
Abstract
To study human skin pigmentation in a physiological in vitro model, we developed a pigmented reconstructed skin reproducing the three-dimensional architecture of the melanocyte environment and the interactions of melanocyte with its cellular partners, keratinocytes, and fibroblasts. Co-seeding melanocytes and keratinocytes onto a fibroblast-populated collagen matrix led to a correct integration of melanocytes within the epidermal basal layer, but melanocytes remained amelanotic even after supplementation with promelanogenic factors. Interestingly, normalization of keratinocyte differentiation using keratinocyte growth factor instead of epidermal growth factor finally allowed an active pigmentary system to develop, as shown by the expression of key melanogenic markers, the production, and transfer of melanosome-containing melanin into keratinocytes. Various degrees of constitutive pigmentation were reproduced using melanocytes from different skin phenotypes. Furthermore, induction of pigmentation was achieved by treatment with known propigmenting molecules, αMSH and forskolin, thus demonstrating the functionality of the pigmentary system. This pigmented full-thickness skin model therefore represents a highly relevant tool to study the role of cell–cell, cell–matrix, and mesenchymal–epithelial interactions in the control of skin pigmentation.
Introduction
Even though the role of the dermal compartment on pigmentation is far less documented, there is now evidence that the mesenchymal compartment, including fibroblasts and fibroblast-derived extracellular matrix (ECM) proteins, influences melanocyte proliferation, apoptosis resistance, morphology, and melanogenic activity.12–25 Melanocyte adhesion and location at the dermal epidermal junction also depend on the microenvironment. In fact, several ECM proteins such as laminin or collagen IV are known to play a role in attachment of melanocytes onto the basement membrane via the interaction with specific integrins. 16 Dermal fibroblasts play a regulatory role in constitutive pigmentation through the secretion of soluble factors. Some of those are the same as those secreted by keratinocytes such as HGF or SCF, and can be released in even greater amounts by fibroblasts than by keratinocytes.20,24 Some are specifically secreted by fibroblasts, as illustrated by Dickkopf-1 (DKK1), which is responsible for the very light color of the palms and soles via a suppressive effect on melanocyte growth and activity as well as melanin uptake by keratinocytes.23,25 More recently, the propigmenting effect of Neuregulin-1 secreted by fibroblasts derived from dark skin suggests its involvement in determining the human skin color. 15 Furthermore, paracrine cytokine regulation loops may also exist; for example, keratinocyte-produced cytokines, such as IL1α or TNFα, may stimulate fibroblasts, which in turn release melanocyte-stimulating factors such as HGF or SCF.20,26,27
Deregulations of melanocyte homeostasis and/or melanogenesis are the cause of various hyper- or hypopigmented lesions. For a better understanding of the pigmentation mechanisms and their deregulations, in vitro systems that reproduce the physiology of the pigmentary system as close as possible are required. The development of in vitro skin-engineered substitutes for biological, photobiological, and pharmacological research purposes has made significant progress over the years.28,29 These models reproduce key structures of native skin, especially a three-dimensional (3D) organization and a differentiated epidermis. The successful integration of normal melanocytes provided a significant improvement of pigmented models, since pigmentation could be thus studied in conditions reproducing the EMU.9,30,31 To overcome the absence of fibroblasts in these reconstructed epidermis, more complex organotypic pigmented skin models reconstructed on a dermal equivalent containing fibroblasts have been designed. Different fibroblast-containing dermal substrates have been used such as recolonized de-epidermized dermis (DED),14,19,21,32 a glycoaminoglycan–collagen sponge, 33 or a fibroblast-populated collagen gel.34–36 However, some of these models can hardly be considered a human normal skin model, because they contain melanocytes from mouse origin grown in a growth medium containing TPA, known to induce phenotypic and functional cellular alterations.35–37 Beside, some researchers have developed disease skin models using non-normal cells such as melanoma or congenital café au lait macules cells.34,35,37 Even though the integration of normal human melanocytes (NHMs) was achieved in the skin substitutes, key pigmentation parameters that could demonstrate that the pigmentary system is physiological (presence of melanogenic proteins, melanin production, and stimulation of pigmentation) have not been checked. Even more, some drawbacks were noticed. Okazaki et al. and Souto et al. cited an insufficient differentiation of the epidermis with an absence of the stratum granulosum and corneum.32,35 Other defects concern the pigmentary system with a limited survival or a deficient functionality of melanocytes21,14 or to the unresponsiveness of the model to physiological propigmenting agents. 19
To study human skin pigmentation in a highly physiological in vitro model, our objective was to reconstruct a pigmented skin model (including human normal melanocytes, keratinocytes, and fibroblasts endowed with a functional pigmentary system) that is able to develop a real constitutive pigmentation (melanin production and transfer) and able to respond to known stimulators.
We first took advantage of our experience of (1) reconstructing skin models with a fibroblast-embedded collagen gel 38 and (2) reconstructing pigmented epidermis9,31 and (3) combining both technologies to succeed in integrating melanocytes in the complete reconstructed skin. To stimulate melanin synthesis, various culture conditions were tested, supplementation with promelanogenic growth factors or modification of the keratinocyte homeostasis, resulting in (1) a correct integration of melanocytes and (2) a physiological melanocyte differentiation that allowed us to reproduce various pigmentation phenotypes. Finally, the functionality of the pigmentary system was verified by stimulation with the known propigmenting agents, αMSH and forskolin.
