
Abstract
In this study, structure and biomechanical properties of freeze-dried decellularized porcine pulmonary heart valves were investigated. Heart valves were dissected from porcine hearts. The tissues were decellularized and separated in three groups: (1) without lyoprotectant, (2) with 5% sucrose, and (3) with a mixture of 2.5% sucrose and 2.5% hydroxyl ethylene starch (HES), and then underwent freeze-drying. Freeze-drying in the absence of lyoprotectants caused an overall more disintegrated appearance of the histological architecture of the porcine valves, especially between the fibrosa and the ventricularis layers. Freeze-dried tissues with lyoprotectants have a looser network of collagen and elastic fibers with bigger pore sizes. Tissue freeze-dried in the absence of lyoprotecants had the largest pore sizes, whereas the tissue freeze-dried in the presence of protectants showed pores of intermediate sizes between the decellularized tissue and the unprotected freeze-dried samples. Tissue freeze-dried with sucrose alone displayed less porosity than tissue freeze-dried with the sucrose/HES mixture, whereas no significant differences in biomechanical properties were observed. Decellularization decreased the elastic modulus of artery tissue. The elastic modulus of freeze-dried tissue without protectants resembled that of decellularized tissue. The elastic modulus values of freeze-dried tissue stabilized by lyoprotectants were greater than those of decellularized tissue, but similar to those of native tissue.
Introduction
Biological scaffolds can be stored at reduced supra zero temperatures (i.e., in standard refrigerators) to avoid degradation.8,9 Alternatively, the scaffolds can be stored in frozen or freeze-dried states.10,11 Freeze-drying has the advantage that it allows storage of the tissue completely dried, making banking and transport easier. Freeze-drying is a technique where water is removed from frozen material under reduced pressure, first by sublimation of the freezable water fraction (primary drying), followed by desorption of the unfrozen water (secondary drying). 12 Freeze-drying, however, is damaging to biomolecules and can cause a collapse of the ECM structure and/or denaturation of sensitive proteins. Thus, it requires protective additives such as sucrose or trehalose to stabilize the scaffold structure and its macromolecular constituents and to prevent or reduce the adverse effects of freeze-drying.13,14 The use of protective substances for freeze-drying is inspired by nature's way to preserve life in the dried state as found in anhydrobiotic organisms such as tardigrades, nematodes, yeast, and seeds and pollens of higher plants. 15 Anhydrobiotic organisms accumulate large quantities of nonreducing disaccharides such as sucrose or trehalose before entering into the anhydrobiotic state. 16 The sugars form stabilizing hydrogen bonds with endogenous biomolecules and build a highly viscous glassy state. Sucrose and trehalose have found widespread applications as stabilizing lyoprotectants for freeze-drying of proteins and liposomes.17,18 More recently, these sugars have also been used for freeze-drying of more complex systems, including mammalian cells,19,20 and ECMs.21,22
The main objective of this study was to investigate histological and biomechanical characteristics of freeze-dried decellularized pulmonary heart valves stabilized by sucrose and hydroxyl ethylene starch (HES) as lyoprotectants. The rationale for using a mixture of sucrose and HES is that sucrose forms hydrogen bonds with tissue proteins in the dried state, whereas HES is used to elevate the glass transition temperature during the freeze-drying procedure. 23 Pulmonary heart valves were dissected from porcine hearts and decellularized by a solution of ionic-detergents. Decellularized valves were freeze-dried: (1) without lyoprotectants, (2) with 5% sucrose, and (3) with a mixture of 2.5% sucrose and 2.5% HES. Native valves, decellularized valves, and valves that were freeze-dried and afterward rehydrated underwent histological and biomechanical analyses.
Materials and Methods
Source of porcine valves
Porcine pulmonary valves were harvested from fresh hearts obtained from a local slaughterhouse and placed into phosphate-buffered saline (PBS) with 1% penicillin/streptomycin, 1% partricin, and 1% gentamycin (PBS+). Adherent fat and most of the adjacent myocardium and vessel were removed, leaving only a thin ridge of subvalvular muscle tissue and a small part (about 4 cm) of the pulmonary artery. Afterward, the valves were either immediately analyzed or further processed as described below.
Decellularization of porcine pulmonary heart valves
Before decellularization, tissues were disinfected with povidone iodide solution (Braunol) for 5 min and then washed with PBS+ for 20 min to prevent the tissue from fixation. The tissues were placed into the decellurization solution, which consisted of 0.5% (w/v) sodiumdeoxycholate and 0.5% (w/v) sodiumdodecylsulfate in bidistilled water, for two times 12 h at room temperature. To remove residual detergents, the heart valve tissues were subsequently washed two times in bidistilled water for 12 h and then transferred into PBS solution for 5 days, changing the solution every 12 h. All steps were conducted under continuous shaking with a rotary shaking incubator (GFL Shaking Incubator 3031 & 3032). The decellularized heart valve tissues were stored in PBS+ at 4°C until further processing.
