
Abstract
Human mesenchymal stem cells form the supportive structure in which the functional cells of a differentiated tissue reside. We describe the creation of ectopic niches within polyurethane scaffolds coated with human mesenchymal stem cells. When implanted subcutaneously in NOD/SCID mice, these niches supported engraftment of primary human acute myeloid leukemia cells. The scaffolds showed vascularization and presence of osteoclasts and adipocytes, suggestive of an organizing human bone marrow microenvironment in the murine host. The chemokine stromal-derived factor-1 (SDF-1 or CXCL12) and its receptor CXCR4 are critical for homing and migration of acute myeloid leukemia. We found that a CXCR4 antagonist could disrupt homing to the ectopic niches, possibly by modulation of the mesenchymal stroma. We believe that these scaffold niches provide a new and powerful tool to study the leukemia stem cell microenvironment and may be useful for identification of novel drug targets.
Introduction
In normal bone marrow, two components have been formally described to establish niches: the osteoblast niche and the vascular sinusoid niche.6–8 The complex bone marrow microenvironment comprises many other cells, for example, macrophages, adipocytes, chondrocytes, and fibroblasts. The common adult stem cell precursors of these cell types are the mesenchymal stem cells (MSC). 9 Throughout the body, MSC form the supportive structure in which the functional cells of a differentiated tissue reside, and tissue repair derived from MSC is currently studied in synthetic niches in vivo, serving as scaffolding for formation of new tissue.10–12 In addition, MSC have great potential as a source for innovative cellular therapies. 13
In this article, we describe the creation of a physiological bone marrow niche in three-dimensional (3D) bioengineered scaffolds coated with adult human MSC. The gold standard for cell biological imaging has been two-dimensional tissue culture. However, a spatial distribution of cells within a 3D matrix is critical to mimic the complex cellular organization of the bone marrow microenvironment. The xenotransplant NOD/SCID mouse repopulation assay is the current model to assess clonogenicity of human leukemic stem cells, but one of the major restrictions of this assay is that human leukemia cells need to engraft in a murine bone marrow environment. We bypassed this hurdle by subcutaneous implantation of human MSC-coated scaffolds in NOD/SCID mice, creating an ectopic permissive human microenvironment. These scaffold niches, after implantation, enabled the engraftment and maintenance of inoculated human AML cells. We further explored the role of the SDF-1/CXCR4 axis in the promotion of the leukemia niche, suggesting possible targets for disruption.
Materials and Methods
Isolation of MSC
Fresh human bone marrow samples were obtained from orthopedic surgery after informed consent (Tissue Donation Program of The Feinstein Institute). The buffy coat containing the mononucleated cells was isolated by Ficoll-density gradient separation (Stem Cell Technologies). Mononucleated cells were plated at a low density in six-well plates in alpha-minimal Eagle's medium (MEM) culture medium (Lonza), 20% heat-inactivated fetal bovine serum (FBS) (Hyclone), 5 mM L-glutamine, and 100 units/mL penicillin/streptomycin (Stem Cell Technologies) for overnight adherence. After 24–48 h, the nonadherent cell fraction was discharged by rigorous pipetting. At day 5, single-cell-derived mesenchymal colonies were processed for immunohistochemistry (Vectastain ABC kit; Vector Laboratories). MSC were stained for CD90/Thy-1 (Mouse anti-human mAb, Clone 5E10; BD Pharmingen), CD105 (Mouse anti-human mAb, clone 266; BD Pharmingen), and CD146 (Mouse monoclonal [P1H12]; Abcam). Cells were also labeled for flow cytometry with CD34-APC (Miltenyi Biotec), CD105-FITC (R&D Systems), CD45-FITC, CD90-PECy5, CD146-PE, and CXCR4-PE (BD Pharmingen), including matching isotype controls.
