
Abstract
Human adipose-derived stem cells (ASCs) are attractive targets for genetic manipulation and cellular therapies. However, current methods of gene transfer are limited by lack of efficiency, toxicity, or safety concerns. Recombinant adeno-associated virus (rAAV) has been extensively assessed as a gene therapy vector and has an excellent safety profile. This study reports the efficient transduction of well-characterized, homogeneous cultures of human ASCs by rAAV serotypes 2, 5, and 6. Transduction with rAAV2 at high multiplicity of infection was associated with reduced cell viability; however, no adverse effect was seen with serotypes 5 and 6. A further increase in transduction efficiency was observed using a rAAV6 Y731F tyrosine capsid mutant. rAAV-transduced ASCs retained their adipogenic potential. Therefore, rAAV serotypes 2, 5, and 6 should be considered the vectors of choice for genetic manipulation of ASCs.
Introduction
Genetic modification of ASCs using lentivirus, retrovirus, and adenovirus vectors has been reported.3,7 Both lentiviral and adenoviral vectors are able to achieve significantly higher rates of transduction (>90%) than retroviral vectors, 7 but there are conflicting reports of the toxicity of adenoviral vectors in ASCs.3,7 Long-term gene expression after differentiation can be achieved with lentiviral vectors. 7 However, there are concerns that lentiviral and retroviral vectors may lead to insertional mutagenesis, limiting their potential clinical use.8,9 Nonviral transfer of genes to ASCs by transfection with lipofectamine or polyethylenimine has also been reported, but lower rates of transgene expression of 9% and 19%, respectively, were achieved. 10 Similar rates of transduction (11%) were achieved with nucleofection of “minicircle” DNA, whereas nucleofection of a plasmid achieved only 2.7% transduction; nucleofection was more toxic than transfection with lipofectamine. 6
Adeno-associated virus (AAV) is a helper-dependent parvovirus that is not associated with any pathology in humans. 11 Recombinant AAV (rAAV) has a coding capacity of ∼4.5 kbp and has received much attention as a vector for gene therapy applications as it encodes no viral proteins, is able to transduce both dividing and nondividing cells, and promote long-term gene expression in a variety of cells in vivo.11,12 Twelve serotypes and many additional clones of AAV have been described. 13 While integration of rAAV into the genome has been demonstrated, it occurs only at a low frequency with the vector DNA persisting in episomal form. 14 Previous studies have shown that rAAV is able to transduce murine adipocytes 15 and bone marrow-derived MSCs in mice, 16 rats, 17 and humans.18–24 However, the ability of rAAV to transduce ASCs has not been investigated.
In this study we screened AAV and a wide range of AAV serotypes and clones for their ability to transduce highly characterized ASC cultures, and examined the potential of vector-transduced cells to differentiate.
Materials and Methods
Isolation and cell culture of ASCs
Isolation
Lipoaspirate was obtained from liposuction procedures from eight different donors in total, under a protocol approved by the appropriate local ethics committee. Lipoaspirate was harvested from the abdomen or thigh region using wet or tumescent liposuction. The fresh lipoaspirate samples were processed in a standardized manner based on methods originally described by Rodbell 25 and modified by subsequent authors.26,27 The lipoaspirate was washed twice with complete medium (CM) consisting of Dulbecco's modified Eagle's medium (DMEM) Nutrient Mix F-12 supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin/glutamine (PSG) (all from Invitrogen). The tissue was then digested in phosphate-buffered saline (PBS) with 0.075% collagenase type I (Gibco Life Technology) for 30–45 min at 37°C with intermittent shaking until digested. The samples were then centrifuged at 700 g for 10 min. The supernatant was discarded and the cell pellet resuspended in CM. The suspension was then filtered through 100 μm cell strainers (BD Falcon™; Becton Dickinson), the filtrate washed with CM, and centrifuged at 1800 rpm for 5 min. The resulting cell pellet was termed the stromal vascular fraction (SVF).
Cell culture
For routine cell culture, the isolated SVF was cultured in 75 cm2 flasks at an approximate density of 1.5×104 cells/cm2 in 12.5 mL of CM. Cells were cultured at 37°C with 5% CO2 and the medium was changed every 3–4 days. Cells were passaged once they reached confluence. TrypLE Express (Invitrogen) was used as cells were resistant to detachment with 0.5% Trypsin-EDTA (Invitrogen) (data not shown). Cells were split 1:3 at each passage. Cells were cryopreserved in FBS +10% solution of dimethyl sulfoxide. Experiments showed that SVF and ASC viability and phenotype were maintained for at least 3 months after cryopreservation (data not shown).
Adipogenesis assay
Adipogenesis was induced by 14 days of exposure to adipogenic media (AM), feeding fresh AM every 3–4 days. AM comprised CM supplemented with 0.5 mM isobutyl-methylxanthine, 1 μM dexamethasone, 10 μM insulin, 200 μM indomethacin (all from Sigma-Aldrich), and fatty acids.26–30 Human fibroblasts derived from dermis were used as negative controls. After 14 days of culture in AM, cells were fixed with 3% paraformaldehyde and incubated for 1 h under Oil Red O stain (Sigma-Aldrich) to stain intracellular lipid.
rAAV production
rAAV was produced by transient triple transfection of HEK 293 cells with polyethylenimine as previously described.31,32 The three transfected plasmids were (1) pAM/CAG-eGFP-WPRE-bGHpA (Supplementary Fig. S1; Supplementary Data are available online at
Alternatively, the rAAV6 tyrosine capsid mutants and wild-type rAAV6 were purified by a modification of a previously described method.33,34 Briefly, cells were harvested in 150 mM NaCl and 20 mM Tris pH 8.0, and incubated for 1 h at 37°C with 0.5% sodium deoxycholate (Sigma-Aldrich) and 50 U/mL benzonase endonuclease (Sigma-Aldrich). The lysate was clarified by centrifugation, loaded on a HiTrap Heparin HP 1 mL column (Sigma-Aldrich), washed with 30 mL of 150 mM NaCl and 20 mM Tris pH 8.0, and eluted in a step gradient of NaCl. The 300 mM and 500 mM NaCl fractions were pooled, concentrated with a 100K MWCO Amicon Ultra Centrifugal concentrator (Millipore), washed with PBS, and filtered through a 13 mm 0.2 μm syringe filter (Acrosdisc; Pall). Aliquots of virus were stored at −80°C before use.