Freeze-drying procedure
Before freeze-drying, decellularized heart valve tissues were incubated in bidistilled water, a 5% sucrose solution, or a mixture of 2.5% sucrose and 2.5% HES, at 37°C for 4 h. The valves were then transferred to Petri dishes and put on the shelves of an Epsilon 2-10D freeze-dryer, with temperature-controlled shelves (Christ). The freeze-dryer pilot unit has an ice condenser temperature of −85°C, with a closed temperature control system via synthetic heart-transfer media, stainless steel chamber. The samples (∼3 cm2) were placed on Petri dishes and subjected to slow freezing at a cooling rate of ∼1°C/min on the shelves of the freeze-dryer until the product temperature reached −30°C. Primary drying was conducted at −30°C in vacuum for 7 h and a pressure of 160 mTorr. For secondary drying the shelve temperature was increased to 20°C at a rate of 1°C/min at the same pressure. After freeze-drying, the valves were stored at 4°C. Dried valves were rehydrated with water for at least an hour before histological inspection or biomechanical testing.
The freeze-drying process was performed (1) in the absence of lyoprotectants, (2) with 5% (w/w) sucrose, and (3) using a mixture of 2.5% (w/w) sucrose and 2.5% (w/w) HES. Native and decellularized tissues were used as controls.
Histological analysis
Tissues were fixed in formalin (10%) for at least 24 h at 4°C. After fixation, the tissues were rinsed with distilled water, and dehydrated in 25% ethanol (two times 30 min) and 50% ethanol (two times 30 min). Samples were then put into a Leica TP 1020 automatic tissue processor (Leica). Tissues were subsequently put in 75% ethanol (two times 30 min), 90% ethanol (two times 30 min), 96% ethanol (two times 45 min), isopropanol (two times 45 min), and roticlear (two times 45 min), and, finally, the tissues were immersed in paraffin for 6 and 24 h.
After complete processing of the tissue, samples embedded in paraffin were cut into slices of 7 μm thickness using a manual rotary microtome (Reichert-Jung Autocut 2040 Microtome) and transferred to microscopy glass slides. Sections were deparaffinized and rehydrated through xylene and graded ethanol to distilled water. Slides were stained with hematoxylin and eosin (H&E; for general histology and structure), Movat's pentachrome stain (for collagen), and Elastica van Gieson stain (for elastin).24–27 After the staining procedures, all sections were dehydrated in graded ethanol, cleared in xylene, mounted using Corbit-Balsam, and finally analyzed and documented using an Olympus light microscope (BX40) with photo documentation software (Axiovision Rel 4.6 System).
Biomechanical testing
Before starting the biomechanical tests, the whole valves were cut along the commissures and expanded on hard paper. Samples were punched out of the pulmonary arterial wall with a specially designed punching tool along both longitudinal and circumferential orientations from each treatment group. The specimens were dumbbell shaped, 35×4 mm overall in size, and 12×2 mm in the black area for elongation measurements (Fig. 1).