Preparation of MSC-coated scaffolds in vitro
Polyurethane scaffold test discs (10 mm diameter and 1.5 mm thickness, pore size 300–400 μm; kindly provided by Biomerix in Material Transfer Agreement) were placed inside a cloning cylinder (Millipore Corporation), one cylinder in each well of a 24-well plate (Falcon, BD) and primed with alpha-MEM culture medium (Lonza), 20% heat-inactivated FBS (Hyclone), 5 mM L-glutamine, and 100 units/mL penicillin/streptomycin (Stem Cell Technologies) in presence of SDF-1/CXCL12-10 ng/mL (R&D Systems). Inserted scaffolds were seeded with freshly harvested human bone-marrow-derived MSC (1 × 107 cells/disc) and cultured in alpha-MEM/20% FBS with SDF-1 (10 ng/mL; R&D Systems) at 37°C, 5% CO2 in humidified air. After 24–48 h, the scaffolds were flushed rigorously with phosphate-buffered saline (PBS) to remove any nonadherent cells and fed with fresh medium for continued tissue culture for 3 days.
Ectopic MSC-coated scaffold implantation in NOD/SCID mice
Polyurethane scaffolds (10 × 1.5 mm, pore size 300–400 μm; Material Transfer Agreement with Biomerix), coated in vitro with human bone-marrow-derived MSC as described above, were implanted in a subcutaneous pocket on the dorsum of nonirradiated NOD/SCID mice (n = 40) (Jackson Laboratory) according to an approved Institutional Animal Care and Use Committee (IACUC) animal protocol. Empty scaffolds (without MSC seeding) were implanted as negative controls.
Imaging of the human leukemic stem cell niche
Primary AML cells were characterized by flow cytometry as 92.1% ± 0.9% CD45 positive and 83.2% ± 2.9% CD34 positive. A total of 1 × 107 thawed leukemia cells were injected into the scaffold in situ of the mice that hosted the ectopic MSC-coated scaffold on their back (n = 40 mice) and analyzed for engraftment. In one experiment (n = 4 mice), primary AML cells were injected in the retroorbital venous plexus to evaluate homing of the human AML cells to the synthetic hematopoietic niche. In some in vivo experiments, scaffold-hosting NOD/SCID mice were divided in two treatment groups: one group received SDF-1/CXCL12 (10 ng/mL) by biweekly in situ injection in the MSC-coated scaffold and the other group received AMD3100 (10 μM), a CXCR4 antagonist. Control mice received PBS buffer only. Each experiment was performed in triplicate, with two mice per experiment in each treatment group and one mouse as a negative PBS control.
After 1, 4, 8, and 16 weeks, the scaffolds were harvested from the mice, fixed, paraffin-embedded, and stained for immunohistochemistry. Briefly, harvested scaffolds were fixed in formalin solution 10%, neutral buffered (HistoPak-StatLab Medical Products), and paraffin embedded and cut on slides. After initial deparaffinization, the slides were subject to 8 min antigen retrieval (citrate buffer pH 6.6) and incubated with the primary antibody for 32 min (Benchmark XT automated stainers—mouse monoclonal anti-CD45 antibody [Ventana Clone RP2/18] or anti-Ki-67 rabbit monoclonal primary antibody [Ventana]). Antibodies used were human specific with no murine cross-reactivity. Negative control stains (no primary antibody, secondary antibody only) were performed to rule out nonspecific staining. Imaging was done by inverted microscopy (Zeiss-Axiovert) and photographed by a Nikon digital camera.
Multiple slides per scaffold were analyzed for imaging by immunohistochemistry and light microscopy. Negative controls included empty scaffolds (no MSC-coating or AML cells inoculation), PBS-injected scaffolds (no SDF-1/CXCL12 or AMD3100), and immunohistochemistry without primary antibody present.
For electron microscopy, MSC-coated scaffolds were fixed by immersion in 2%, 0.05 M cacodylate buffered, glutaraldehyde, postfixed in OsO4, dehydrated in a graded series of ethanol, and prepared for electron microscopic study by standard methods. Appropriate cellular areas were identified on 1 μm plastic sections by light microscopy and their ultra structure was evaluated using a JEOL JEM 100CXII transmission electron microscope.