Viral genome copies per mL (vg copies/mL) were determined by quantitative real-time PCR as previously described. 31 In addition, transduction units/mL (TU/mL) were determined by infection of HeLa cells and detection of eGFP-positive cells by flow cytometry as previously described. 32 All vectors produced were able to transduce HeLa cells. 31
Transduction of ASCs by rAAV
ASCs were seeded at 2×104 cells per well in a 12-well plate the day before infection. On the day of infection the medium was replaced with 500 μL of serum-free DMEM + 1% PSG, and the cells infected at a multiplicity of infection (MOI) of 105 viral genome copies per cell unless otherwise stated. After 3 h the medium was replaced with CM, and the cells were incubated for 72 h at 37°C/5% CO2 before analysis by flow cytometry.
Transfection of ASCs using lipofectamine
As a positive control, ASCs were transfected with pAM/CAG-eGFP-WPRE-bGHpA using lipofectamine 2000 (Invitrogen) as per the manufacturer's instructions and as previously described. 10 Cells were analyzed by flow cytometry at 72 h post-transfection.
Flow cytometry
Cells were harvested with TrypLE and filtered through 100 μm meshes (BD Biosciences) before incubation with antibodies. Fluorophore-conjugated antibodies to the following antigens were used. The following were obtained from BD Biosciences: CD13-APC (catalogue #557454), CD29-APC (#559883), CD34-PE (#348057), CD34-PerCP (#559881), CD44-APC (#559942), CD49d-APC (#559881), CD49e-PE (#555617), CD54-APC (#559771), CD59-FITC (#555763), CD73-PE (#550257), CD146-PE (#550315), and CD166-PE (#559263). The following was obtained from Immunotech/Coulter: CD51-FITC (#1855). The following were obtained from Serotec: CD31-Alexa647 (#MCA1738A647), CD38-FITC (#MCA1019F), CD45-PE (#MCA87PE), CD62e-FITC (#MCA1969F), CD90-FITC (#MCA90F), CD105-Alexa488 (#MCA1557A488), and CD106-PE (#MCA2237PE). Cells were labeled with secondary antibodies anti-mouse IgG2a Alexa Fluor 488 (A21131), anti-mouse IgG2b Alexa Fluor 633 (A21146) (both from Invitrogen), and anti-mouse Ig FITC goat F(ab')2 (F0479, from DAKO Cytomation). All cells were stained with the viability stain 7-AAD (BD Biosciences) or propidium iodide (Invitrogen). Cells were analyzed using a FACSCalibur (BD Biosciences). In some experiments eGFP+ cells were sorted by fluorescence-activated cell sorting (FACS) using a FACS Vantage (BD Biosciences).
Cell counting by flow cytometry
In some experiments unlabelled caliBRITE beads (BD BioSciences) were included to control for cell recovery. At the time of seeding ASCs, an equal number of beads were added to the culture. After infection with rAAV, as outlined below, beads and ASCs were enumerated by flow cytometry; beads and ASCs could be readily distinguished by their forward scatter/side scatter profile (Supplementary Fig. S2). The number of ASCs was calculated by the following formula:
where Nbr is the number of beads recovered, Nbs is the number of beads added to the sample, Ncr is the number of ASCs recovered, and Ncs is the number of ASCs seeded.
Statistical analysis
Viable ASCs, as determined by forward scatter/side scatter profile and negative staining with 7AAD or propidium iodide, were considered positive for eGFP if fluorescent intensity was >3 times the maximum autofluorescence. Each experiment was performed three times in triplicate, and the average of technical replicates compared by a one-way analysis of variance (ANOVA), using SigmaPlot for Windows Version 11.0 (Systat Software, Inc). If a statistically significant difference was detected between groups (p<0.05), then pairwise multiple comparison was performed by the Holm-Sidak method. Where indicated, probability testing was performed on log10-transformed data.