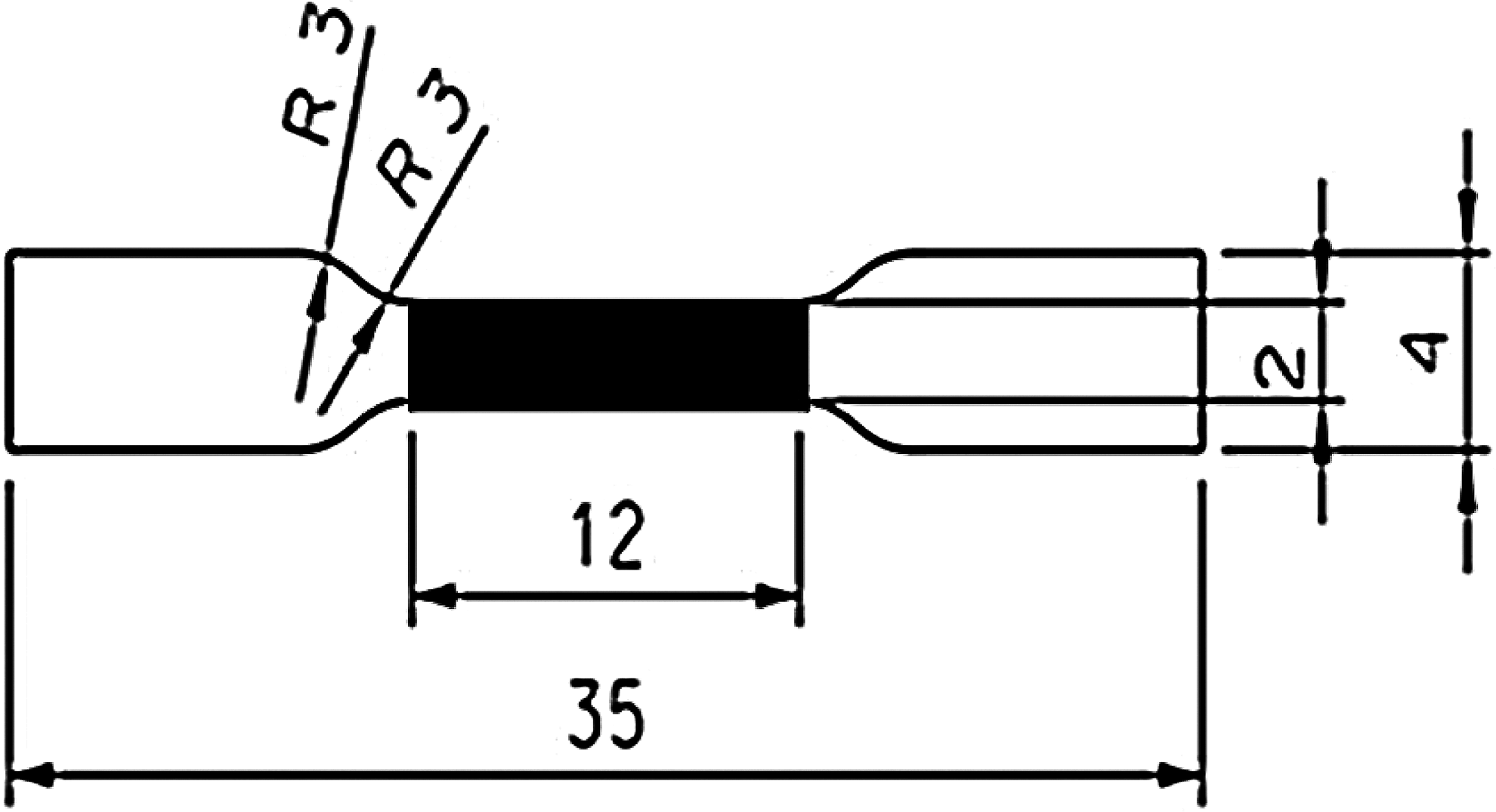
A schematic illustration showing the dimensions (in millimeters) of the tensile test sample (made by Nordpraezision Werkzeugfabrik GmbH). The measurement size of the dumbbell-shaped test sample of specimens within the black marked area has a length of 12 mm and a width of 2 mm. The radius of the curvature (R) is 3 mm.
Biomechanical testing was performed with a Zwick/Roell Z0.5 instrument. First, a force transducer with a loading threshold of 10 N was used to measure the thickness of each specimen. The punched samples were put on the testing plate of the instrument. The transducer descended from the starting position with a constant velocity of 15 mm/min until the head of the transducer reached a force of 0.02 N upon touching the surface of the sample. The thickness of the sample was measured as the distance between the head of the transducer and the plate. For tensile testing, a transducer with a maximum loading threshold of 200 N was used. The two end sections of the tissue specimens were each firmly held by grips with a rubber layer to prevent slipping. At the starting position, the distance between both grips was set at 12 mm (the length of the black marked area in Fig. 1).
Stress–strain curves were recorded after resetting the transducer to 0 N, at which point the tensile force is 0 N and the strain is zero. The tissue was loaded at 10 mm/min until rupture of the sample. This loading velocity is based on a modified procedure presented by Seebacher et al. 6
From the recorded load data, the stress was calculated as follows:
where σ equals the stress in N/mm2, F is the acquired force in newton (N), A is the cross-sectional area of the specimen in square millimeters (mm2), d is the width of the specimen (mm), and b is the thickness of the specimen (mm). The measured thickness values from each specimen were used to calculate the cross-sectional areas. Mechanical properties of the pulmonary artery tissue samples were analyzed during the elastic deformation phase until the changes became nonreversible. This point was taken as the maximum stress σmax (different from the rupture point of the tissue). The strain value at this point was ɛmax.
Strain was calculated from the extension data as follows:
where ɛ is the strain expressed in % of specimen elongation, Δl the extension of the specimen in mm, and l0 the original length of the specimen in mm at zero force. The slope in the stress–strain curves in the deformation region was used to calculate the elastic modulus.
Stress and strain values at the end of the elastic phase (σmax and ɛmax) were analyzed and compared between the different groups. All values are shown as mean±standard deviation. A Student's t-test was used to determine statistically significant differences between the groups. Calculations were done using Excel 2007. Tests among different groups resulting in a p-value<0.05 were considered significant.
