
Abstract
Regenerative therapies are promising treatments for early intervertebral disc degeneration. To test their efficacy, an in vitro tissue-level model would be valuable. Nucleus pulposus (NP) explant culture may constitute such a model, as the earliest signs of degeneration are in the NP. However, in NP explant cultures, balancing tissue osmolarity is crucial to preventing swelling, proteoglycan (PG) loss and, therefore, maintaining a native cell environment. In this study, we investigated the effect of medium osmolarity on NP explants. We hypothesized that balancing the inherent tissue osmolarity would prevent swelling and thus maintain NP tissue in a native state. Bovine NP explants were cultured for 21 days in hypo-, iso-, and hyper-tonic conditions using either sucrose or polyethylene glycol (PEG) to raise medium osmolarity. Explants were analyzed for water and biochemical content, cell viability, gene expression, and tissue histology, and compared to day 0 samples. In hypo-tonic and both sucrose cultures, swelling was not prevented, resulting in PG loss and changes in cell behavior. Only PEG cultures maintained water and biochemical content and a histological aspect similar to those of native tissue, with better results for hyper- than for iso-tonic conditions. Using PEG to raise culture medium osmolarity, we were able to maintain the NP tissue specific matrix composition, important for disc cell behavior. This approach, thus, constitutes a promising model to test regenerative therapies for early intervertebral disc degeneration.
Introduction
The intervertebral disc is the soft cartilage-like tissue that is located in between the vertebral bodies, preserving motion and dampening peak loads in the back. The intervertebral disc consists of a gelatinous core, the nucleus pulposus (NP), and a fibrous outer ring, the annulus fibrosus. The NP main constituents are collagen type-II, proteoglycans (PGs), and water. 5 The PGs are highly negatively charged glycosylated proteins that attract water. This gives rise to an increased swelling pressure inside the NP, which is constrained by the annulus fibrosus. When the disc degenerates, there is a shift from an anabolic to a catabolic environment. The NP cells decrease their production of the main matrix proteins and increase their production of degrading enzymes.4,6,7 As a result, the NP loses PGs and water, 8 initiating a progressive degenerative cascade resulting in tissue fibrosis and loss of integrity, which is manifested as decreased disc height and loss of mechanical function that may ultimately lead to low back pain.2,9
Regenerative therapies aim to treat disc degeneration at an early stage to prevent low back pain from occurring.10,11 These therapies have been shown to be promising in animal models,12–14 as they could delay degeneration,13,14 but still need to be further developed as they cannot fully restore the disc to its original healthy state.13–15 To screen for possible therapies, experimental models of the disc are needed.3,15 Although animal models are the most sophisticated preclinical system for therapy development, disc degeneration in such models are mostly artificially induced and their similarity to the human disease remains questionable. 16 Further, they are low-volume throughput, cost-intensive, and have ethical considerations.16,17 Hence, the use of in vitro models is appealing. The whole-disc explant culture system preserves the native environment best, but the throughput volume also remains low and culture duration may not be sufficient to detect changes of the tissue matrix. 18 Although cell culture is less costly and may be accomplished in high-throughput systems, the effect of removing the cells from their native tissue environment on the observed therapeutic response is unknown. Thus, explant culture of NP tissue may constitute a suitable in vitro model as the earliest detectable changes in degeneration take place in the NP. 19 A key benefit of an explant system over cell culture is that the efficacy of cell and growth factor-based therapies can be tested in an environment that is very close to the in vivo environment. 20 This environment includes hypoxia, low glucose, low pH, and high osmolarity. 21 In such a system high osmolarity is particularly difficult to maintain, as swelling pressure is not balanced by the annulus fibrosus anymore. As a result the NP tissue swells and subsequently PGs leach out, reducing the tissue osmolarity. 21 Balancing the high osmolarity is thus critical in maintaining the in vivo environment in cultures of NP explants.
In previous studies, up to 1 week culture of NP cells with raised osmolarity via sodium chloride (NaCl) showed positive effects on cell viability, 22 PG production,22,23 and gene expression of matrix proteins 24 when compared to culture in normal media. A study on rat discs showed that the cell viability was maintained for 1 week, when osmolarity was balanced. 25 When NP explants were cultured up to 5 h in balanced osmolarity, by NaCl, sucrose, 26 or polyethylene glycol (PEG),26,27 the PG production was increased compared to cultured in normal medium. Although these models are promising to test regenerative therapies, NP tissue should be cultured in vitro for weeks, not hours, to be able to see a significant therapeutic effect.