Results
Characterization of ASC cultures
ASCs were purified as reported26–30 by culturing adherent cells within the SVF from human adipose tissue single-cell suspensions after removal of mature adipocytes by differential sedimentation. To ensure that transduction experiments were performed on a homogenous population of ASCs, cell surface phenotype was assessed in the SVF and following passage. Flow cytometry revealed that SVF contained more than one population, as defined by CD34, CD31, and CD45 (Figs. 1 and 2B), consistent with previous reports.35–37 While these populations were still present at the time of first passage (P0), cells were almost uniformly CD31− CD45− by passage 2 (data not shown), consistent with a previous report. 36 The effect of passage on other phenotypic markers was also assessed. Although most cells in the SVF and P0 culture were CD73+, CD73− cells were present; by passage 4 CD73 expression had been upregulated and no CD73− cells were seen (Fig. 1), suggesting that CD73 is a good marker to distinguish CD73+ ASCs from contaminating CD73− cells in the SVF. Other changes in cell phenotype with serial passage included reduced expression of CD34 and increased expression of CD105 and CD166 (Fig. 1), consistent with previous reports.35,36
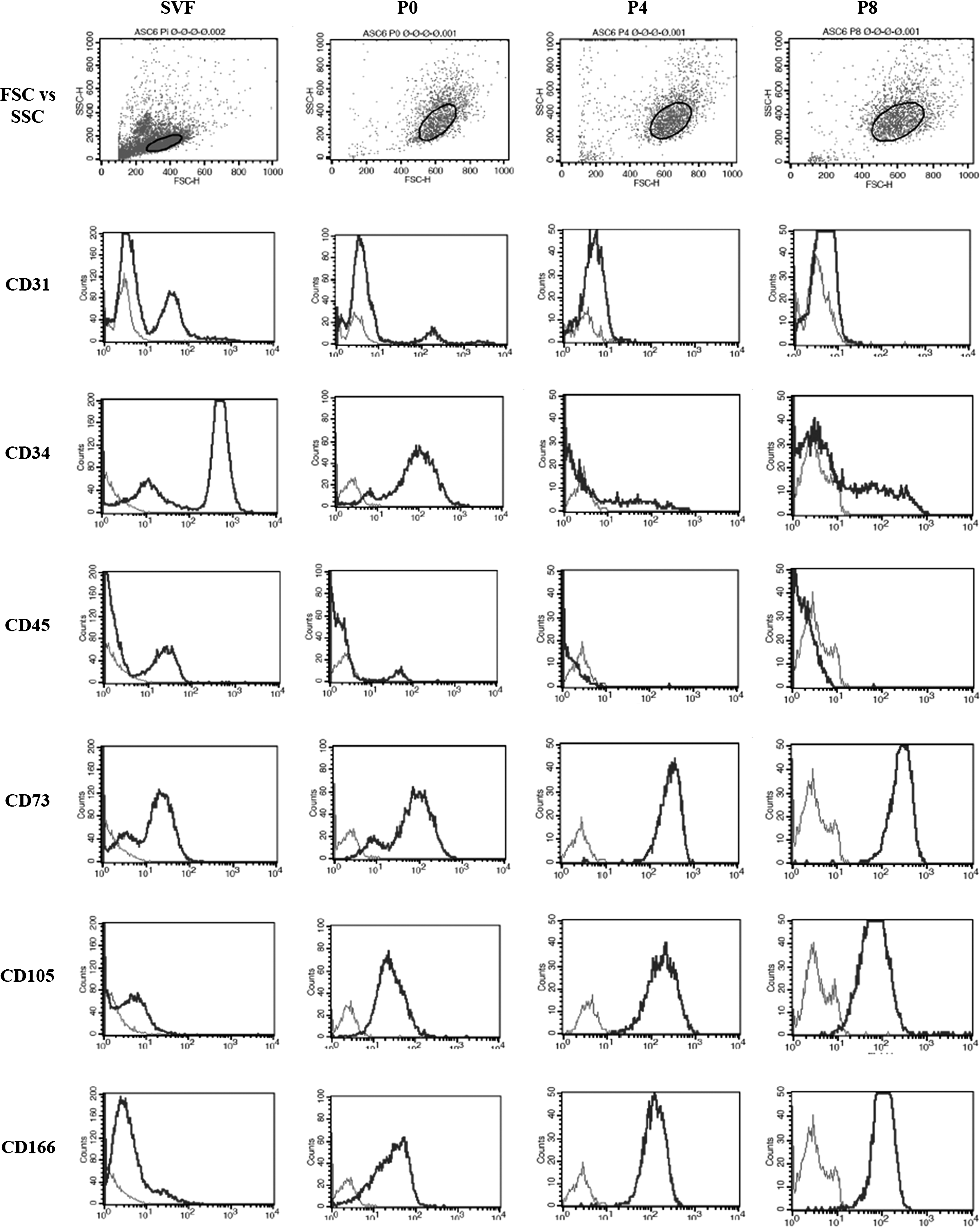
Changes in cell surface marker expression with serial passage of stromal vascular fraction (SVF) cultures. Representative histograms showing the change in cell surface markers, size (FSC), and granularity (SSC) with serial passage of a single adipose-derived stem cell (ASC) line (patient 6). Light gray, unstained control; dark gray, stained cells; P, passage; FSC, forward scatter; SSC, side scatter.

Differentiation of ASC and fibroblasts.
Cultured ASCs exhibited a morphology similar to fibroblasts (Fig. 2A). Furthermore, the surface phenotype of ASCs (passage 8) and fibroblasts was very similar (Table 1), with both cell types expressing adhesion molecules (CD29, CD45, CD49d, CD49e, CD51, CD54, and CD166) and other markers (CD13, CD59, CD73, CD90, and CD105), which in combination are typical of mesenchymal cells, but not specific endothelial (CD31, CD62e, and CD144) or leukocyte (CD45) markers. To confirm that these late passage ASC cultures were derived from CD34+ CD31− cells in the SVF, CD34+ CD31− cells were sorted from the other cell populations present in the SVF by flow cytometry (Fig. 2B). CD34+ CD31− cells sorted directly from SVF established adherent cell cultures with the same characteristics as late passage SVF cells, and accumulated lipid when exposed to AM (Fig. 2C), whereas the other cells sorted from SVF (the cells within the gate marked “non-ASC” in Fig. 2B) did not (data not shown).
The surface phenotype was determined by flow cytometry. Adipose-derived stem cell lines from five different donors were assessed.
In addition to expression by fibroblasts, CD90 is also expressed by endothelial cells, a subset of thymocytes and CD34+ bone marrow cells.
Therefore, all subsequent experiments were performed on cell lines derived from SVF from passage 2 onward as these cultures consisted of homogeneous populations of ASCs capable of differentiating into adipocytes.