Materials and Methods
Cell culture
Epidermal normal human keratinocytes (NHKs) were isolated from Caucasian breast skin obtained from plastic surgery procedures, cultured as described 39 on a feeder layer of Swiss 3T3 fibroblasts, and used at passage 2. Human dermal fibroblasts were isolated after spreading from Caucasian mammary skin explants, amplified in the Dulbecco's modified Eagle's medium+10% fetal calf serum (FCS), and used at passage 8. NHMs were obtained from young foreskins of four different donors selected according to their pigmentation phenotype. Three Caucasian donor skins were lightly, moderately, darkly pigmented, and one African donor skin was black. Isolated melanocytes were amplified in a defined melanocyte culture medium M2 (Promocell) containing no phorbol ester and used at passages 4–6.
Skin reconstruction
NHMs, NHKs, and fibroblasts were amplified separately in their respective growth medium.
The dermal equivalent was obtained after contraction of a mixture of bovine type I collagen and fibroblasts (106 cells for 7 mL) as described previously.38,40 NHKs and NHMs were co-seeded on the contracted lattice at a concentration of 33,000 cells/cm2 each: for this, 50,000 melanocytes and 50,000 keratinocytes were seeded inside a 1.5-cm2 steel ring laid on the dermal equivalent. The culture was then kept submerged for 7 days, allowing cells to form a monolayer. The culture was then raised to the air–liquid interface (emersion phase) and kept at least for 1 week to allow the keratinocytes to stratify and differentiate. The culture medium used for the immersion and emersion phases was composed of the Minimal Essential Medium (Invitrogen), 10% FCS (Sigma), 10 ng/mL epidermal growth factor (EGF) (Becton Dickinson), 10−10 M Cholera toxin (Biomol), and 0.4 μg/mL hydrocortisone (Sigma) and was supplemented with M2 melanocyte growth factors (0.625 mL/L). Growth factors were SCF (10 ng/mL; R&D Systems), ET3 (100 nM; Sigma), bFGF (5 ng/mL; Sigma), and keratinocyte growth factor (KGF, 10 μg/mL; R&D Systems).
Induction of pigmentation by propigmenting agents
Reconstructed skin containing melanocytes from Caucasian moderately pigmented skin was made to test two propigmenting references, α-MSH (Sigma) and forskolin (Sigma). α-MSH and forskolin were added to the culture medium at the final concentration of 50 nM and 40 μM, respectively. They were renewed every second day. The treatment started on the first day of emersion phase (ED0) and ended 18 days after emersion (ED18). Another experiment was performed by adding 40 μM forskolin as soon as from the seeding of the keratinocytes and melanocytes onto the dermal equivalent.
Melanocyte morphology and distribution
After separation of the reconstructed epidermis from the dermis using forceps, dihydroxyphenylalanine (DOPA) reactions were performed as previously described, 41 to visualize the melanocyte morphology and distribution throughout the surface of the epidermis.
Histology and immunostaining
Samples were fixed in 10% neutral formalin and treated for histology. Five- micrometer paraffin sections were stained with hematoxylin, eosin, and saffron. Melanin was colored by Fontana-Masson (FM) staining.
For immunolabeling, samples were embedded in Tissue Tek (Miles) and frozen in liquid nitrogen, or embedded in paraffin. Five-micrometer cryosections were postfixed in methanol at −20°C. Paraffin sections were deparaffined after several xylene/ethanol baths, and epitope demasking was performed by heating at 95°C for 20 min in a citrate buffer (pH 9). To detect melanogenic proteins, primary monoclonal antibodies (MoAb) NK1-beteb specific for Pmel-17 (Monosan), T311 specific for human tyrosinase (Novocastra), Ta99 specific for human tyrosine related protein-1 (TRP-1) (Signet), D5 specific for MITF (Neomarkers), and a polyclonal antibody D18 specific for human TRP-2 (Santa Cruz) were used.
For fluorescent immunochemistry, NKI/beteb (dilution 1/5), which recognizes Pmel-17, and Ta99 (dilution 1/50), which recognizes TRP-1, were both applied 30 min at room temperature (RT) on cryosections. Then, Alexa-488-coupled secondary antibodies (Molecular Probes) were applied for 30 min at RT. Nuclear counterstaining was performed with propidium iodide (PI).
For the enzymatic detections of tyrosinase by T311 (1/10), TRP-2 by D18 (1/100) on cryosections, and MITF by D5 (1/40) on paraffin-embedded section, biotinylated species specific of the secondary antibody were applied overnight at 4°C and revealed using the streptavidin–biotin peroxidase complex (LSAB2; Dako). 3-amino-9-ethylcarbazole (AEC) (Dako) or Histogreen (Vector Lab.) were used as enzymatic substrates. Counterstaining was performed with Papanicolaou Blue for AEC–LSAB2 staining and with Nuclear fast red (Vector Lab.) for Histogreen–LSAB2 revelation. Histogreen sections were dried after being passed through ethanol and xylene baths. AEC- and Histogreen-revealed slides were respectively mounted with immunomount (Thermo) and Vectamount (Vector Lab.).