Hence, the aim of this study was to culture NP explants for 3 weeks in a swelling balanced environment. To do so, we cultured bovine NPs in different medium osmolarities and investigated if their biochemical content, cell viability, and gene expression could be maintained. From possible osmotic agents, we selected sucrose 26 and PEG, 27 because NaCl might alter the cellular ionic balance. 28 Even though sucrose might not be able to fully prevent tissue swelling, 26 we were still interested in using sucrose for longer culture durations to investigate the beneficial effects of a balanced cell osmolarity even in a swollen tissue environment. We hypothesized that by balancing the osmolarity of NP explants, swelling and PG loss can be prevented and tissue matrix content can be maintained.
Materials and Methods
Culture
To test our approach bovine coccygeal NP tissue, which resembles human tissue very closely in size, content, and osmolarity, 29 was used. NP explants were harvested under sterile conditions from fresh caudal discs of 24-month-old cows. These were obtained from the abattoir according to local regulations and a total of 60 discs (CC1 to CC4) were harvested from 15 donors. The different levels were distributed equally and randomly among the different experimental conditions. The discs were opened transversally, directly underneath the endplate, and NP explants were punched with an 8-mm-diameter biopsy punch (Kruuse) from the center of the NP. The samples were immediately placed in culture.
From literature,
26
bovine in situ NP tissue osmolarity was expected to be 430 mOsm/kg H2O. Five culture groups were then defined in three osmotic conditions:
• Hypo-tonic to native in situ NP (323 mOsm/kg H2O): standard medium • Iso-tonic to native in situ NP (430 mOsm/kg H2O): standard medium+107 mM sucrose (Merck) or 8.2% w/v PEG (20 kDa; Sigma) • Hyper-tonic to native in situ NP (570 mOsm/kg H2O): standard medium+247 mM sucrose or 13.3% w/v PEG
Standard medium was prepared from high glucose (4.5 g/L) Dulbecco's modified Eagle's medium powder (Gibco, Invitrogen) in milliQ filtered water, supplemented with 1% penicillin/streptomycin (Lonza), 3.7 g/L sodium bicarbonate (Sigma), 100 mg/L ascorbic acid (Sigma), and 10% fetal bovine serum (Gibco). All media were filter sterilized and the pH was adjusted to 7.1, pH of a healthy human disc. 30 The osmolarity of all media was checked with a freezing point osmometer (Osmomat 030; Gonotec). In hypo-tonic and sucrose cultures, NP explants were directly placed in the medium (Fig. 1A). In PEG cultures, NP explants were placed inside dialysis tubing (15 kDa molecular weight cut-off [MWCO]; Spectra-Por) and closed with clips (Spectra-Por) to prevent PEG (20 kDa), which might be cytotoxic, 27 from entering the tissue (Fig. 1B). Samples were cultured for 21 days in an incubator at 37°C, 21% O2, and 5% CO2 and media were changed three times a week.

Culture systems (for hypo-, iso-, and hyper-tonic conditions).
For every culture condition, 10 samples (corresponding to 10 donors) were cultured and cut in half at the end of the culture period. From these 20 half samples, 5 were used for each of the four different analyses: biochemistry, gene expression, histology, and viability. As a control, 10 day 0 samples (fresh native tissue) were analyzed immediately after harvesting in a similar manner (n=5 discs each from different donors).
Biochemical and water content
At day 0 and 21, the samples were blotted dry, weighed, and subsequently stored frozen at −30°C. The samples were then placed in a lyophilizer (Freezone 2.5; Labconco) overnight and the dry weight was measured. The water content was calculated from the difference in wet and dry weight, divided by the wet weight. The dried samples were then digested in papain solution (100 mM phosphate buffer, 5 mM L-cystein, 5 mM ethylene diamine tetraacetic acid, and 125–140 μg/mL papain, all from Sigma) overnight at 60°C. The digested samples were then used to determine their content of: sulfated glycosaminoglycans (sGAG), as a measure of PGs, hydroxyproline (HYP), as a measure for collagen content, and DNA. sGAG content was determined with the Dimethyl Methylene Blue (DMMB) assay 31 using shark cartilage chondroitin sulfate as reference (Sigma). The fixed charge density (FCD) was determined from the GAG content per wet weight, similar to the method used by Narmoneva et al. 32 HYP content was measured using the Chloramin-T assay 33 and a trans-4-hydroxyproline (Sigma) reference. DNA content was measured using the Hoechst dye method, 34 with a calf thymus DNA reference (Sigma). The amounts of sGAG, HYP and DNA were expressed per mg dry weight of tissue.