Transduction of ASCs with rAAV vectors
ASCs from three donors were selected (subjects 1 (passage 3), 2, and 3; Table 2), and infected with 9 variants of rAAV at an MOI of 105 viral genomes per cell. After 3 days eGFP expression was analyzed by flow cytometry (Fig. 3 and Supplementary Fig. S3). Marked variation in ability to transduce ASCs was noted between variants. rAAV2 was the most efficient, transducing 61% of ASCs. Other serotypes that successfully transduced ASCs were rAAV5 (17%), rAAV6 (28%), and rAAV1 (3%). By comparison, 6% of cells were transfected using lipofectamine, consistent with previous studies. 10
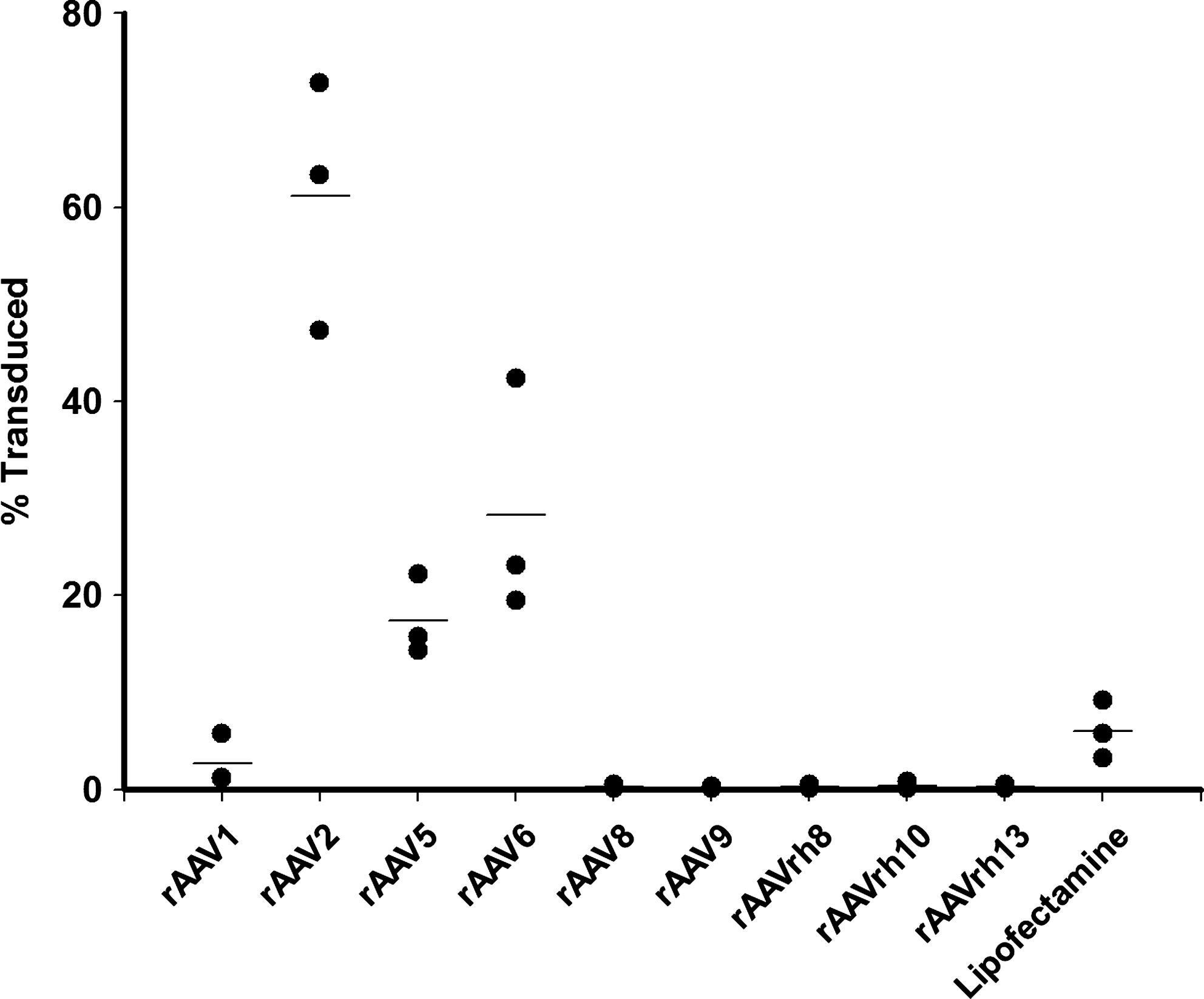
Percentage of viable ASC transduced by recombinant adeno-associated virus (rAAV) variants 3 days postinfection. ASC were infected at a multiplicity of infection (MOI) of 105 vg copies/cell. Transduction was assessed at 72 h postinfection by flow cytometric detection of eGFP expression. Comparison was made with transfection with lipofectamine. The experiment was performed in triplicate with ASC preparations from donors 1 (passage 3), 2, and 3 (see Table 2). The means of triplicate measurements are plotted. Lines represent the mean of different experiments. Variants were compared by a one-way ANOVA on log10-transformed data. Pairwise multiple comparisons were made by the Holm-Sidak method. rAAV2, rAAV5, rAAV6, and lipofectamine transduced a significantly higher percentage of cells than serotypes 8 and 9, and clones rh8, rh10, and rh13 (p<0.001); rAAV2 and rAAV6 transduced a significantly higher percentage of cells than rAAV1 (p<0.001). There was no statistically significant difference between rAAV2, rAAV5, rAAV6, and lipofectamine.
Comparison of rAAV2, rAAV5, and rAAV6
To further investigate transduction of ASCs by rAAV three ASC preparations from different donors (subjects 4–6, Table 2) were infected with rAAV2, rAAV5, or rAAV6 at an MOI of 103, 104, and 105. Cells were analyzed 72 h after infection. At an MOI of 103 transduction efficiency with rAAV2 was comparable to transfection with lipofectamine (Fig. 4A), whereas significantly less efficient transduction was seen with rAAV5 and rAAV6 (p<0.001). At MOIs of 104 and 105 rAAV2 transduced a significantly greater proportion of ASCs than lipofectamine (p<0.001). At an MOI of 105 there was no significant difference between the proportion of ASCs transduced by rAAV5, rAAV6, and lipofectamine (p>0.1).