Results
First, bone-marrow-derived MSC were phenotypically characterized by flow cytometry and immunohistochemistry. MSC were adherent to plastic when maintained in culture and stained positive for CD90, CD105, and CD146 but negative for the hematopoietic lineage markers CD34 and CD45. A small subfraction of MSC expressed CXCR4. These studies confirmed that MSC isolated from human bone marrow samples and used in our experiments met criteria proposed by the Mesenchymal and Tissue Stem Cell Committee of the International Society of Cellular Therapy to define human MSC.11,14
We next determined whether bone-marrow-derived MSC seeded in polyurethane scaffolds could establish an ectopic human bone marrow environment in NOD/SCID mice (Fig. 1). We hypothesized that the spatial distribution of cells within a 3D matrix (Fig. 1A) would be critical to allow for the complex cellular organization of the bone marrow microenvironment to develop, as well as for retention of the cells at the intended site. When the scaffold was coated in vitro with MSC in the presence of SDF-1/CXCL12, we were able to show cell adhesion and cell division of MSC (data not shown). We then asked whether the MSC-coated scaffolds could support MSC survival after implantation in NOD/SCID mice (n = 6). Analysis of the scaffolds 1 week after implantation revealed vascularization in all scaffolds, while the empty control scaffolds without MSC-coating had only growth of reticular fibers with signs of murine foreign body reaction at the borders of the scaffold (Fig. 1B, C). Eight weeks later, the MSC-coated scaffolds showed the presence of adipocytes, blood vessels, and osteoclasts, suggestive of stromal tissue components creating an ectopic human bone marrow microenvironment (Fig. 1D, E). The scaffolds were well tolerated in the immunodeficient NOD/SCID hosts, without infection or ulceration.

MSC-coated scaffold creates an ectopic human bone marrow microenvironment in NOD/SCID mice. (
Second we asked if the ectopic human bone marrow environment in the implanted MSC-coated scaffolds could support leukemic hematopoiesis. Implanted scaffolds were inoculated with primary AML cells in 40 mice and analyzed for engraftment by diaminobenzidine (DAB) peroxidase staining for human CD45 (common leukocyte antigen, present on leukemic blasts) after immunofixation of harvested scaffolds at weeks 1, 4, and 8 (Fig. 2A) and at 4–5 months (Fig. 2B-C). Human CD45-positive AML cells resided in the perivascular space of the scaffold stroma, 1 week after in situ scaffold injection (Fig. 2A-1). At 4 and 8 weeks, AML cells engrafted, survived, and proliferated in the ectopic niches (Fig. 2A-2, A-3), while in the empty control scaffolds that were not coated with MSC before implantation, no stromal bone marrow elements developed and the AML cells did not survive (Fig. 2A-4). After 4 months, large subcutaneous tumors (2–5 cm diameter) developed at the implantation site (Fig. 2B-1), while the mice did not appear sick or debilitated. The tumors consisted of proliferating human leukemic cells that stained positive for human Ki67, CD45, and CD34 by histology (Fig. 2B-2, B-3, and B-4). Flow cytometric analysis of the tumor validated that about 86.3% (±1.8%) of the cells were positive for human CD45, confirming that the tumors were derived of human cells and not of murine origin (Fig. 2C). Antibodies used in immunohistochemistry as well as for flow cytometry were human species specific, without murine cross-reactivity. In one experiment (n = 4 mice), the AML cells were injected in the retroorbital venous plexus of the mice. After 4 weeks the AML cells were present in the human synthetic niches, but none were found in the murine femurs or spleens, suggesting preferential homing of the human leukemia to the human microenvironment (data not shown).