Gene expression
Samples were snap-frozen in liquid N2 and stored at −80°C until RNA isolation. A 316 SS 8 mm bead and custom-made lid were placed in a 2 mL Eppendorf tube and snap-frozen in liquid N2. Frozen samples were placed in between bead and lid, and disrupted with a micro dismembrator (Sartorius) for 20 s at 1500 rpm. This was repeated if necessary with the sample snap frozen between each cycle. After disruption, RNA was isolated using TRIzol (Invitrogen) and purified using the Qiagen mini-kit (Qiagen). The quantity and purity of isolated RNA was measured with a spectrophotometer (ND-1000; Isogen). Absence of genomic DNA contamination in the isolated RNA was checked using end-point PCR (C-1000; Biorad) with genomic DNA primers for GAPDH and gel-electrophoresis. The VILO-kit (Invitrogen) was used to reverse-transcribe 90 μg total RNA. The genes of interest and corresponding primer pairs are summarized in Table 1. Primers were designed with Beacon designer software (Premier Biosoft) and ordered from Sigma. 18S (PrimerDesign) was selected as reference gene from three genes (RPL13A, GAPDH and 18S) as the most stable gene throughout our experimental conditions. Gene expression was investigated with real-time PCR (iCycler; Biorad), and expression difference (Δ Ct) was calculated as the difference between the gene of interest and reference gene 18S. Levels of expression are expressed as 2−ΔCt. When gene expression was not detected, the 2−ΔCt value was set to 0 to conduct the statistical analysis.
MMP, matrix metalloproteinase; TIMP, tissue inhibitors of metalloproteinase.
Histology
Samples were fixed with 3.7% phosphate buffered formalin for 24 h at room temperature and then dehydrated and embedded in paraffin using a tissue-processor (STP-120; Microm). Subsequently 5-μm-thick slices were cut with a microtome (Leica) and stained with Weigerts Hematoxylin for cell nuclei, Safranin-O for PGs, and Fast Green for collagen. 35
Cell viability
Directly after harvesting, samples were placed in 10 μM Calcein AM (Sigma) and 10 μM Propidium Iodide (Invitrogen) in phosphate-buffered saline. These were first incubated for 1 h at 4°C to allow for dye diffusion into the tissue, followed by 1 h at 37°C to allow hydrolysis of Calcein AM. With a confocal microscope (LSM 510 META; Zeiss), the cell viability was assessed at a depth of 100 μm from the cutting edge.
Statistics
Statgraphics (Manugistics, Inc.) was used for statistical analysis. For the biochemical data, one-way analysis of variance was performed for all the groups, followed by Bonferonni corrected post-hoc t-tests. For the gene expression data, Kruskal-Wallis test was performed on the 2−ΔCT data, followed by Bonferonni corrected post-hoc Mann-Whitney U tests. Statistical significance in all cases was assumed for p<0.05.
Results
Biochemical and water content
After 21 days in hypo-tonic medium, the water content of NP tissue (93%) increased considerably compared to day 0 (82%, Fig. 2A). Similarly, DNA and HYP contents (0.12% and 6% compared to 0.04% and 3%, respectively) were also significantly increased (Fig. 2B, C). Most of the sGAG content, however, was lost (10% compared to 52%, Fig. 2D); therefore, the FCD decreased substantially (0.03 compared to 0.39 mEq/kg, Fig. 2E).

Effects of osmolarity on nucleus pulposus (NP) tissue biochemical composition after 21 days.
The water content in NP tissue increased significantly for both iso- and hyper-tonic sucrose conditions (90% and 85%, respectively, Fig. 2A). DNA content decreased significantly only in hyper-tonic sucrose (0.02%, Fig. 2B). HYP content was similar to day 0 in both conditions (Fig. 2C). Most strikingly, the sGAG content decreased (4% and 3%, respectively, Fig. 2D), even more than in hypo-tonic medium resulting in substantially decreased FCD (0.02 mEq/g for both conditions, Fig. 2E).
