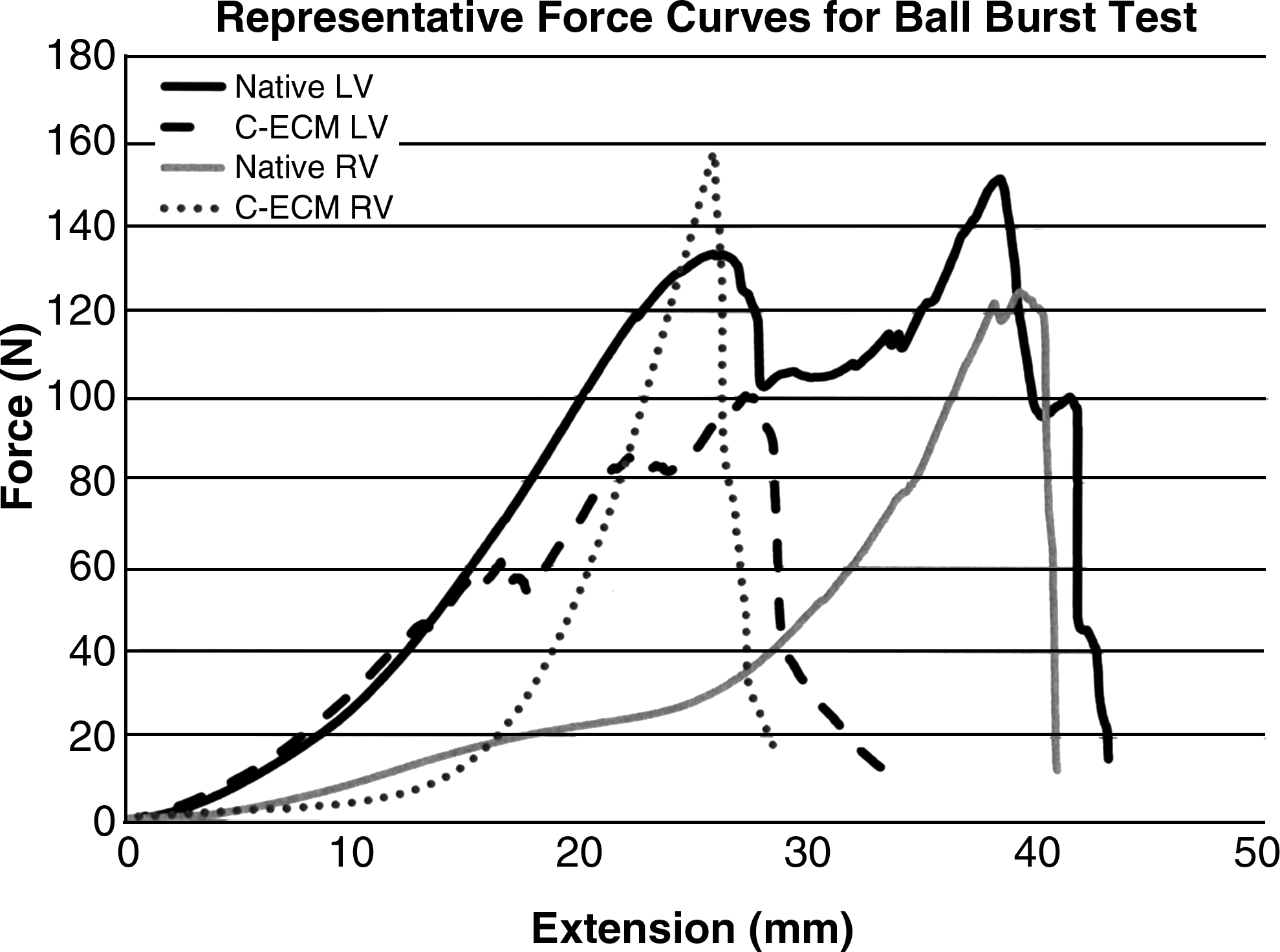
Graph of representative force curves for ball burst test.
LV, left ventricle; RV, right ventricle.
In vitro cell seeding
C-ECM sheet supported the formation of organized CM sarcomere structure, as indicated by striations of the α-actinin fibers. 30 The α-actinin fibers showed no global preferred orientation on the scaffold. CMs were the primary population of cells observed on the C-ECM, as only a few cells solely stained positive for β-tubulin (Fig. 9).