Results
Heart valves were studied in five different treatment groups: native, decellularized, freeze-dried, freeze-dried with sucrose, and freeze-dried with sucrose/HES. Various histological staining methods were used to assess overall dimensional and structural changes; verify that cell nuclei were removed after decellularization; and assess the effects of processing on the leaflet trilayered structure and the collagen structure. Biomechanical testing experiments were done on pulmonary artery tissue.
Comparison of native pulmonary valves with decellularized valves
The valves consist of three defined layers: (1) the ventricularis layer, closest to the inflow surface, contains a meshwork of elastic fibers; (2) the fibrosa layer, at the outflow surface, contains collagen fibers, which are mostly aligned circumferentially; (3) the spongiosa layer lies in between and is largely composed of glycosoaminoglycans.1,28 H&E stain was used to verify the absence of cell nuclei after decellularization. Cell nuclei are visible as black spots in native tissue in pulmonary valves (Fig. 2A). Figure 2B shows the absence of cell nuclei after decellularization. Movat's pentachrome stain was used to elucidate the trilayered leaflet structure of the valves composed of collagen, elastin, and glycosaminoglycans (Fig. 2C, D). Elastica van Gieson stain was used for high-contrast visualization of elastic fibers within the connective tissue. Decellularization resulted in a thinner collagen layer (Fig. 2E, F).
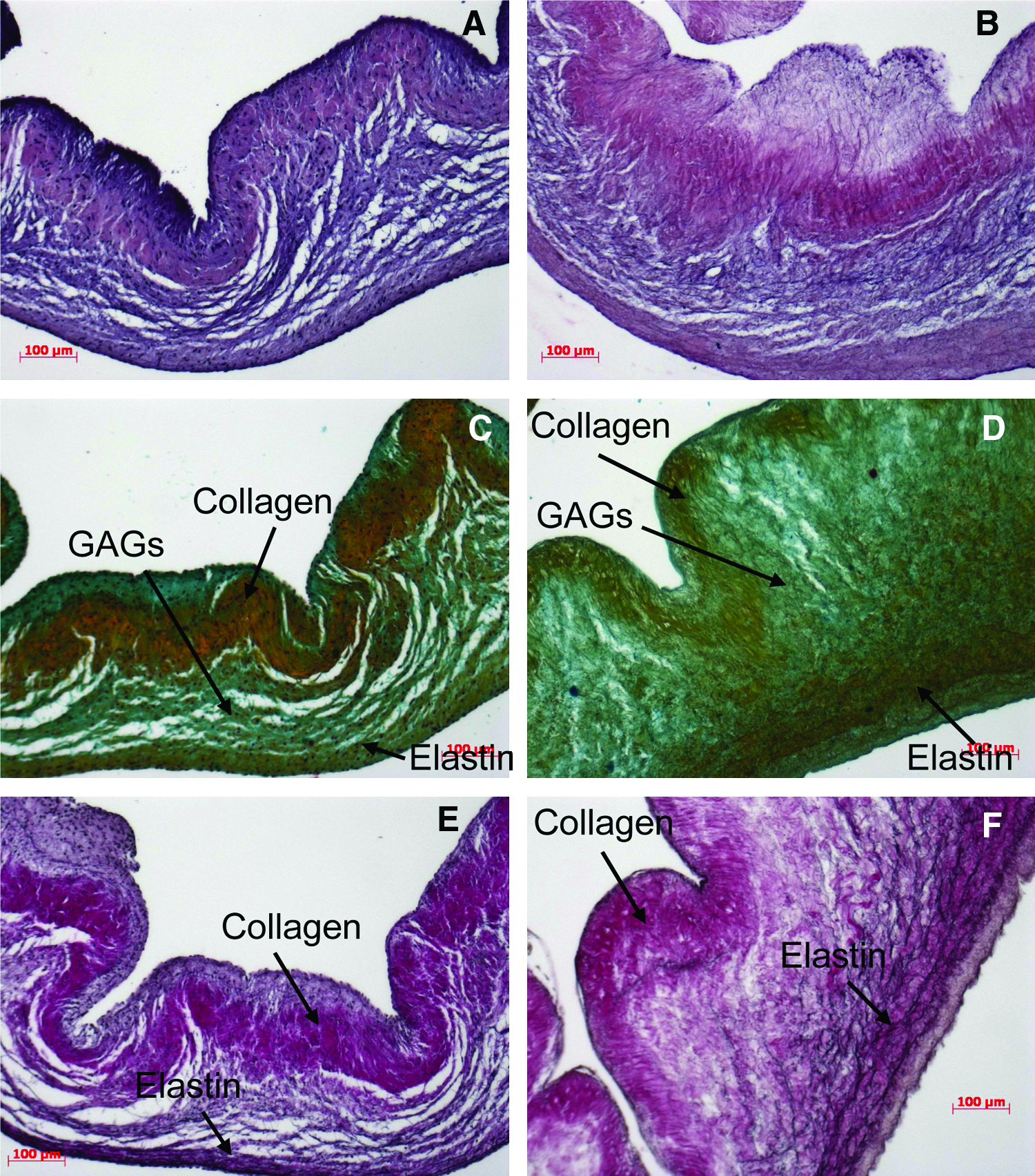
Histological stains of pulmonary heart valve leaflets. Comparison of native specimens
Effects of freeze-drying on decellularized pulmonary heart valves