For fluorescent co-staining of fibroblasts and melanocytes on cryosections, fibroblasts were first labeled with a WM15 MoAb specific of the CD13 antigen (dilution 1/10; AbDSerotec), which was applied for 30 min at RT, and which was followed by Alexa-488-coupled secondary antibody (dilution 1/150, 30 min RT; Molecular Probes). Melanocytes were fixed with methanol before labeling with Ta99 (dilution 1/50) and Alexa-568-coupled secondary antibody (dilution 1/200; Molecular Probes), both applied for 30 min at RT. Finally, nuclear counterstaining was performed with Hoechst dye (dilution 1/2500; Invitrogen). Negative controls have been performed by omitting primary antibodies.
The percentage of melanocytes present in basal layer and the Melanocyte/Keratinocyte ratio were assessed at ED14 by determining the ratio of TRP-1-stained cells to the number of total basal cells on cryosections. The number of total cells was estimated by the count of PI-labeled nuclei. Measurements were taken from three to five different fields for each sample (magnification×100), and in three independent experiments, each of which were performed in duplicate. The value was expressed as mean with the standard error of the mean.
Transmission electron microscopy
Samples were fixed for 1 h in 4% glutaraldehyde, then washed in PBS, and postfixed for 90 min in 2% osmium tetroxide. Samples were dehydrated and embedded in a Spurr resin. The resin was polymerized at 60°C. Eighty-nanometer-ultrathin sections were counterstained with uranyl acetate and lead citrate and observed with a JEOL 1010 electron microscope.
Colorimetric measurements
A Mercury 2000 spectrocolorimeter (Datacolor) was used to measure the luminance (L*) of pigmented reconstructed skin samples (three to six samples in each experiment) in the CIE L*a*b* color space. The lower the L* value, the darker the color of the skin sample.
Melanin content
Melanin pigments revealed by FM staining on 5-μm paraffin sections were quantified by an image analysis using the Pathfinder™ Cellscan morphoscan platform associated to CellMarker software. Melanin content is expressed as the percentage of melanin surface/epidermis surface.
Statistical analysis
Luminance and melanin content values are expressed as mean±standard deviation calculated for three to six different samples in each experiment and analyzed using the two-tailed unpaired Student's t-test.
Results
Induction of constitutive pigmentation
Integration of melanocytes in the reconstructed skin model
To develop a human pigmented skin model, NHKs and NHMs from Caucasian moderately pigmented skin were firstly amplified independently and then were co-seeded onto a dermal equivalent composed of a contracted fibroblast-embedded collagen gel (lattice). NHKs and fibroblasts were both from Caucasian skins. The skin was reconstructed in an EGF-based keratinocyte growth medium as described in the Material and Methods section. Several cell concentrations (from 33,000 to 300,000 keratinocytes/cm2 and from 6700 to 33,000 melanocytes/cm2) and different NHK/NHM ratios (from 1/10 to 1/1) were tested (data not shown). These preliminary trials allowed us to select the optimal seeding concentration for each cell type, eventually set at 33,000 keratinocytes/cm2 and 33,000 melanocytes/cm2. Using these conditions, a skin model was obtained 7 days after emersion (ED7) with (1) a good epidermal structure (Fig. 1a) and (2) a correct integration of melanocytes in the basal layer revealed by immunostaining of TRP-1, a specific melanosome marker (Fig. 1b). DOPA reaction performed on the epidermal sheet showed that melanocytes were homogeneously distributed and maintained with a mostly bipolar morphology (Fig. 1c). However, using FM staining, no melanin pigment was visible neither in the melanocytes nor in the neighboring keratinocytes, indicating that melanocytes were inactive (Fig. 1d).