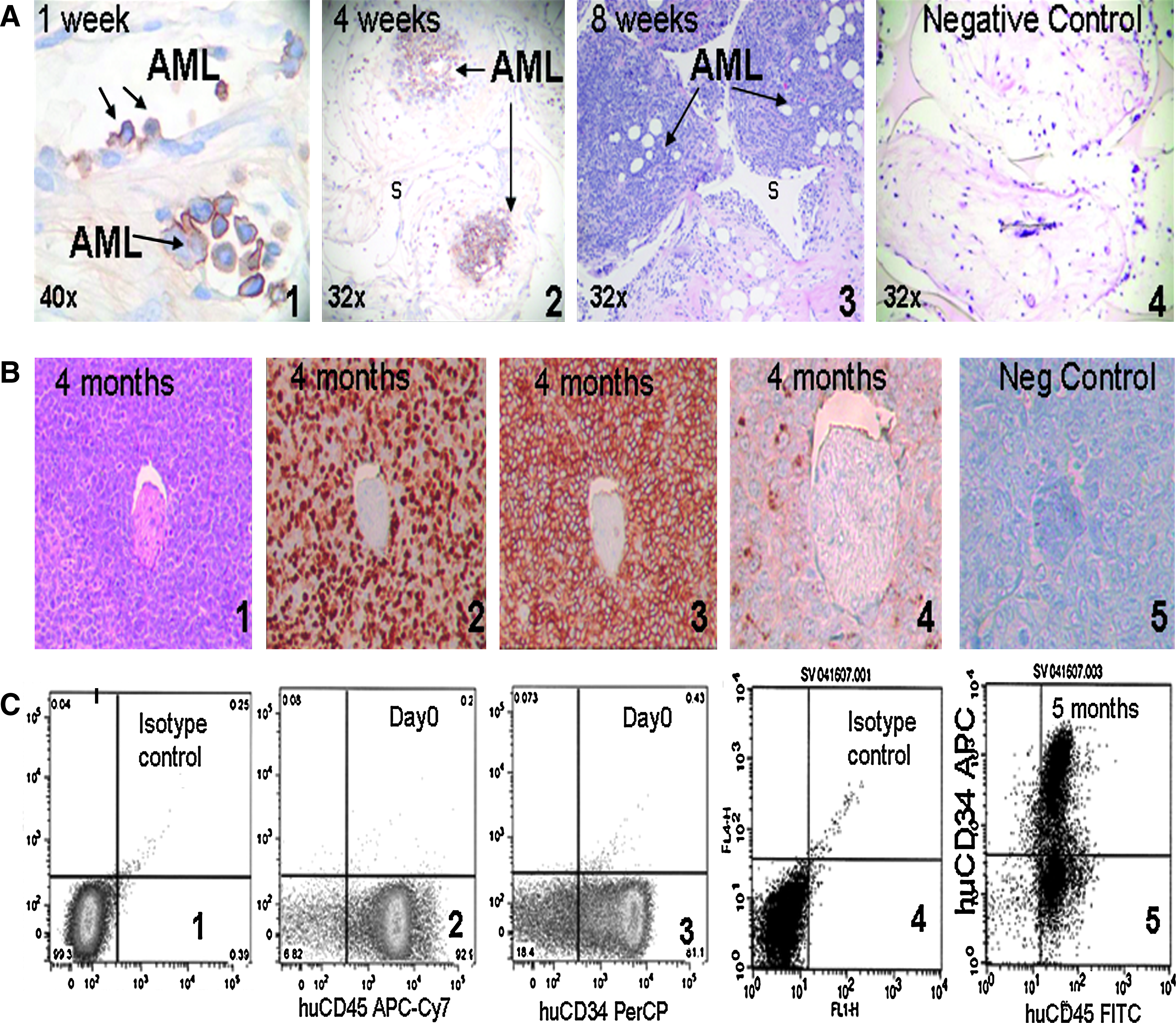
Leukemia progression in the scaffold niche in vivo. (
At 4 months, there was evidence of hematogenous seeding of the human AML cells to the murine femoral bone marrow, liver, and spleen without local invasiveness of the tumors at the implantation site (Fig. 3A–C). Hematogenous seeding appeared to be an adaptive process of the AML blasts, since at 4 and 8 weeks, murine femurs, livers, and spleens did not contain human AML cells as determined by immunohistochemistry (data not shown).