Figure 3 shows the histology of valve leaflets after freeze-drying and rehydration. Figure 3A–C shows the layered structure of the decellularized valve leaflet with the ventricularis layer, the spongiosa layer, and the fibrosa layer using H&E staining, Movat's pentachrome staining and Van Gieson staining. Tissue that has been freeze-dried without lyoprotectants shows an overall collapse in ECM structure with only a few collagen-containing structures detectable within the ventricularis layer (Fig. 3D–F). The leaflets freeze-dried with 5% sucrose (Fig. 3G–I) and those freeze-dried with a mixture of 2.5% sucrose and 2.5% HES (Fig. 3J–L) show a more loose structure and partial detachment of the fibrosa from the spongiosa layer. Figure 4 shows cross sections through the vessel wall of pulmonary arteries before and after freeze-drying. The collapse of freeze-dried artery tissue in the absence of lyoprotectants is not as drastic as within the valve-leaflet structures. In the absence of lyoprotectants, however, artery tissue displays a greater porosity compared to tissue freeze-dried in the presence of sucrose. Tissue freeze-dried with sucrose alone reveals less porosity compared to tissue freeze-dried with the mixture from both of the used lyoprotectants—for the valve leaflet as well as for the pulmonary artery tissue.
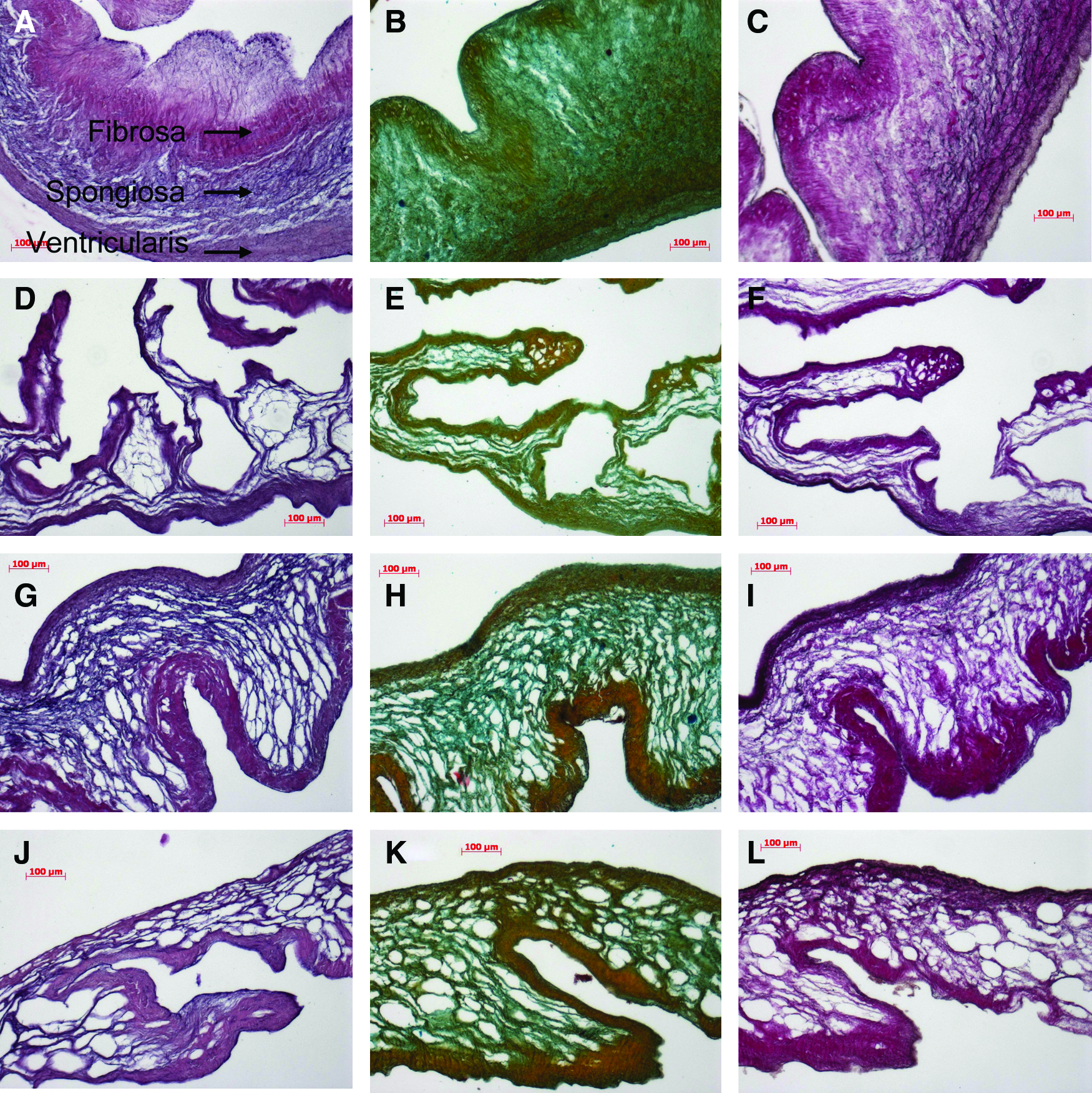
Histological stains of pulmonary heart valve leaflets. Decellularized specimens
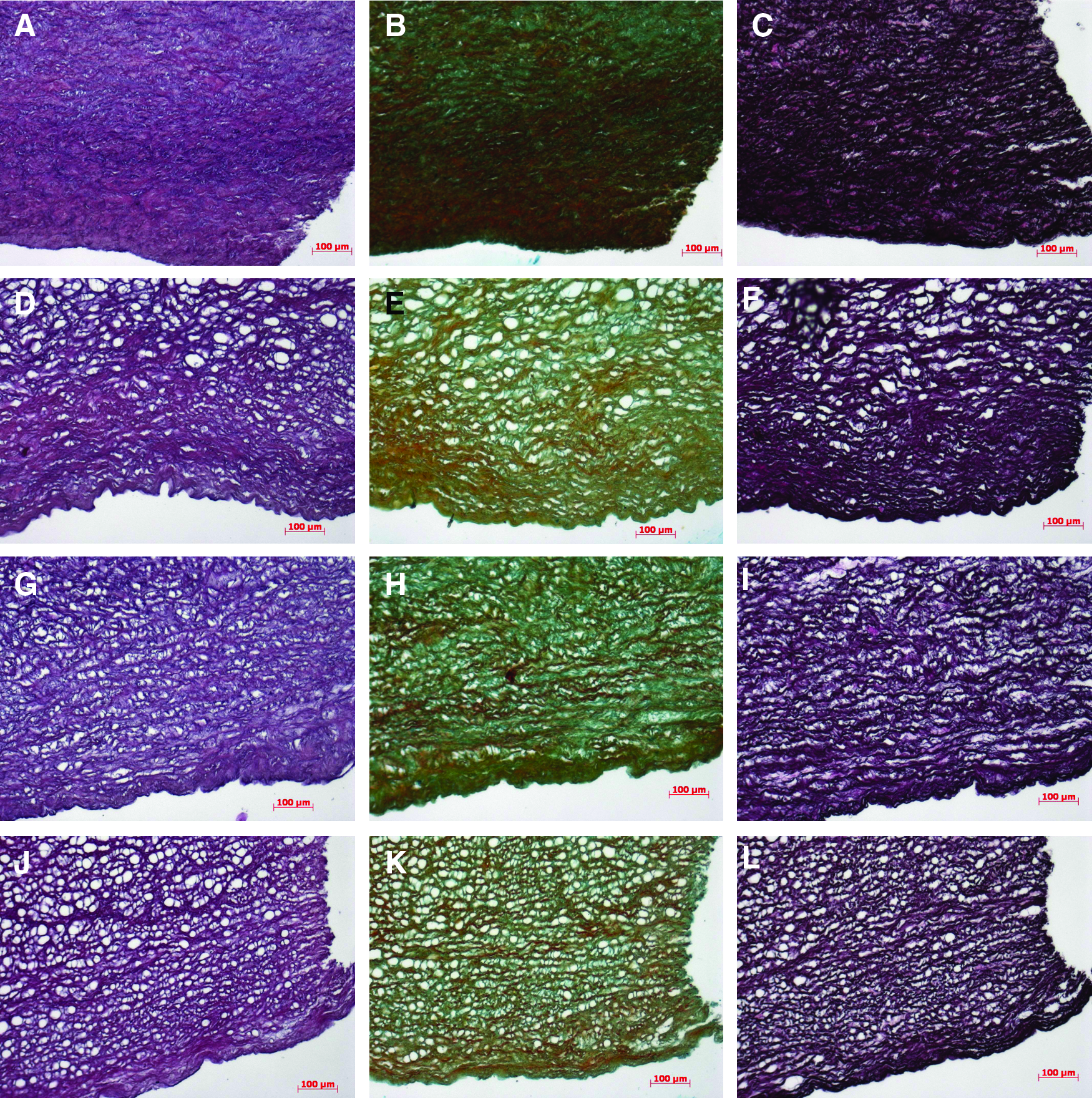
Histological stains of pulmonary artery tissue. Decellularized specimens
Biomechanical testing