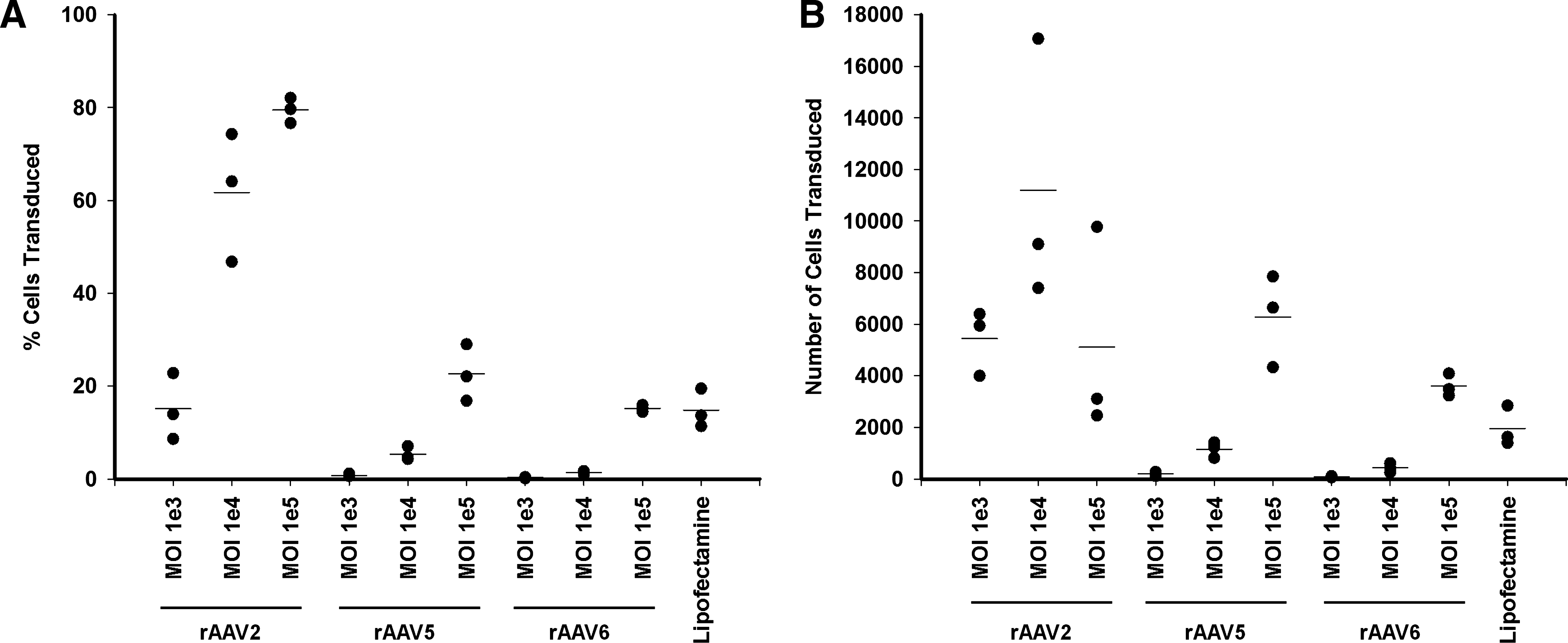
Infection of ASC at high MOI with rAAV2 but not rAAV5 or rAAV6 reduces cell viability. ASC from donors 4–6 (see Table 2) were infected with rAAV of serotypes 2, 5, or 6 at the indicated MOI. Transduction was assessed at 72 h by flow cytometric detection of eGFP expression.
While these results immediately suggested that rAAV2 would be the preferred serotype for transducing ASC, it was noted that the total cell count was reduced in rAAV2-infected samples. Indeed, the absolute number of ASCs transduced by rAAV2 was greater at an MOI of 104 than at an MOI of 105, although allowing for multiple comparisons the difference did not reach statistical significance (p=0.01) (Fig. 4B). Similarly, there was no statistical difference between the absolute number of ASCs transduced by rAAV2 at an MOI of 103 and 104 (p=0.05) or 103 and 105 (p=0.48), or rAAV5 or rAAV6 at an MOI of 105 (p=0.004). Both rAAV2 at an MOI of 104 and rAAV5 at an MOI of 105 transduced significantly more cells than lipofectamine (p<0.002). To confirm the reduction in the yield of viable cells after infection with rAAV2, we included Calibrite beads to control for any variation in cell recovery during sample preparation. A representative experiment is shown in Figure 4C. While infection with rAAV5 and rAAV6 had no effect on the number of viable cells, a decrease was noted after infection with rAAV2 at an MOI of 104 or 105. Of note, a 70% decrease in cell numbers was also seen with lipofectamine; a similar magnitude of decrease was only seen at the highest MOI with rAAV2. Hence, the apparent high transduction efficiency of rAAV2 at higher MOI was offset by a decreased viable cell yield. Therefore, of the protocols tested here, use of rAAV2 at an MOI of 104 yields the greatest number of transduced ASCs, although not significantly different from rAAV2 at an MOI of 103 or rAAV2, rAAV5, and rAAV6 at an MOI of 105.
Ability of transduced ASCs to differentiate
To establish that ASC differentiation capacity was not compromised after transduction with rAAV, 3 days postinfection with rAAV2, rAAV5, or rAAV6 eGFP+ ASCs were sorted by flow cytometry and cultured in AM to induce adipogenesis. Comparison was made with sorted cells cultured in CM and with unsorted cells cultured in CM or AM. After 14 days, cells were stained for incorporation of lipid with Oil Red O. All ASCs exposed to AM showed incorporation of lipid by Oil Red O staining (Fig. 5), whereas no adipogenesis was seen in ASCs cultured in CM or in dermal fibroblasts cultured in AM (data not shown).