The hydration of NP tissue increased only in iso-tonic PEG (86%, Fig. 2A), but DNA and HYP contents stayed similar to day 0 in both conditions (Fig. 2B, C). The sGAG content and FCD decreased some in iso-tonic PEG (43% and 0.24 mEq/g, respectively, Fig. 2D, E), but much less than in the hypo-tonic and sucrose media. In fact for hyper-tonic PEG, all the biochemical content, including sGAG and FCD, were similar to native tissue (day 0).
Histology
The Safranin-O/Fast Green staining showed a severe loss of PGs in hypo-tonic medium (Fig. 3B) and both sucrose groups (Fig. 3C, D) compared to day 0 (Fig. 3A). Moreover, there was a fibrous layer visible at the edge of the NPs cultured in hypo-tonic conditions (data not shown). Although PGs were lost from the extracellular matrix, most of the cells showed a PG-rich pericellular matrix (Fig. 3B–D). The loss of PGs from the extracellular matrix was partially preserved in iso-tonic PEG (Fig. 3E) and resembled native tissue best in hyper-tonic PEG (Fig. 3F).

Safranin-O/Fast Green stained sections of NP tissue after 21 days. Day 0
Cell viability
In day 0 tissue, chondrocyte-like cells were visible, and at a depth of at least 100 μm, there was high cell viability (Fig. 4A). In all groups, this cell viability was preserved during culture (Fig. 4B–F). However, in