Biomechanical tissue properties of pulmonary arteries were tested before and after decellularization, as well as after freeze-drying and rehydration. First, the thickness of the tissue samples was measured. On visual inspection, pulmonary arteries appear to be fairly homogeneous and distinct from surrounding connective tissue. Thus, the inhomogeneity in thickness is considered negligible. One of the end areas of the sample was used for the thickness measurements. The thickness of native tissue, decellularized tissue, freeze-dried tissue without lyoprotectant, freeze-dried tissue with sucrose, and tissue freeze-dried with a mixture of sucrose and HES were determined to be 2.02±0.24, 1.48±0.37, 1.56±0.38, 1.63±0.37, and 2.06±0.32 mm, respectively. The elastic modulus was calculated from the slope in the stress–strain curves in the collagen phase as indicated in Figure 5. The E-modulus values of the specimens in circumferential and in longitudinal direction are shown in Figure 6. Decellularization significantly decreases the E-modulus in longitudinal as well as in circumferential direction. Interestingly, the biomechanical properties of the leaflets freeze-dried in the absence of lyoprotectants are not significantly different from those of the decellularized tissue. The E-modulus (circumferential direction) of the sample dried with sucrose is greater compared to that of the decellularized control tissue (*p<0.05, Fig. 6), but similar to that of native tissue.

Stress–strain curve of native pulmonary arteries in circumferential direction illustrating the elastin phase, transition phase, and the collagen phase. The slope in the collagen phase is used to calculate the elastic modulus. The maximum stress (σmax) and maximum strain (ɛmax) values at the end of collagen phase have been determined for the different treatment groups.
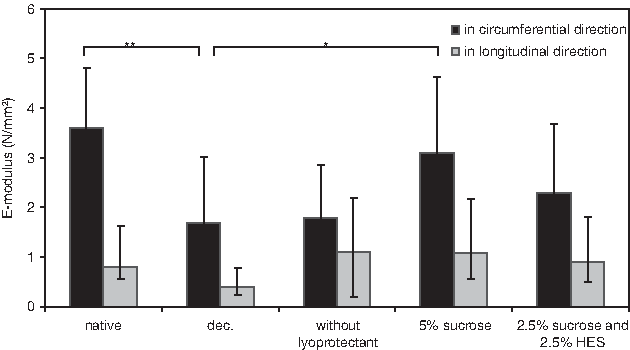