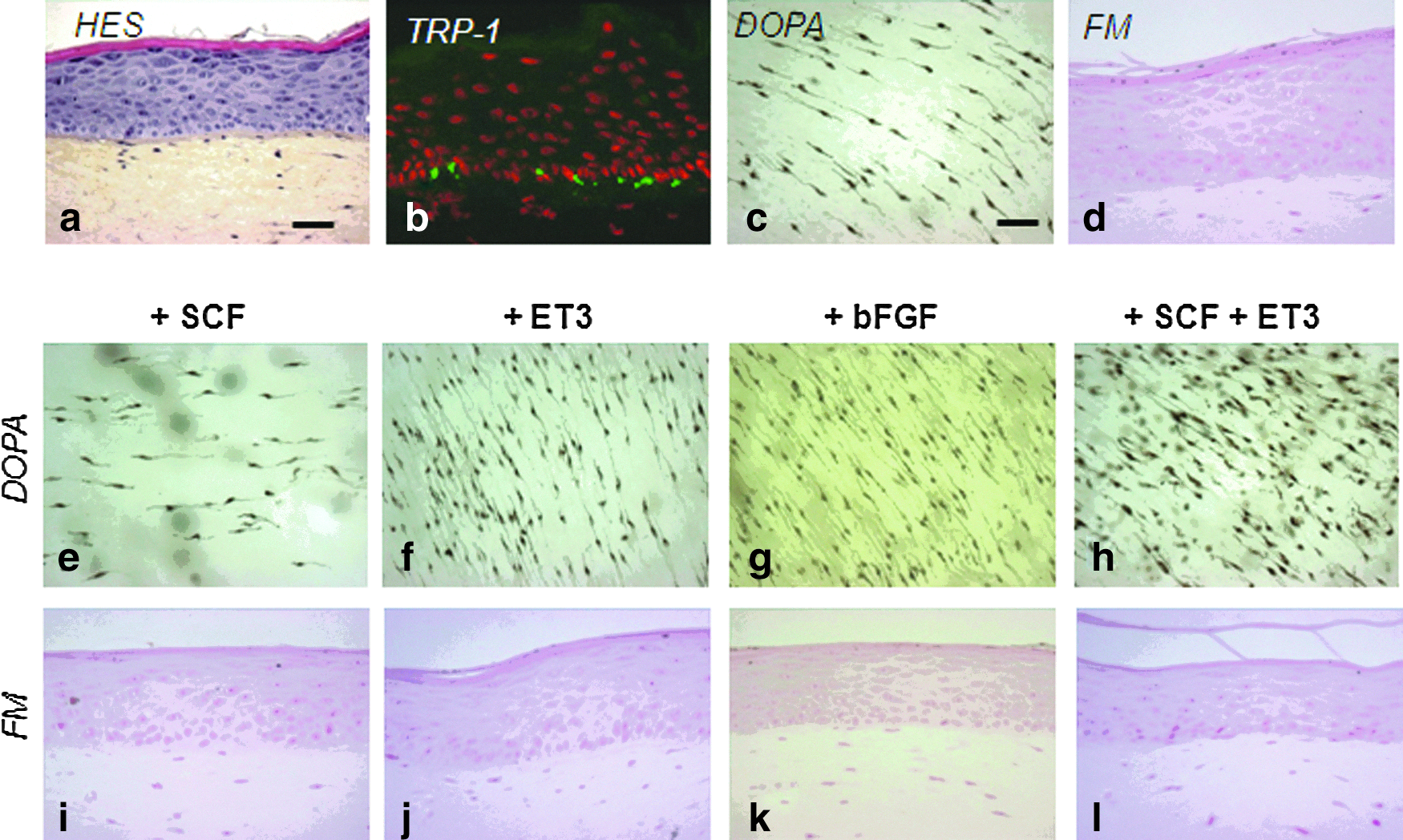
Integration of melanocytes and melanin synthesis in reconstructed skin: effect of various melanogenic growth factors.
Effects of melanogenic growth factors on constitutive pigmentation
To stimulate the differentiation of melanocytes and melanin synthesis, various growth factors were used to supplement the skin growth medium. In the presence of SCF, ET3, and bFGF, largely known to influence melanocytes and melanogenesis, the density and/or morphology of melanocytes was modulated. While the melanocyte density was increased by ET3 and bFGF, it was decreased by SCF alone (Fig. 1e–g). When combined with ET3, SCF induced an increase in the number of melanocytes, but an adverse cytotoxic effect was observed as shown by the round-shape morphology (Fig. 1h). However, in all conditions tested, the melanin pigment was still not detectable in the epidermis as shown by FM staining (Fig. 1i–l).
Induction of melanin production in the reconstructed skin cultured with KGF
Since keratinocytes are recognized for promoting melanocyte stimulation, we tested a growth factor, KGF, which is known to act on keratinocyte differentiation. Replacement of EGF by KGF, 4 days after NHK-NHM seeding, did not modify the basic architecture of the skin model that displayed a correct epidermal differentiation process with horny layer formation (Fig. 2a). Interestingly, KGF induced a normalized or an increased expression of differentiation markers such as K10 and loricrin at an early time ED7, compared to those observed with EGF (Supplementary Fig. S1; Supplementary Data are available online at

Stimulation of melanin production in reconstructed skin cultured with keratinocyte growth factor (KGF). Pigmented reconstructed skin (PRS) was cultured in KGF (10 ng/mL) instead EGF (10 ng/mL) containing a medium from the 4th day after epidermal cell seeding to the 7th or 14th day after emersion (ED7 or ED14): HES staining
Characterization of the pigmentary system in the reconstructed skin
The pigmentary system was further characterized on the reconstructed skin model cultured in a KGF-containing medium, at ED14. DOPA reaction on epidermal sheet showed that melanocytes were homogeneously distributed and developed a dendritic shape (Fig. 3a, b), compared to that of melanocytes in vivo. The percentage of melanocytes present in the basal layer was calculated by measuring the number of TRP-1 positively stained cells compared to the number of unstained cells (keratinocytes) (see Materials and Methods section). We found that 5.88%±0.39% of the cells present in the basal layer were melanocytes, which corresponds to a melanocyte/keratinocyte ratio of around 1/18.

Characterization of the pigmentary system in the reconstructed skin. The pigmentary system in the pigmented reconstructed skin (PRS) has been analyzed at ED14 using
The transfer of melanosomes within keratinocytes was studied using transmission electron microscopy. Detected ultrastructurally in between keratinocytes, dendrites contained stage III and stage IV mature melanosomes, which correspond to melanosomes with melanin deposits along fibrous striations or full of melanin, respectively (Fig. 3c). Interestingly, individual or clusters of mature melanosomes were observed within the keratinocytes (Fig. 3d).
The pigmentary system was characterized using immunolabeling of melanogenic proteins, Pmel-17, Tyrosinase, TRP-1, TRP-2, and the key melanogenic transcription factor MITF.