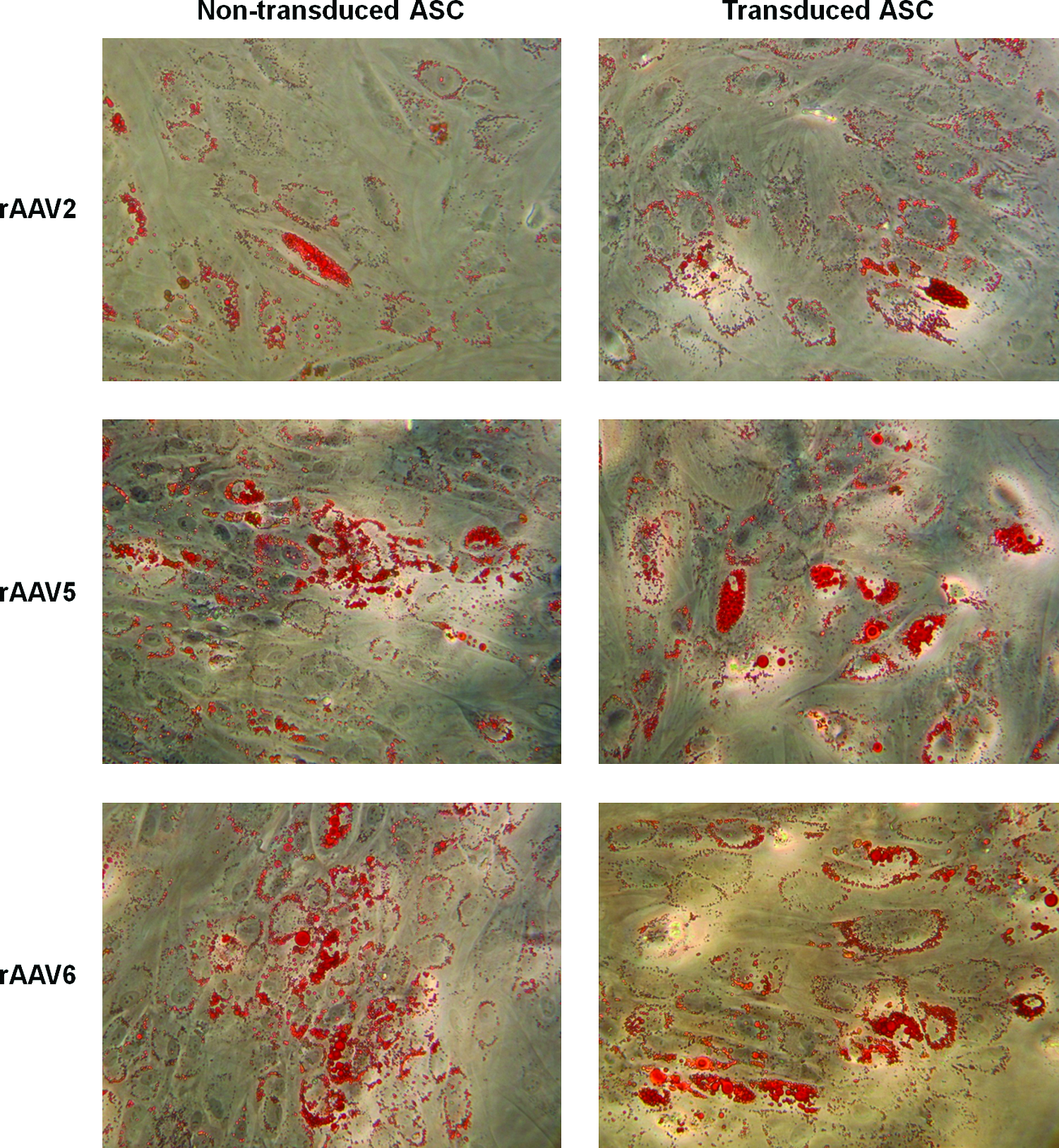
rAAV-transduced ASC are able to undergo adipogenic differentiation. ASC infected with rAAV2, rAAV5, and rAAV6 were sorted 72 h postinfection by fluorescence-activated cell sorting into transduced and nontransduced populations. Transduced and nontransduced ASC were cultured in AM for 14 days. Representative photographs of ASC stained with Oil Red O for intracellular lipid accumulation are shown (all 200×magnification). No staining was seen in wells not exposed to adipogenic medium. Color images available online at
Increased transduction with rAAV6 tyrosine capsid mutant Y731F
Increased transduction of murine and human bone marrow-derived MSCs by rAAV2 containing mutated surface-exposed tyrosine residues has recently been described. 38 Mutation of surface-exposed tyrosine residues blocks capsid phosphorylation by epidermal growth factor receptor protein tyrosine kinase (EGFR-PTK) and subsequent polyubiquitination and proteosomal degradation. 39 Given the cytotoxicity of rAAV2 in human ASCs we investigated the utility of rAAV6 tyrosine capsid mutants for increasing transduction efficiency. We have previously reported that with the exception of Y501 the AAV6 capsid contains analogous surface exposed tyrosine residues to AAV2, and that mutation of residues Y445 and Y731 increases transduction efficiency in HeLa cells. 31 We therefore assessed the effect of these mutations on the transduction of human ASCs (Fig. 6). ASCs from three donors (subjects 1 [passage 13], 7, and 8 in Table 2) were infected with rAAV6 (Y445F), rAAV6 (Y731F), the double mutant rAAV6 (Y445F + Y731F), or wild-type rAAV6 and the transduction efficiency assessed by flow cytometry. A modest increase in transduction efficiency was seen with Y731F (2.2-fold at an MOI of 104) but not with Y445F or with the double mutant Y445F + Y731F. Transduction of ASCs with capsid mutants had no adverse effect on viability (Supplementary Fig. S4).
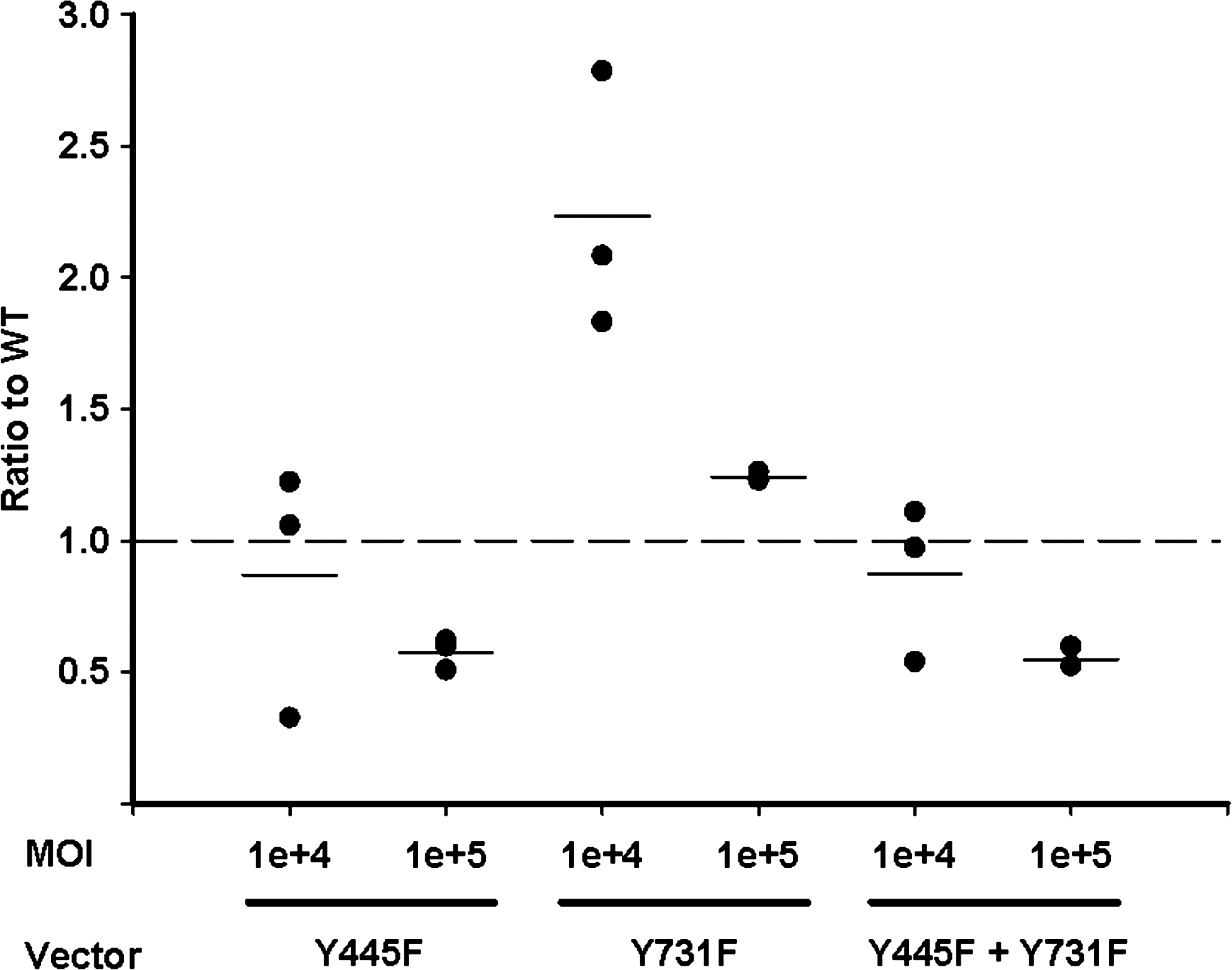
Transduction of ASCs by rAAV6 tyrosine capsid mutants. ASCs from donors 1 (passage 13), 7, and 8 (see Table 2) were infected at an MOI of 104–105 vg copies/cell with wild-type rAAV6, rAAV6 (Y445F), rAAV6 (Y731F), or the double mutant rAAV6 (Y445F + Y731F). Transduction was assessed 72 h postinfection by flow cytometric detection of eGFP expression. Transduction efficiency is plotted relative to wild-type rAAV6. The means of triplicate measurements are plotted; lines represent the mean of different experiments. Groups were compared by one-way ANOVA. Pairwise multiple comparisons were made by the Holm-Sidak method. rAAV6 (Y731F) at an MOI of 104 was significantly different to wild type (p<0.001); other differences were not significant. For viability, see Supplementary Figure S4.
Discussion