hypo-tonic medium, the cell density was much higher and cell morphologies similar to a fibroblast phenotype were visible (Fig. 4B). In sucrose cultures, these morphology changes were also observed, but to a lesser extent (Fig. 4C, D). Cell density and morphology were similar to day 0 in both PEG cultures (Fig. 4E, F).
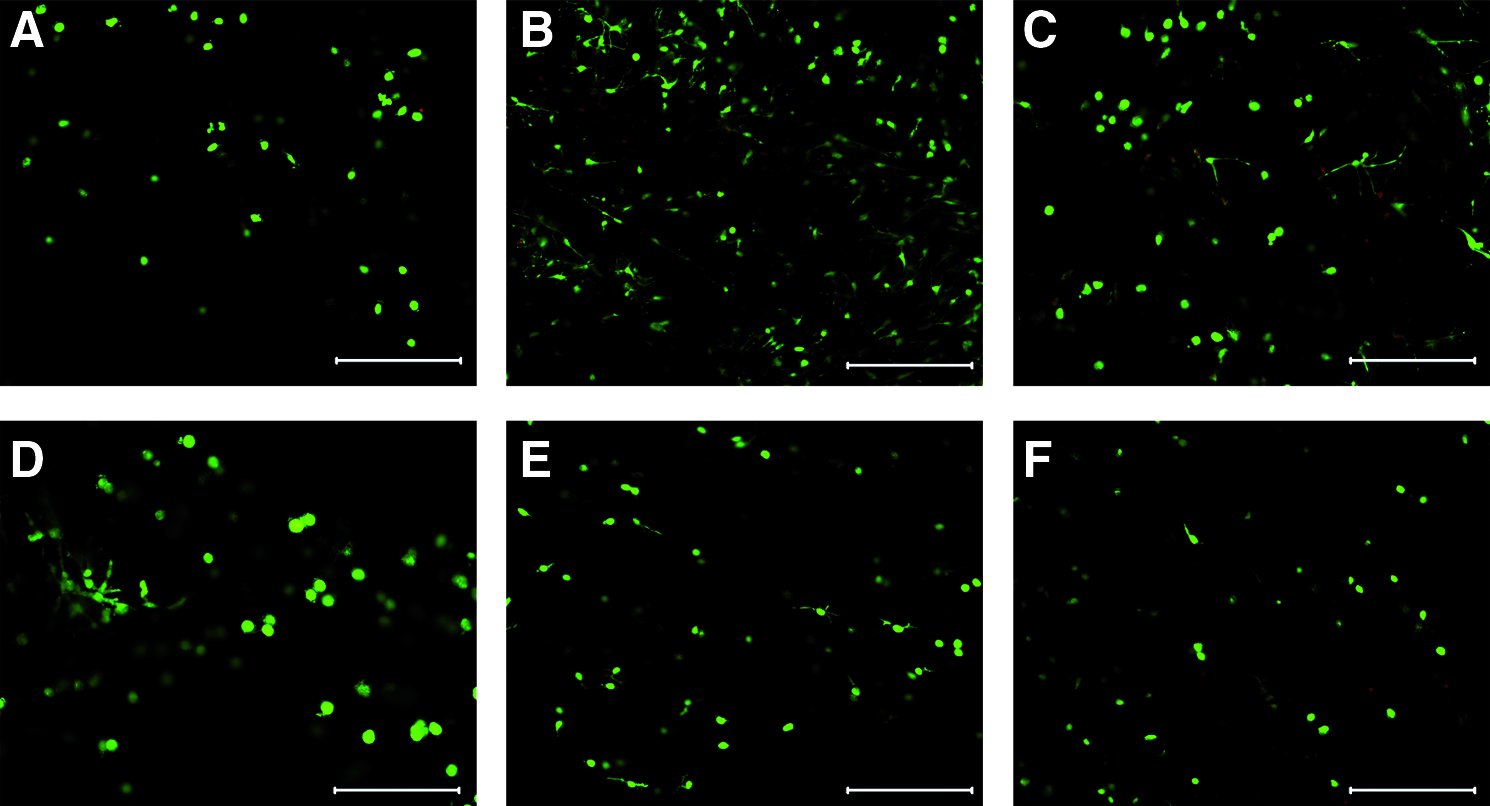
Calcein AM/propidium iodide staining of NP tissues after 21 days. Day 0
Gene expression
Aggrecan mRNA expression was downregulated in hypo-tonic medium and both PEG groups compared to day 0. Collagen type II expression was also downregulated in both PEG groups compared to day 0. Collagen type I expression was similar to day 0 in all groups, but in iso-tonic PEG its expression was significantly lower than in iso-tonic sucrose (Fig. 5). No significant differences were observed for tissue inhibitors of metalloproteinase 1 (TIMP 1) expression (Fig. 6A) for all groups. Expression of TIMP 2 was higher in both PEG groups compared to hypo-tonic medium (Fig. 6B); however it was not upregulated compared to day 0. Matrix metalloproteinase 13 (MMP-13) was not expressed in day 0 tissue, but was upregulated in iso-tonic sucrose and PEG (Fig. 6).
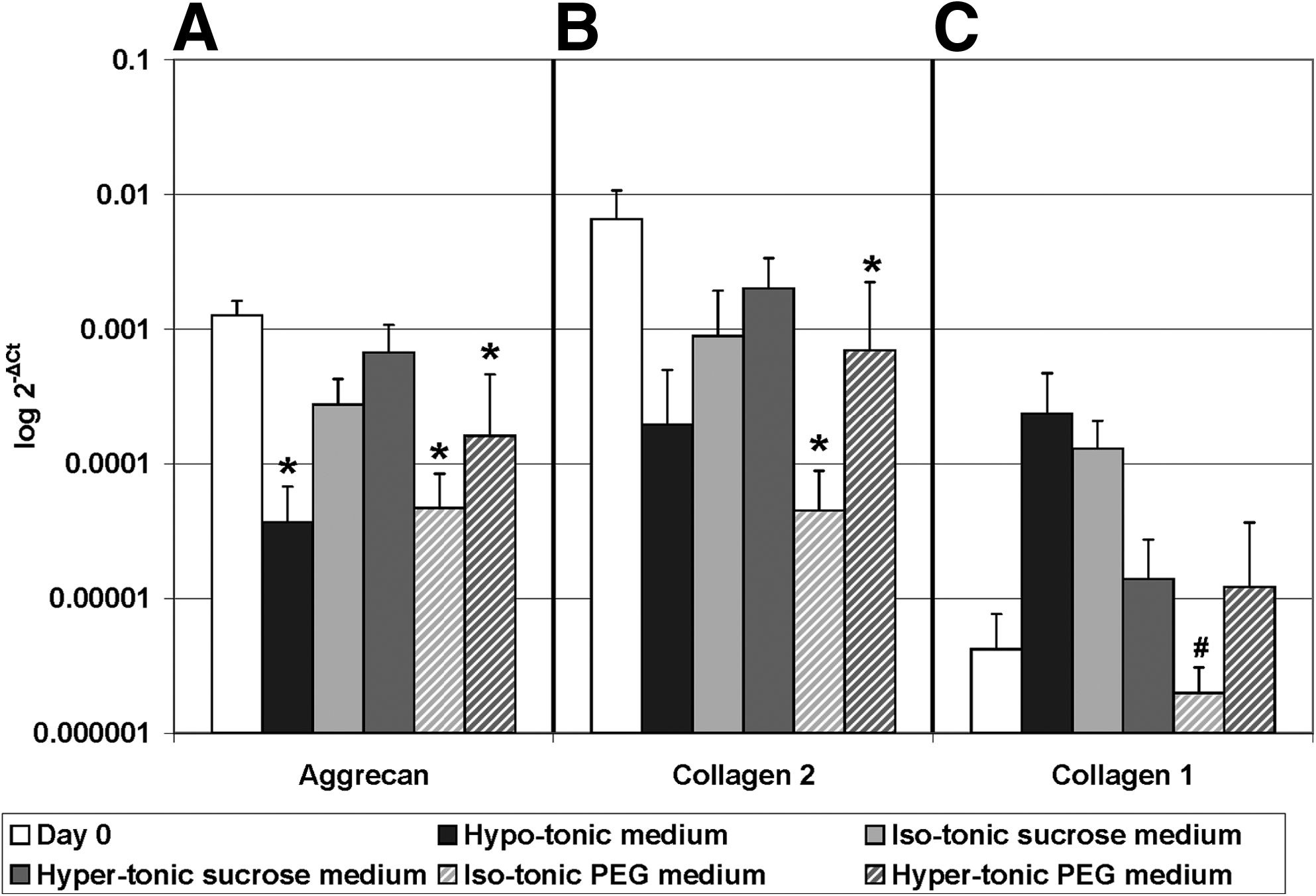