Hematogenous spread of human leukemia at 4 months. (
Within the hematopoietic osteoblast niche of the bone marrow, hematopoietic stem cells adhere to osteoblasts by developing long, tentacle-like projections called pseudopods. 15 In our scaffolds, similar cell-to-cell (AML cells/MSC) interactions were observed at a single-cell level. AML cells anchored intimately to MSC via pseudopod formation (EM 6700 ×). The granular cytoplasm of the AML cell appeared to envelop an adjacent MSC (EM 27,000 ×—arrows), implying that a part of the AML cytoplasm migrated underneath the MSC (Fig. 4).
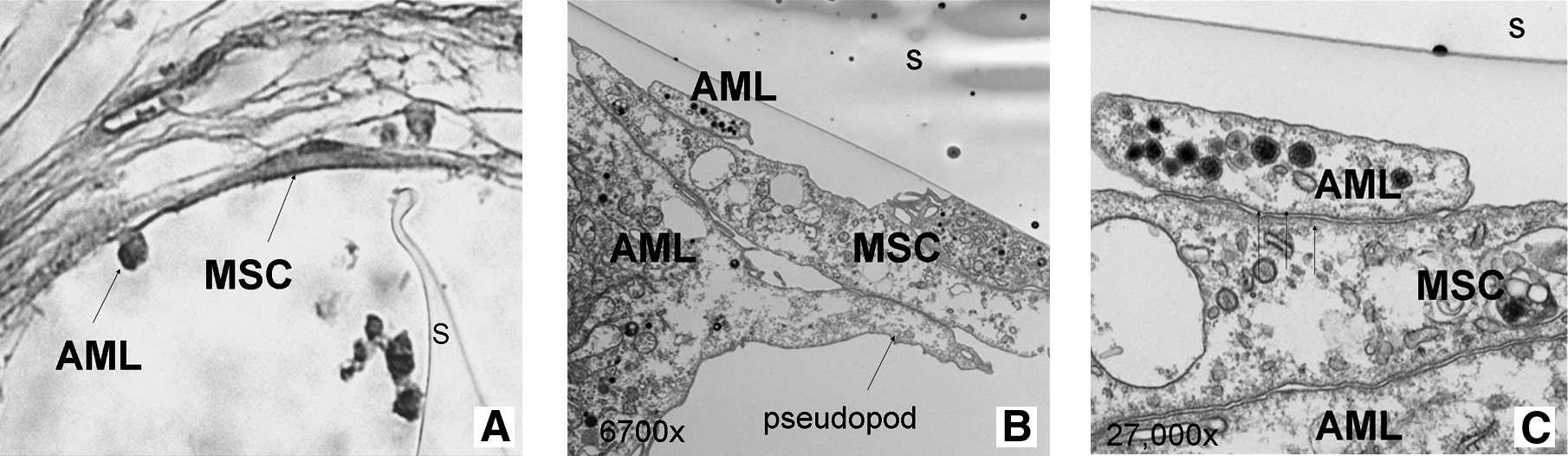
Ultrastructural evaluation of the leukemic stem cell niche: cell-to-cell contact via pseudopods. (
Ki67 is a histochemical marker for cell mitosis. The AML cells adherent to MSC remained Ki67 negative, whereas intravascular and nonadherent AML cells stain positive for Ki67 (Fig. 5), implying that the MSC niche might provide a protective milieu for dormant AML cells. The picture shown in Figure 5 is a representative slide of many images taken of 20 different scaffolds examined by the clinical pathologist and the first author. In all cases, adherent cells were Ki67 negative and nonadherent Ki67 positive.