They confirmed that melanocytes were located at the basal layer and revealed that the pigment cells expressed Pmel-17, Tyrosinase, TRP-1, TRP-2, and MITF in the same way as they do in vivo (Fig. 3e–n). To assess the life span of the model, several parameters were evaluated using skin samples left in the emersion phase until ED28 (Supplementary Fig. S2). Although cell proliferation revealed by Ki67 staining was maintained until ED28, we found that the morphological qualities of the epidermis—global histological architecture, organization, and thickness of the viable epidermis—did not rise a sufficient level later than ED18. Therefore, the melanocyte behavior was analyzed from ED7 to ED18. Actually, melanocyte density, location, and activity assessed, respectively, by DOPA reaction, TRP-1, and FM staining, were constant all over this period. Precisely, the percentage of melanocytes at the basal layer at ED18, equal to 6.20±0.49 (corresponding to a melanocyte/keratinocyte ratio of 1/16), was very close to the one calculated at ED14 (see above). Altogether, the optimal lifespan of the model has been estimated to be ED18.
Reconstruction of skins of different phenotypes
We addressed the issue of integrating melanocytes from various constitutive pigmentation donors into the skin model. Four melanocyte strains were isolated from light to black skins and amplified before being used for skin reconstruction. A single keratinocyte and fibroblast strain from Caucasian donors was used. Samples were analyzed at ED14. As illustrated in Figure 4, all melanocyte strains, from the lighter to the darker, were correctly integrated and located into the skin model, and produce melanin. In correlation with the original skin phenotype, the color of skin samples varied gradually from light to dark. Luminance measurements and quantification of the FM staining confirmed that the more pigmented the melanocytes, the darker the skin, and the higher the amount of melanin.
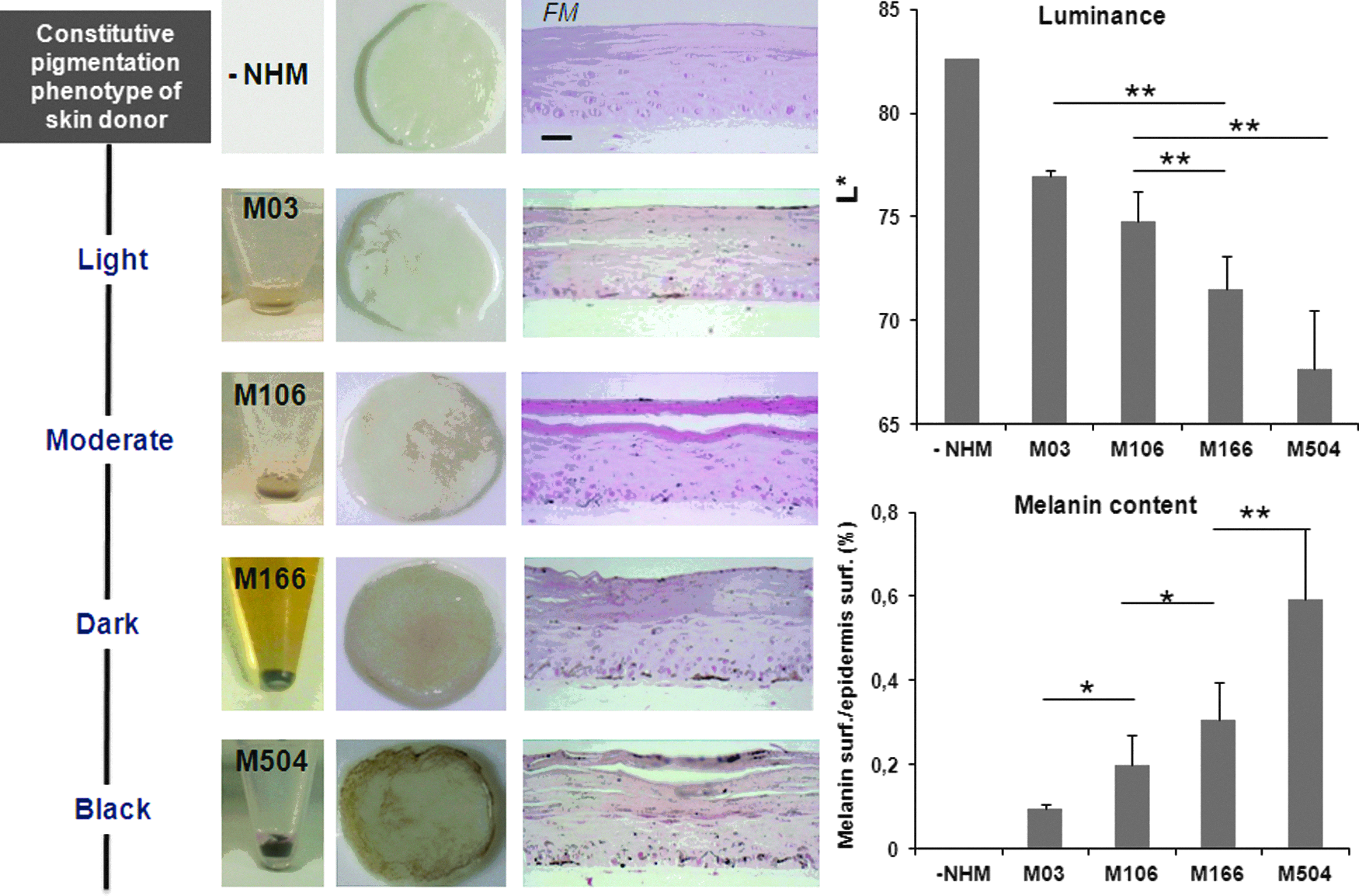
Reconstruction of skins of different phenotypes. Melanocyte strains (M03, M106, M166, and M504) isolated from increasing constitutive pigmentation skins (light to black) have been integrated into the reconstructed skin containing same keratinocytes and fibroblasts. For comparison, a reconstructed skin containing no melanocytes (NHMs) is shown. At ED14, the macroscopic pictures of the samples, the melanin content revealed by FM staining on histological section, and quantified by image analysis, as well as the Luminance value (L*) showed that the more pigmented the melanocytes, the darker the reconstructed skin and the higher the amount of melanin. Two tailed Student's t-test *p<0.05, **p<0.01. Scale bar=25 μm.