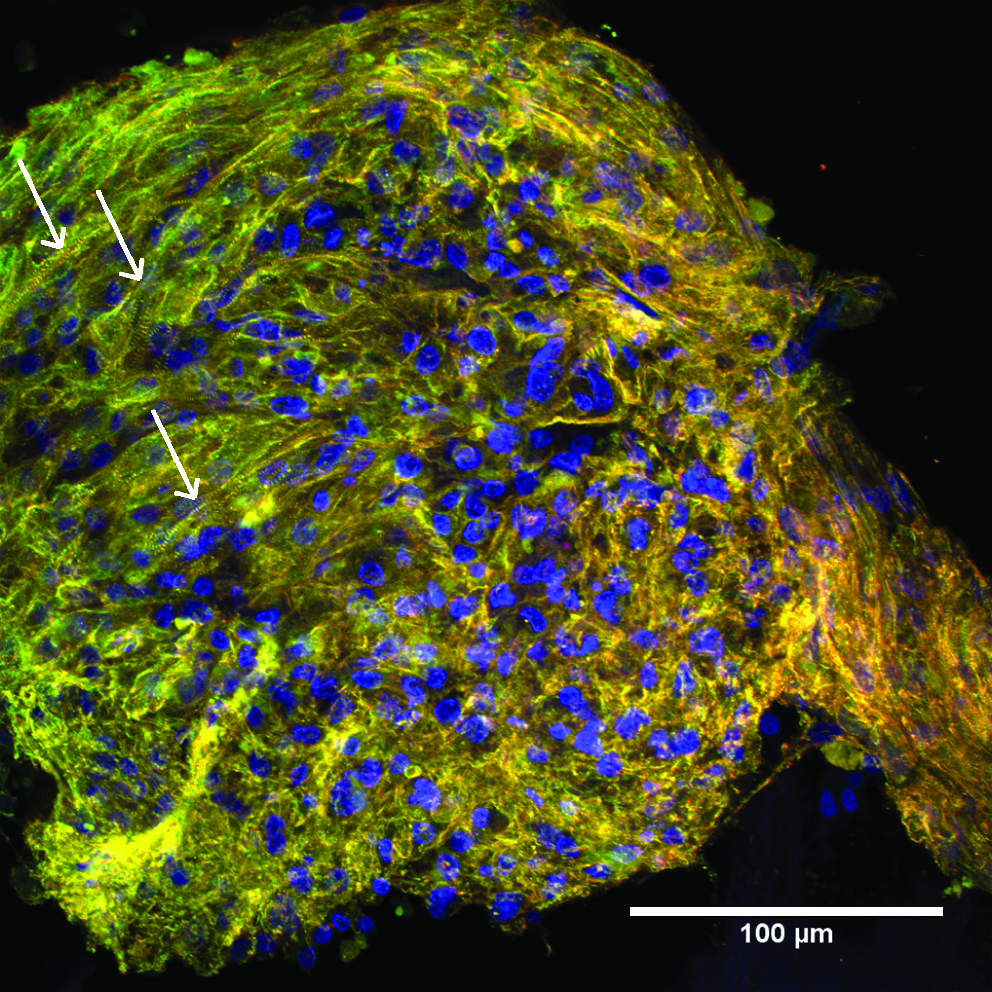
Cardiomyocytes (CMs) cultured on ECM for 4 days on endocardial surface of C-ECM (400 ×). CMs are identified by positive α-actinin staining and positive β-tubulin staining (green–yellow). Non-CMs are recognized by negative α-actinin staining and positive β-tubulin staining (red). Arrows denote example of striations.
Discussion
The present study described a reproducible and time-efficient decellularization technique for the intact porcine heart. The technique utilized retrograde aortic perfusion with successive hypertonic, hypotonic, enzymatic, acid, and detergent solutions to maximize the distribution of chemicals throughout the tissue, maximize the disruption of cells, and minimize the damage to the ECM. The protocol took less than 10 h to complete, and effectively removed DNA from the tissue to levels comparable to other commercially available ECM products.24,31
It is important to limit the tissue exposure time and concentration of trypsin, Triton® X-100, deoxycholic acid, sodium dodecyl sulfate, peracetic acid, or other commonly used decellularizing agents, as each solution can have a disruptive effect on the ECM structure while removing the cellular components. 32 By appropriate utilization of the series of reagents described in the present method, the decellularization time was significantly reduced from the time reported for decellularization of porcine heart valves, sliced porcine heart, and intact rat hearts.19,33–35 A previous study on the decellularization of heart valves with trypsin and Triton X-100 found that collagen and elastin structure were severely disrupted and that stable suture lines for anastomosis could not be formed. 36 In the present study, the IHC, SEM, and mechanical data showed intact collagen structure. The complex 3D architecture of the heart was maintained, including vasculature and basement membrane structures. The biaxial rupture strength of the C-ECM was not different from the native tissue; this result is especially impressive as the dry weight of the C-ECM samples tested was approximately 1/3 of the native samples (data not shown).
The composition of the resulting C-ECM was largely preserved, with similar GAG (essential for water retention) and elastin content compared to native ventricles. The C-ECM was shown to be a suitable substrate for cardiac cell attachment with maintenance of CM phenotype. Although the method described is specific to cardiac tissue, the technique could be systematically modified for decellularization of other tissues and organs.
As shown by Ott et al. on a smaller scale, an intact decellularized porcine heart could serve as a scaffold for cardiac organ engineering. 19 The remnant vasculature serves as a conduit for distribution of select cells throughout the tissue and for delivery of cell culture media to support cell viability and differentiation. The presence of an organ-specific ECM may also provide signals to seeded cells to enhance their differentiation and function.37,38 Additional work will be required to determine the optimal culture conditions for recellularization of a porcine heart with appropriate cellular diversity and restored tissue function before transplantation of a tissue-engineered heart becomes feasible.
Summary and Conclusion
The porcine heart can be efficiently decellularized using aortic retrograde perfusion. The resultant natural biodegradable scaffold maintains much of its complex structure and composition. While there is much work to be done, the methodology described herein provides a useful step to fully realizing an engineered complex organ. With slight modification, it is conceivable that the perfusion decellularization method described herein could be used to decellularize other organs, such as the liver, kidney, or lung.
Footnotes
Acknowledgment
Partially supported by the NIH-NHLBI training grant (T32-HL76124) entitled “Cardiovascular Bioengineering Training Program.”
Disclosure Statement
No competing financial interests exist.