Gene expression of matrix proteins: Aggrecan, Collagen type II and type I. Gene expression was calculated relative to reference gene 18S. Values are mean±standard deviation, n=5. *Different from day 0; p<0.05; #different from sucrose 430; p<0.05 (please note the logarithmic Y-axis and that error bars are also logarithmic).
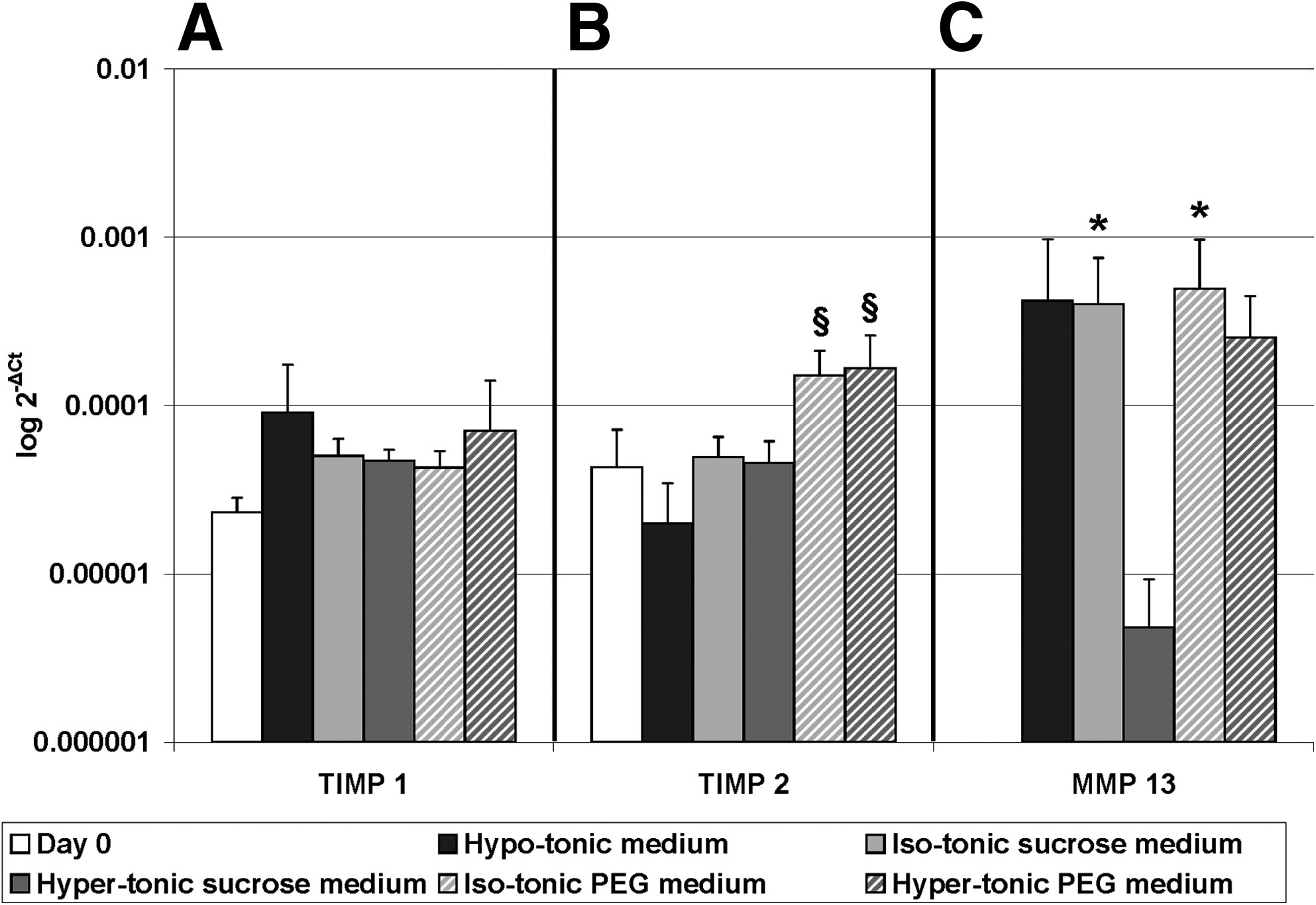
Gene expression of tissue inhibitors of metalloproteinase 1 (TIMP 1) and 2 and matrix metalloproteinase (MMP-13). Gene expression was calculated relative to reference gene 18S. Values are mean±standard deviation, n=5. *Different from day 0, p<0.05, §different from hypo-tonic medium, p<0.05 (please note the logarithmic Y-axis and that error bars are also logarithmic).
Discussion
Our results indicate that balancing tissue osmolarity is crucial in in vitro NP explant cultures. When NP tissue is cultured in hypo-tonic culture medium (323 mOsm/kg) for 21 days, swelling occurred due to the difference in osmotic pressure between tissue and medium. This swelling may lead to disruption of the collagen matrix and subsequent PG loss 21 (Figs. 2D and 3B). With this change in tissue osmolarity (decreased FCD), the cells dedifferentiated into a fibroblast-like phenotype (Fig. 4B) and a fibrous layer formed at the edge of the sample. This dedifferentiation is confirmed by cell morphology, decrease in aggrecan mRNA expression and increase in collagen type-I expression (Fig. 5).
In the present study, either sucrose condition was not able to maintain PG content for 21 days (Fig. 2), as expected from short-term studies where sucrose was able to maintain PG production but not to impede PG loss. 26 Because no semipermeable membrane was used to exclude sucrose from the tissue, we suspected the sucrose to enter the tissue; therefore, its content in the cultured NP was measured with a biochemical assay. In both groups, sucrose content increased compared to day 0, with higher content in hyper- (67%) than in iso-tonic (28%) groups. However, the cell response in these conditions was not as strong as with hypo-tonic medium. Thus, although the samples still swelled and lost most of their PGs, the sucrose was able to create higher tonicity at the cell level, and in doing so to maintain cell phenotype. This is supported by the gene expression data showing that the cells retain their phenotype (Fig. 5) and the Safranin-O/Fast green staining, which showed a GAG-rich pericellular matrix around the cells (Fig. 3C, D). This confirms previous short-term studies22–24,26 which showed that sucrose can be used as an osmotic agent to balance osmolarity in systems using isolated NP cells (2D culture or 3D seeded scaffolds).