Induction of pigmentation by propigmenting agents
To check the functionality of the pigmentary system in the skin reconstructs containing melanocytes from moderately pigmented skin, two propigmenting agents, αMSH and Forskolin, were tested in a set of independent reconstructed samples. The culture medium was supplemented with 50 nM α-MSH or 40 μM forskolin, from ED0 to ED18. In both treatment conditions, a visible darkening of the reconstructed skin was noticed with a corresponding decrease in the luminance value (DL>6) and a significant increase in melanin production within the epidermis, as illustrated with αMSH in Figure 5A. In another protocol, 40 μM forskolin was added 7 days earlier, that is, when the cells were seeded onto the dermal equivalent. This resulted in an intense pigmentation—the skin becoming almost black—correlated to a significant higher melanin content within the epidermis and a 26-point reduction in the luminance value (Fig. 5B).
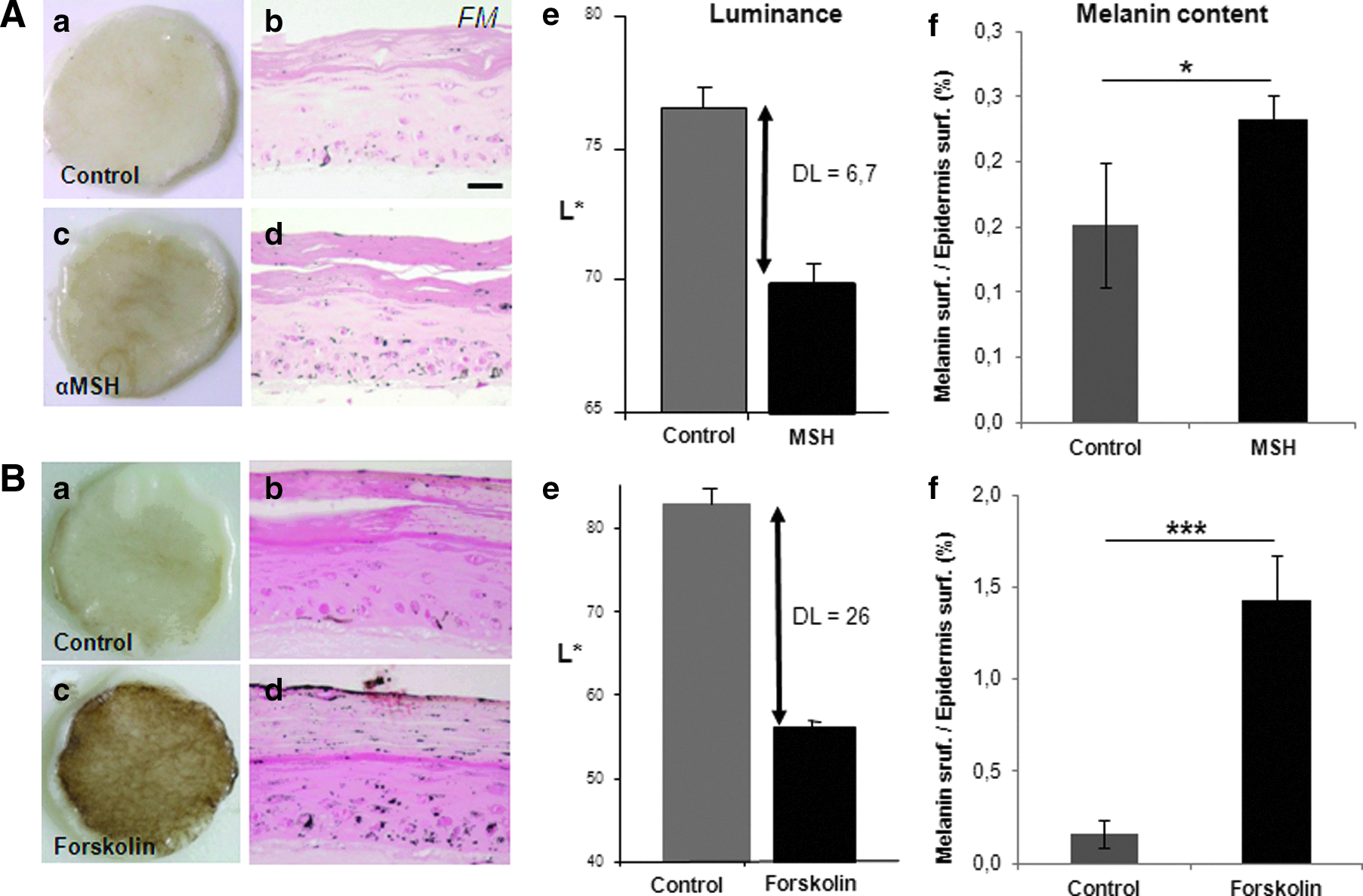
Induction of pigmentation in the reconstructed skin by propigmenting agents. The culture medium was supplemented with
Discussion
To study cutaneous pigmentation in a more physiological context, we have developed a pigmented reconstructed skin that reproduces (1) the 3D architecture of the melanocyte environment; (2) the interactions between melanocytes and their active cellular partners, keratinocytes, and fibroblasts; and (3) the functionality of the pigmentary system. The first experiments were based on our know-how of human skin reconstruction on fibroblast-populated living dermis and production of pigmented reconstructed epidermis onto DED.9,31,38 They allowed us to obtain the integration of melanocytes within the epidermis, using a combined EGF-based keratinocyte medium supplemented with melanocyte factors from the M2 medium (devoid of any phorbol ester). However, although the skin model displayed a good histological quality, melanocytes were mostly bipolar and were found to be in a quiescent status with no melanin production.