In this study we assessed the ability of various serotypes and clones of rAAV to transduce human ASCs. To ensure that experiments were conducted with a homogeneous population of ASCs, the phenotype of our ASC cultures was first extensively evaluated. Adherent cells isolated from SVF were found to be of mixed phenotype, including a population that corresponded to ASCs capable of differentiating into adipocytes (CD34+ CD31− cells), consistent with previous reports.35–37 The most prominent of the other populations was CD31+ CD45+, which probably represents contaminating monocytes, macrophages and granulocytes. Indeed CD31+ CD45+ CD14+ and CD31+ CD45+ CD15+ populations have previously been shown to comprise 3%–15% and 20%–40% of the SVF, respectively. 36 By passage 2 this population had disappeared, probably because it failed to survive, although down-regulation of CD31 and CD45 expression cannot be excluded. Similarly Yoshimura et al. demonstrated a loss of CD31+ and CD45+ expressing cells with culture. 36 Therefore, all transduction experiments were conducted on ASC cultures of passage 2 or later.
To confirm that cultured cells were indeed ASC, the surface phenotype of passage 8 ASC was assessed. Twelve cell surface markers were analyzed and the expression pattern of these markers on ASCs was consistent with previous reports26,35,36,40 and very similar to that of cultured dermal fibroblasts. We confirmed that both the late passage ASC cultures and CD34+ CD31− cells sorted directly from the SVF were able to differentiate into adipocytes, whereas the dermal fibroblast cell lines were not. It should be noted, however, that some dermal fibroblast cell lines are able to differentiate into different cell lineages, including adipocytes. 41
Having confirmed the homogeneity of our ASC cultures, we screened the ability of nine variants of rAAV to transduce ASCs. Our data reveal that rAAV serotypes 1, 2, 5, and 6 are capable of transducing ASCs. Notwithstanding the apparent cytotoxic effect of rAAV2 infection at higher MOI, rAAV2, rAAV5, and rAAV6 were the most efficient vectors for genetic engineering of ASCs. In contrast, serotypes 8 and 9 and clones rh8, rh10, and rh13 showed no ability to infect ASCs, even at an MOI of 105. The infectivity of all vector subtypes was confirmed in HeLa cells. 31
These data are broadly consistent with previous reports of rAAV-mediated transduction of MSCs derived from human bone marrow. Although several articles have described efficient transduction of bone marrow-derived MSC with rAAV of a single serotype (either unstated serotype19,20,24 or rAAV222,23) only two studies report a comparison of the transduction efficiency of different serotypes. Chng et al. assessed rAAV serotypes 1–6 and 8 and concluded that rAAV2 was the most efficient, whereas rAAV3 was within one log10. 18 Kim et al. assessed the same range of serotypes and demonstrated efficient transduction of MSC derived from both bone marrow and umbilical cord blood was restricted to rAAV2 and rAAV5. 21 Therefore, while our observation that rAAV2 and rAAV5 are the most efficient serotypes for the transduction of ASCs is consistent with previous studies, the finding that rAAV6 also efficiently transduces ASCs suggests subtle differences in the tropism of various vector serotypes for MSCs derived from different sources.