Comparison of E-modulus in circumferential (black bars) and in longitudinal (gray bars) direction of pulmonary artery tissue: native, decellularized, freeze-dried and rehydrated, freeze-dried with 5% sucrose and rehydrated, and freeze-dried with 2.5% sucrose/2.5% HES and rehydrated. The data are mean values from 10 to 12 experiments with corresponding standard deviation. Significant differences between groups are indicated as *p<0.05 and **p<0.01.
The stress and strain values at the end of the collagen phase are shown in Figure 7 (σmax and ɛmax in the stress–strain curves as indicated in Fig. 5). Note that this is not the rupture point of the tissue. Statistically significant differences in maximum stress values were observed between native and decellularized specimens, indicating that decellularization affects biomechanical tissue properties of arteries. Compared to native arteries, the maximum stress level of decellularized arteries is significantly smaller in circumferential direction (***p<0.001, Fig. 7A). The maximum stress level of arteries freeze-dried with sucrose is significantly greater compared to decellularized tissue, but not significantly different compared to native tissue. No statistically significant differences were observed between samples freeze-dried with 5% sucrose and those that were freeze-dried with a mixture of 2.5% sucrose and 2.5% HES. In longitudinal direction, the maximum stress levels are slightly increased in the freeze-dried groups (Fig. 7B). The maximum strain level in circumferential direction at which tissue failure is reached of native samples is 113.9%±30.6% compared to 102.9%±26.1% in decellularized tissues (Fig. 7C). Maximum strain levels of arteries, however, did not show clear differences among the different groups neither in circumferential nor in longitudinal direction (Fig. 7C, D).