One of the major reasons for the observed absence of constitutive pigmentation may be the presence of dermal fibroblasts. The development of a functional pigmented reconstructed epidermis without a living dermis has largely been reported by ourselves and others,9,30,31,42,43 but the introduction of fibroblasts in 3D models seemed more complex with regard to constitutive pigmentation and melanin synthesis. Indeed, fibroblasts have been previously shown to play a regulating role on melanocytes and melanogenesis. Although fibroblasts and the ECM increased pigmentation in 2D co-cultures,13,20 fibroblasts seemed to contribute to keeping melanocytes in an amelanotic state in 3D reconstructed skin models.14,19,21 Indeed, Cario-Andre et al. found that the introduction of fibroblasts, whatever their phototype, decreased pigmentation in epidermis reconstructed on dead DED. 14 In addition, dermal fibroblasts have been shown to exert a negative regulative effect on melanocyte adhesion and survival.19,21
One hypothesis was based on the fact that fibroblasts may inhibit melanin synthesis and reduce the number of melanocytes as a result of their known stimulatory effect on keratinocyte proliferation. 21
Actually, the effects of fibroblasts on pigmentation may be more complex, since the recent work of Choi et al. reported a stimulating effect on pigmentation of one secreted protein, Neuregulin-1, found highly expressed in fibroblasts from phototype VI skin. 15 However, the present experiments have been performed with fibroblasts from Caucasian skin. Introduction of fibroblasts from light to black skin in this full-thickness skin model should be investigated to provide an insight into their differential influence on pigmentation.
To (1) induce melanin synthesis in the skin model containing Caucasian fibroblasts and melanocytes and to (2) obtain a level of pigmentation similar to the in vivo situation, we tested the effects of different growth factors, SCF, ET3, and bFGF, known to act on melanocytes through their specific membrane receptors.44–47 Literature reports multiple effects for these factors, especially for SCF, whose role in the development and the survival of melanocytes is well known, but which has also been described as a melanogen for human melanocytes.48–50
However, the results were disappointing, suggesting that direct stimulation of melanocytes by SCF, ET3, or bFGF was not sufficient to induce pigmentation. It can be argued that other natural melanocyte activators may have been tested such as ET1 or pro-opiomelanocortin-derived peptides such as α-MSH. Although ET1 is thought to enhance melanocyte differentiation more than ET3 (though they share the same membrane receptor), its effect as a potent downregulator of E-cadherin, a key protein for melanocyte maintenance at the basal location, 51 possibly via the upregulation of MCAM, 52 could negatively affect melanocyte integration in reconstructed skin. Regarding the addition of αMSH in the basal culture medium, we have chosen to use it for functionality studies only, to avoid a potential saturation of the α-MSH/MC1R pathway, which is largely involved in the signaling of UV-induced pigmentation.
Based on the well-documented involvement of keratinocytes in pigmentation via the release of numerous paracrine factors and the direct connections taking place between keratinocytes and melanocytes,2,3,10 we tried to overcome the problem of lack of pigmentation through an original approach, that is, the modulation of keratinocyte homeostasis.
Indeed, in a standard EGF-based medium, keratinocytes are highly proliferative, leading to a rapid epidermal turnover. We tried to improve keratinocyte differentiation and turnover, assuming that this could promote melanin production. Previous studies showed that the replacement of EGF by KGF in the culture medium may be a solution.53,54 KGF, a human epithelial growth factor, member of the family of FGFs (FGF-7), has been shown to be a mitogen as efficient as EGF in conventional low-calcium keratinocyte cultures and, in contrast to EGF, to be able to stimulate the expression of early and late markers of keratinocyte differentiation (keratin 1 and filaggrin). 54 In addition, Gibbs et al. showed that the replacement of EGF by KGF for the reconstruction of epidermis on a dead DED normalized (1) the number of living cell layers (6–8 versus 14–16), (2) the Ki67 proliferation index (15% versus 30%), and (3) the granular expression of transglutaminase 1 and involucrin, and repressed the expression of keratinocyte keratins 6, 16, and 17 as observed in vivo. 53
Concerning K16 expression, it could be detected in both EGF and KGF conditions in our reconstructed skin model (Supplementary Fig. S1), in agreement with previous results obtained in nonpigmented organotypic models containing fibroblasts.55,56 Altogether, these data suggest that the culture conditions as well as the presence of fibroblasts contribute in keeping keratinocytes in a relatively activated status.