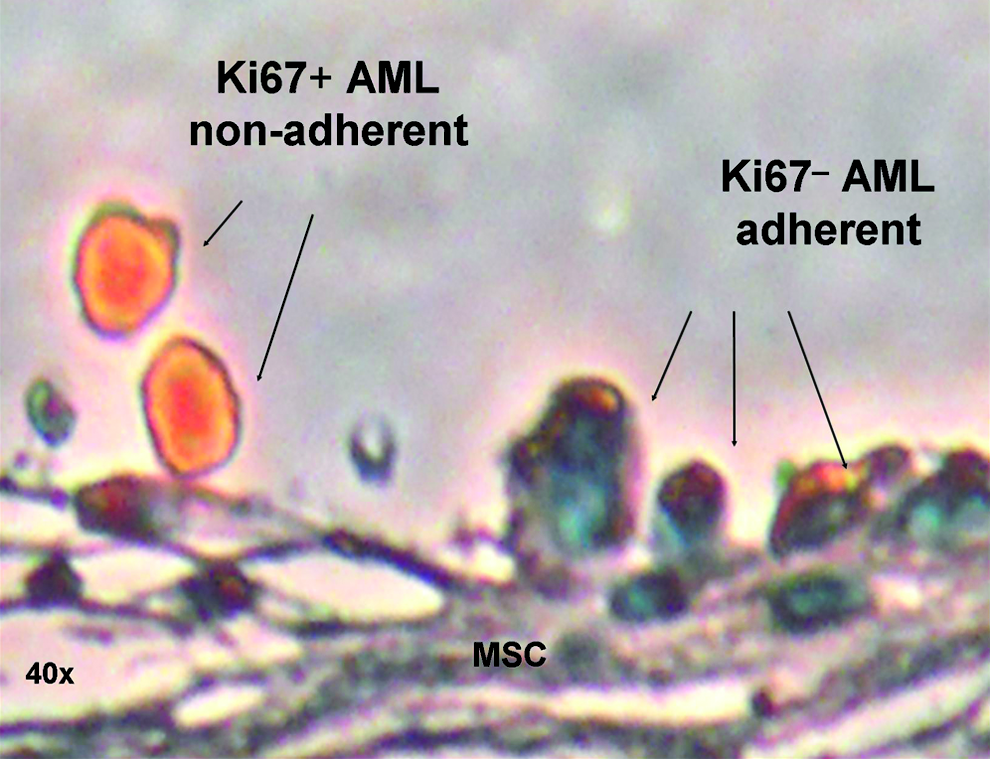
Leukemic cells are quiescent in the niche sanctuary. Human Ki67 stain (cell cycle marker, orange): representative example of nonadherent AML cells that are Ki67 positive (cell division), while adherent AML cells remain Ki67 negative as quiescent cells hiding in the niche. A total of 20 implanted scaffolds were examined by light microscopy.
We then asked whether the ectopic human bone marrow environment can create a leukemic niche as a tool for drug screening of a niche-disrupting agent. The interaction between the chemokine SDF-1/CXCL12 and its receptor CXCR4 plays a major role in leukemogenesis and leukemia progression. Studies on the CXCR4 antagonist AMD3100 in AML have revealed a regulatory role of CXCR4 in leukemia migration. However, the exact mechanism of AML cell engraftment via the SDF-1 (CXCL12)/CXCR4 axis is not fully understood. Therefore, we chose to study SDF-1 and AMD3100 in our ectopic leukemia niche model (Fig. 6). In three different experiments, scaffold-implanted mice were divided into two treatment groups: one group received SDF-1/CXCL12 (10 ng/mL) by biweekly in situ injection in the MSC-coated scaffold, and the other group received AMD3100 (10 μM). Control mice received PBS buffer only. Four weeks later, the SDF-1/CXCL12-treated scaffolds showed thick proliferation of the MSC stromal layer with multiple adherent AML cells present (Fig. 6A), whereas the AMD3100-treated scaffolds had a thin stromal lining that was disrupted at several points, leaving AML cells floating freely in proximity (Fig. 6B). The PBS control scaffolds showed a single layer of MSC with only a few AML cells attached (Fig. 6C). In long-term follow-up of these animals, both treatment groups developed subcutaneous leukemia tumors at the site of AML/MSC scaffold implantation, but in the AMD3100-treated mice the tumors appeared to be much smaller than those in the SDF-1/CXCL12-treated mice. The subcutaneous leukemic tumors did not show any sign of local invasiveness or tissue destruction.