Only culture media supplemented with PEG could prevent tissue swelling and PG loss from NP tissue. In particular with hyper-tonic PEG (570 mOsm/kg), the tissue biochemistry stayed similar to the fresh tissue, even after 21 days in culture (Fig. 2). However, there was an unexpected decrease in matrix protein gene expression (Fig. 5) for both PEG groups compared to day 0. And for isotonic PEG only, there was an increase of MMP-13 mRNA expression (Fig. 6). Although the biochemistry and histology results did not show any signs of degradation (Figs. 2 and 4), the cells might be less active than in native NP. Moreover, as turn-over might be low, we cannot exclude that there will be degradation in longer culture durations. A possible cause for this difference might be the semipermeable membrane used in both PEG conditions. The membrane prevents PEG from entering the tissue, but also impedes certain growth factors to enter and certain catabolic factors to leave, which might result in an unfavourable environment. This can be investigated by using a higher MWCO membrane and PEG with a greater average molecular weight. Another possibility is that the other factors in the in vivo environment (hypoxia, dynamic loading and low glucose) are important to maintain the cell phenotype. These factors are already being implemented in the model in our lab and we are also culturing for longer periods to exclude that degradation is occurring.
Interestingly, hyper-tonic PEG performed better than iso-tonic PEG, indicating that the osmolarity of our bovine NP samples may have been higher than 430 mOsm/kg H2O, a value derived from literature.
26
If we calculate the osmolarity from the FCD of day 0 tissue (Fig. 2C), using
36
:
with Cex the external salt concentration, then native tissue should have an osmolarity of 496 mOsm/kg. So the osmolarity of our bovine NP tissue was indeed higher than that expected from literature, 26 but not as high as hyper-tonic PEG (570 mOsm/kg). Alternatively, we can use this equation to calculate the final osmolarity of the tissue after 21 days in culture, from the FCD of both PEG conditions (Fig. 2C). This results in 387 mOsm/kg for iso-tonic PEG (aimed to be 430 mOsm/kg) and 457 mOsm/kg for hyper-tonic (aimed to be 570 mOsm/kg). Thus, there is still a discrepancy between the tissue osmolarity and the measured osmolarity of our PEG solutions. A possible explanation may come from our measurements of the osmolarity of PEG solutions with a freezing point osmometer. According to Schiller et al., 37 the osmolarity of PEG solutions is overestimated when measured with this device and suggests that a vapor pressure osmometer is more accurate in determining the osmolarity of PEG solutions. The extent of this effect needs to be investigated for our PEG culture media.
To use this approach to screen regenerative therapies, ideally human degenerated tissue needs to be used instead of bovine. Using data from literature on water and GAG content from different grades of the human degenerated NP, 38 the PEG concentration can be adjusted to balance the osmolarity of these tissues in culture. This approach is actually being implemented in our group.
Only a few other studies have reported on NP explant cultures. Feng et al. cultured rabbit NP explants for 14 days in free swelling conditions. 39 During culture the gelatinous aspect of the NP tissue was gradually lost and the number of apoptotic cells increased, showing the importance of counteracting NP swelling. Le Maitre et al. cultured human NP explants inside a plastic ring for 21 days, thus constraining the explants from the sides, but not the top and bottom. 40 Cultured samples preserved macroscopic matrix integrity as confirmed by histology. However, as the samples were not fully constrained, there is still the possibility that the samples swelled to some extent and a more quantitative assessment (biochemical assays) might show that PG content changed. In our system, the swelling pressure is balanced by elevated media osmolarity, assuring a homogenous constrain to NP swelling and it is possible to add further dynamic hydraulic pressure. 41
Conclusion
These results show that balancing the osmolarity of NP tissue is crucial to prevent swelling and GAG loss, which lead to phenotypical changes. 13.3% w/v PEG dissolved in culture medium was able to keep fresh bovine NP tissue stable for 21 days in culture on the tissue level and therefore constitute a promising first step toward a physiological NP explant model to test regenerative therapies for intervertebral disc degeneration. This system is currently implemented in our lab for a longer culture period and under physiological conditions.
Footnotes
Acknowledgment
This research forms part of the Project P2.01 IDiDAS of the research program of the BioMedical Materials institute, co-funded by the Dutch Ministry of Economic Affairs, Agriculture and Innovation.
Disclosure Statement
The authors have nothing to disclose concerning the content of this manuscript.