However, interestingly, in presence of KGF, but not EGF, normalization of the expression of some differentiation markers of the epidermis such as K10 or loricrin could be observed (Supplementary Fig. S1). At the same time, our data show that the switch from EGF to KGF yielded the expected results in terms of development of constitutive pigmentation with the synthesis of melanin by melanocytes.
Chen et al. have recently shown that cultured melanocytes exposed to KGF have an increased tyrosinase and KGFR expression, thus suggesting that a direct effect of KGF on melanocytes and melanogenesis occurred. 57 However, this experiment was conducted in a monolayer system clearly different from the 3D and tricellular condition we created, which is able to restore physiological expression of the KGF receptor on the keratinocyte membrane only. Indeed, it has been reported that KFG is able to act only on keratinocytes through its specific receptor, FGFR2-IIIb, expressed exclusively in the epithelial cell lineage.58,59 Furthermore, several other arguments support the idea that an indirect action of KGF via keratinocytes could prevail on melanocytes and melanogenesis. KGF has been shown to upregulate melanosome transfer by acting primarily on the recipient keratinocytes. 60 Additionally, KGF has shown to induce expression by keratinocyte of the melanocyte-stimulating factor SCF. 61
Finally, improvement of keratinocyte differentiation with KGF may be involved in the secretion of specific paracrine factors requested for melanogenesis, as illustrated by Grabbe et al. for SCF. 62 The normalization of the differentiation induced by KGF can also contribute to promote melanosome transfer to the keratinocytes, as suggested by Belleudi et al. 63
Characterization of the pigmentary system revealed that melanocytes correctly resided in the basal layer, developed dendrites, contained melanosomes expressing major melanogenic proteins like Pmel-17, tyrosinase, TRP-1, and TRP-2. Furthermore, the master melanocyte transcription factor MITF was detected in the melanocytes. Finally, the transfer of melanosomes containing melanin to neighboring keratinocytes demonstrated the functionality of the EMU with many similarities to the in vivo situation.
The melanocyte/keratinocyte ratios within the basal epidermal layer were estimated to be 1/16 and 1/18, which are in the same range of those found in vivo (1/8–1/21).43,64 The basal ratio does not seem to depend on the seeding NHM/NHK ratio, which in our condition was high (1/1), reflecting the low keratinocyte density used (33,000 NHKs/cm2). This implies that keratinocytes strongly proliferate to form a confluent epidermal monolayer during the immersion phase (7 days). In most of the pigmented models, keratinocytes are seeded at a higher density, but cultured for a shorter immersion phase.14,19,21,30,34,35 The seeding density of melanocytes (33,000 NHMs/cm2) is similar to previously published data.9,21,31,35 Seeding melanocytes with few keratinocytes can give an advantage to the melanocytes in terms of space and time to attach to the support as previously suggested.19,35 However, the most interesting finding is that the final basal melanocyte/keratinocyte ratio equilibrated to a physiological balance as checked at ED7 and ED18. This reinforced previous assumptions that natural homeostasis between both cell types was strongly governed by keratinocytes.65,66
As we obtained melanin synthesis with Caucasian melanocytes, the capacity of the model to integrate different strains of melanocytes from various phenotypes was tested. The results revealed that despite difficulties to culture highly pigmented melanocytes—primary thawed cultures are less prone to growth in the M2 culture medium used—the reconstructed skin model could accept more or less pigmented melanocytes, leading to pigmentation that was similar to the original phenotype. This indicates that such a system allows the study of various constitutive pigmentations. Finally, the stimulation of the pigmentary system by propigmenting modulators α-MSH and forskolin revealed the functionality of the model. In parallel, its potential use to evaluate depigmenting agents was also checked with Rucinol, a known in vitro and in vivo lightening agent (see Supplementary Fig. S3).67,68 The stimulation of melanin synthesis by α-MSH demonstrated the responsiveness of the α-MSH/MC1R pathway, in contrast to negative data previously reported on pigmented epidermis reconstructed on a fibroblast-repopulated DED. 19 The propigmenting effect of forskolin, a direct cAMP inducer bypassing MC1R, was even more impressive in our reconstructed skin model and confirmed the high efficacy of this diterpene derivative to darken the skin as previously demonstrated by the Fisher's group in MC1R-deficient humanized mice. 69 In the present study, we focused on the constitutive pigmentation and the responsiveness of melanocytes to chemical pro- or depigmenting agents. Future experiments will be designed to study the functionally of the pigmented skin model face to UV radiation.
In conclusion, a functional and pigmented human skin model comprising normal epidermal keratinocytes, melanocytes, and dermal fibroblasts was developed and will allow us to study the role of cell–cell and cell–matrix interactions in the control of skin pigmentation. This model will represent a powerful tool for elucidating the role of mesenchymal–epithelial interactions involved in melanocyte homeostasis.
Footnotes
Acknowledgments
We would like to thank Pr. Meenhard Herlyn for fruitful discussion and advices, Xavier Marat for help in supplying the depigmenting reference, and Catherine Olivry for artwork.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.