The leukemic stem cell niche is regulated by the SDF-1/CXCR4 axis. (
Discussion
MSC have great potential for use in tissue engineering 11 and regenerative medicine. 16 We aimed to establish an ectopic human bone marrow environment, using MSC as the primary cell source. Bone marrow develops from the mesodermal layer during embryogenesis, and bone-marrow-derived adult MSC maintain the potential to differentiate into cells and tissues of mesodermal origin (i.e., adipocytes, cartilage, bone, and skeletal muscle). 17 As such, MSC progeny define various cells present in the bone niche, for example, osteoblasts, vascular lining cells, and fat cells. 18 Studies have shown that MSC aid in the regrowth of hematopoietic-supporting stroma. 19 Intramedullary transplantation of enhanced green-fluorescent-protein-marked human MSC into NOD/SCID mice resulted in a functional human hematopoietic microenvironment integrated in the bone marrow of the murine host. Enhanced green-fluorescent-protein-marked human MSC differentiated into myofibroblasts, bone marrow stroma cells, osteocytes in bone, bone-lining osteoblasts, and endothelial cells. 20 Bone-marrow-derived CD146-positive reticular cells also have been reported to create a hematopoietic microenvironment in heterotopic sites when transplanted subcutaneously in a xenograft transplantation model. 21 In addition, Matsuzaki and colleagues demonstrated that transplanted adult mouse bone marrow MSC could differentiate in hematopoietic niches of the murine bone marrow. 22
Scaffold design plays an important role and defines the functional properties of the implanted cells, such as cell migration and adhesion. 23 The Biomerix Reticulated Matrix is a biodurable, reticulated, elastomeric polyurethane scaffold, and the material has superior biostability compared to other polyurethanes that are currently approved for long-term implantation in humans. The scaffold consists of a reticulated architecture, defined as a porous structure with an interconnected network of pores (300–400 μm pore size, 10 mm diameter, and 1.5–2.0 mm thickness). We preferred the use of nonresorbable polyurethane material because it better maintains the 3D structure required to mimic the physiological bone marrow microenvironment. However, one of the problems with synthetic scaffolds is their hydrophobic quality. 11 We overcame this problem by priming the scaffolds in the MSC culture medium containing SDF-1. We hypothesize that the presence of SDF-1 promotes adhesion of the MSC to the polyurethane scaffold, which is necessary for successful coating.
In our in vivo study, 3D scaffolds were coated with MSC derived from human bone marrow samples, and the MSC-coated scaffolds provided the supportive network for a tissue-engineered murine model. The MSC-coated scaffolds supported adipocytes, osteoclasts, and blood vessels, mimicking a human bone marrow stromal network that enabled growth and maintenance of inoculated human AML cells. Control empty scaffolds, without human elements, showed only the in-growth of murine-derived reticular fibrous tissue. We suggest that our engineered scaffolds may offer many advantages in the study of the leukemia niche since they can be easily manipulated and removed for histologic and molecular analysis without disrupting the niche interactions. Histological assessment may characterize the pattern and distribution of cell-to-cell interactions within the heterogenous population of the stem cell niche. In addition, the implanted device creates a homologous niche wherein human stroma interacts with human leukemia cells, bypassing the restrictions of a conventional xenograft transplantation model. Finally, in situ injection of the inoculated cells can enable studies of the niche environment, circumventing the limitations of standard intravenous cell injections, including poor delivery and poor retention of cells at the intended site.