rAAV serotypes 1 to 5 have been shown to successfully transduce mouse adipose tissue in vivo, with rAAV1 exhibiting the greatest tropism. 15 However, it is not possible to assess the susceptibility of ASCs in that study because the adipose tissue was analyzed in totality and excipients such as Pluronic gels were used to improve transduction—indeed in some experiments, no transduction was observed without the excipients. 15
Overall, 70% of ASCs were transduced by rAAV2 at an MOI of 105, although due to serotype-specific cytotoxicity the absolute number of transduced cells was not statistically different from that seen with rAAV5 (20% transduced) and rAAV6 (22% transduced) at an MOI of 105 or rAAV2 at an MOI of 103 (15% transduced). The greatest number of transduced cells was seen with rAAV2 at an MOI of 104, with 62% of cells transduced with only moderate cytotoxicity, although this did not reach statistical significance. Significant inter-individual variation in transduction efficiency was seen with serotypes 2, 5, and 6 (see Figs. 3 and 4A). The efficiency of transduction with rAAV compared favorably with lipofectamine-mediated transfection, which transfected 10% of ASCs, consistent with published studies. 10 Moreover, marked cytotoxicity was seen with lipofectamine, with a 70% reduction in viable cell yield. This is in contrast to a recent report, which examined ASC viability at 72 h post-lipofectamine-transfection and found it to be reduced by only 20%. 42 The greater level of cytotoxicity seen in our study may be due to differences in seeding density or lipofectamine-to-DNA ratio. Reductions in viable cell yield were also observed when ASCs were infected with rAAV2 at an MOI of 105, with a smaller reduction at an MOI of 104. In contrast, rAAV5 and rAAV6 had no effect on cell viability at any MOI.
The increased cytotoxicity of rAAV2 in ASCs may be explained by its usage of αVβ5 integrin as a coreceptor. 43 αVβ5 integrin, which is expressed by ASCs, 44 has been shown to be important in the attachment, self-renewal, and proliferation of human embryonic stem cells in vitro. 45 In other cell lines blockade of αVβ5 integrin has been associated with apoptosis or suppression of proliferation. 46 Different coreceptors are used by rAAV5 and rAAV6.47,48
Mutation of surface-exposed tyrosine residue Y731 but not Y445 in the AAV6 capsid led to an increase in transduction of ASCs. This effect was most evident at the lower MOI of 104, with a 2.2-fold increase in the number of ASC transduced compared with wild type. Homologous mutations (Y444 and Y730) in the AAV2 capsid have recently been shown to increase the transduction of murine and human bone marrow-derived MSCs by 10- and 3-fold and 6- and 5-fold, respectively; double mutants increased transduction by 31-fold and 8-fold, respectively. 38 In contrast, we found the double mutant to be no more efficient than wild type at transducing human ASCs. This likely reflects a difference in the biology of the AAV2 and AAV6 capsids. Indeed, we have previously found the Y445F mutation to be deleterious to the transduction of human monocyte-derived dendritic cells but not HeLa cells. 31 This may reflect a difference in receptor usage, postuptake trafficking, or uncoating in ASCs/MoDCs and HeLa cells. Nonetheless, the increase in transduction seen with rAAV6 (Y731F) suggests that phosphorylation of surface-exposed tyrosine residues by EGFR-PTK is a rate-limiting step to transduction of human ASCs by rAAV. Therefore, similar improvements in transduction may be expected with AAV2 tyrosine capsid mutants, which may allow vector dose to be reduced with a concomitant reduction in cytotoxicity.
The utility of rAAV as a vector for gene transfer to ASCs may be limited by its packaging capacity. The wild-type genome is 4.7 kb, whereas genomes up to 5.2 kbp can be packaged without loss of transduction efficiency.49,50 Larger genes can be transferred using two vectors and trans-splicing. 51 iPS cells can be generated from human fibroblasts by enforced expression of Oct3/4 and Sox2 with either Klf4 and c-Myc5,52 or Nanog and Lin28. 53 In ASCs, delivery of Oct3/4, Sox2, Nanog, and Lin28 using “minicircle” DNA induced pluripotency. 6 Delivery of more than two transcription factors by rAAV would most likely require two or more vectors. However, it is possible that basal ASC expression of some of the iPS genes might allow induction of pluripotency with fewer than four genes. Indeed, iPS cells have recently been generated from ASCs with Oct3/4, Sox2, and Kfl4 alone. 54 Given the highly efficient transduction of ASCs by rAAV, improvements in reprogramming efficiency may be achievable without the use of retroviral vectors and the associated risk of integration.8,9 While there are also concerns over the potential of rAAV to cause insertional mutagenesis,55,56 these are tempered by the lack of tumorigenicity in mice 57 and of pathological association with wild-type infection in humans. 58
Pre-existing immunity to wild-type AAV is high in the community and may limit in vivo gene transfer. Neutralizing antibodies to AAV2, AAV5, and AAV6 have been found in 30%, 13.5%, and 30%, respectively, of healthy adults. 59 Neutralizing antibodies impair infection with rAAV vectors in vivo. 60 Pre-existing cytotoxic T cells specific for the AAV capsid may also eliminate cells transduced in vivo.61,62 As manipulation of ASCs is likely to be performed ex vivo antibody-mediated neutralization of the vector is unlikely to be a problem. However, the elimination of vector-transduced ASCs upon implantation by pre-existing AAV capsid-specific cytotoxic T cells cannot be excluded.
In conclusion, using a well-characterized, homogeneous population of human ASCs, we have identified rAAV serotypes 2, 5, and 6 as efficient vectors for gene transfer. While rAAV2 adversely affected ASC viability at higher MOI, no such effect was seen with serotypes 5 and 6. Furthermore, a modest increase in transduction efficiency was achieved with the AAV6 tyrosine capsid mutant Y731F, suggesting that capsid degradation is a rate-limiting step in transduction of ASCs. Therefore, rAAV5, rAAV6 (in particular the Y731F capsid mutant), or at lower MOIs, rAAV2, should be the vectors of choice for genetic manipulation of human ASCs.
Footnotes
Acknowledgments
We thank Stephen Edgar for expert assistance with cell sorting. We gratefully acknowledge funding from the Stevenson Trust, the William and Lois Manchester Trust, the Royal Australasian College of Surgeons, and the Maurice Wilkins Centre. This research was conducted during the tenure of a Clinical Research Training Fellowship from the Health Research Council of New Zealand (J.U.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.