Maximum stress (σmax) and maximum strain (ɛmax) of pulmonary arteries in circumferential
Discussion
Successful scaffold materials to generate a valve substitute that reproduces the function of the native valve should have a controlled degradation, be compatible with cells, lack cytotoxicity, and not elicit an immune or inflammatory response. 29 An engineered scaffold should possess cell attachment and signaling molecules that will promote cell population and function, resulting in remodeling of the scaffold into a tissue construct, which can mimic the function of the native valve. Such molecules are naturally present in biological scaffolds. Decellularized porcine pulmonary valves can be used to obtain viable and biomechanically stable constructs, resembling native aortic heart valves using ovine cells and dynamic tissue culture conditions. 30 The aim of this study was to analyze the effects of freeze-drying on decellularized porcine pulmonary heart valves (leaflet and vessel tissue). Freeze-drying in the absence of lyoprotectants caused an overall more disintegrated appearance of the histological architecture of the valve leaflets, especially between the fibrosa and the ventricularis layers. The histological architecture is better preserved when freeze-drying is done in the presence of lyoprotectants. Tissues freeze-dried with lyoprotectants have a looser network of collagen and elastic fibers with bigger pore sizes as opposed to the denser collagen network and decreased pore size of decellularized tissues. The increased pore size of freeze-dried tissue could be advantageous for the adherence of circulating stem cells. Cells may adhere better in the more porous freeze-dried tissue. Further studies, however, are needed to test this idea.
Like most soft biological materials, blood vessels do not have simple linear elastic properties. They exhibit nonlinear behavior when distended, so elastic stiffness increases with the degree of loading. 31 This nonlinear behavior has also been observed for biological scaffolds made from decellularized skin tissue. 32 The essential elastic nonlinearity of the artery wall has been shown in a classic study by Roach and Burton. 33 They showed that the initial stiffness of the artery wall reflects the elasticity of elastin, whereas the much higher stiffness at higher strains can be attributed to the contribution of fully tensed collagen fibers. Collagen is one of the most important component of pulmonary arteries that determines its biomechanical properties. 34 Much of the behavior in the collagen phase is a direct result of changes in internal structure by strain, which involves both straightening of highly crimped collagen fibers and rotation of these fibers toward the stretch axis. 35
Decellularization caused a decrease in the area and thickness of the pulmonary arterial wall, which could explain the reduced biomechanical stress of the tissues. Decellularized arteries have reduced stress and strain properties compared to native arteries, particularly in circumferential direction. Biomechanical properties of tissue freeze-dried in the absence of lyoprotectants were similar to those of decellularized tissue. Interestingly, in the presence of lyoprotective agents, the elastic modulus after freeze-drying and rehydration was found to be similar compared to native tissue. The slightly greater elastic modulus of the tissue freeze-dried with sucrose alone compared to that of the tissue dried with the mixture (not significant) coincides with a reduced porosity in the tissue dried with sucrose. Likely, the collagen undergoing freeze-drying with sucrose alone is more condensed than in the other situations, which might explain the increased elastic modulus. Narine and coworkers 36 examined the feasibility of cryopreservation for storage of porcine aortic valve matrices. Their studies showed extensive disruption of the extracellular collagen matrix and inter-fibrillar proteoglycan associations after cryopreservation. They showed that cryopreservation does not significantly alter the biochemical properties of porcine aortic valve matrices. Cryopreserved matrices, however, showed significantly higher strain when stressed compared to noncryopreserved matrices.
Taken together, freeze-drying could provide a promising tool for the preservation of ECM structures and storage of heart valves. The histological tests have shown beneficial effects of adding lyoprotectants to protect the tissue during the freeze-drying procedure. Freeze-dried tissue, however, has a more porous structure compared to native or decellularized tissue. Qualitatively, tissue freeze-dried with sucrose looked more intact than tissue freeze-dried using a sucrose/HES mixture. No significant differences in biomechanical properties were observed, however, between tissue that was freeze-dried using sucrose and tissue freeze-dried with a sucrose/HES mixture. Biomechanical tests show that decellularization significantly affects the biomechanical properties of the tissue. Interestingly, the biomechanical properties of the valves freeze-dried with sucrose more closely resemble those of native tissue than the decellularized tissue. The in vivo performance and durability of freeze-dried heart valve constructs remains to be elucidated. Conduit arteries become stiffer with age due to alterations in their morphology and the composition of the their major structural proteins, elastin and collagen. 37 Elastin and collagen may become stiffer due to calcification and the formation of cross-links due to advanced glycation end-products. 37 Further studies are needed to demonstrate in vitro and in vivo performance of freeze-dried tissues.
Footnotes
Acknowledgments
We thank Doreen Unger for excellent technical support. This work was financially supported by the German Research Foundation (Deutsche Forschungsgemeinschaft, DFG), Cluster of Excellence “From regenerative biology to reconstructive therapy” (REBIRTH), and by the German Federal Ministry of Economics and Technology (AiF Project KF2654702FO0).
Disclosure Statement
No competing financial interests exist.