Animal models have been critical to the field of tissue engineering and regenerative medicine, and we plan further research to focus on unanswered questions. The cell–scaffold implant needs to be evaluated for scar formation and induction of an inflammatory response or rejection. Inoculated cells need to be monitored for survival, trafficking, and proliferation, and their differentiation fate (including tumorigenicity) needs to be examined. In our hands, the MSC-coated scaffolds did not elicit any inflammatory reaction or scar formation at the implantation site. In vivo exposure of the human MSC to the subcutaneous microenvironment of the mice appeared to induce their differentiation along the mesodermal lineage. The inoculated leukemia cells were evaluated for survival in the niche by harvesting the scaffolds at various time points to monitor leukemia engraftment, proliferation, and disease progression. The leukemia progressed within the ectopic human bone marrow environment of the scaffold for >2 months, and no circulating human AML cells were detected in the murine peripheral blood during that initiation period, suggesting the initial preference for growth of the human AML cells in the human microenvironment. However, after 4 months, there was hematogenous seeding of the human AML cells to the murine femoral bone marrow, liver, and spleen, without local invasiveness of the subcutaneous leukemic tumors that had formed. The adaptive pathways involved in this metastatic process will be a topic for future investigations.
We also tested our model to evaluate the CXCR4 antagonist, AMD3100, which disrupts the chemokine/chemoreceptor signaling axis of SDF-1 and CXCR4. Injection of SDF-1 or AMD3100 or PBS (control) showed a differential effect on the interaction of AML cells with the mesenchymal stroma in the niche. SDF-1 upregulated adhesion of AML cells to the stroma and induced hyperproliferation of the mesenchymal stroma compartment. In contrast, in the presence of AMD3100, the MSC stroma became ruffled and nonadherent, and most of the AML cells remained only loosely attached. The control PBS-injected scaffold showed a single layer of MSC lining with sporadic adhesion of AML cells. AMD3100 is known to mobilize leukemia cells, but our data imply that there is also an effect on the stromal compartment of the niche. By blocking the autocrine SDF-1 pathway of MSC, AMD3100 appears to disrupt the stromal layer and prevent leukemia—niche signaling. Likely, other signaling pathways are involved in this process and need to be identified in future experiments.
We suggest that our model may lead to further insights into stem-cell-to-niche signaling through the use of transcriptional profiling.24,25 We have been able to grow AML cells for 4 months in scaffolds, until subcutaneous tumors developed at the implantation site. Single-cell suspensions derived from the tumors after 5 months were analyzed by flow cytometry and confirmed to be 86.3% ± 1.8% human CD45 positive and 63.1% ± 6.0% human CD34 positive. (At day 0 before injection, AML cells were 92.1% ± 0.9% CD45 positive and 83.2% ± 2.9% CD34 positive.) This suggests maintenance of CD34 antigen expression on AML cells retained in the niches after 5 months. We plan further investigation of the dynamic process of quiescence, self-renewal, and differentiation by which the CD34-positive cells are maintained within the synthetic niche. Comparison of the genetic signature of the AML cells before and after niche exposure may reveal differences in genetic signature and identify new targets for therapy.
Finally, we believe that our strategy may be applicable beyond the leukemia stem cell niche described here. Repopulation of implantable scaffolds with different stromal cell–stem cell mixtures might provide information on a diversity of functional niches with long-term potential.
Footnotes
Acknowledgments
We would like to thank C. Mason, M. Keogh, and the orthopedic team at North Shore LIJ Health System, New York, for collecting anonymous human bone marrow samples via the Tissue Donation Program at The Feinstein Institute. The polyurethane scaffolds were kindly provided by Biomerix (Material Transfer Agreement). S. Teichberg assisted with electron microscopy. B. Steinberg (The Feinstein Institute) and K. Sakamoto (University of California at Los Angeles [UCLA]) improved the manuscript with valuable suggestions. S.R. Vaiselbuh is funded by a Saint-Baldrick's Research Grant and a Saint-Baldrick's Career Development Award. This research is also supported by a grant from the Pediatric Cancer Foundation.
Disclosure Statement
Sarah R. Vaiselbuh: No competing financial interests exists.
Morris Edelman: No competing financial interests exists.
Jeffrey M. Lipton: No competing financial interests exists.
Johnson M. Liu: No competing financial interests exists.