
Abstract
Methods of three-dimensional (3D) cell culture have made significant progress in recent years due to a better understanding of cell to cell interactions and the cell's interface with their surrounding environment. We hypothesized that a microgravity 3D culture system would improve upon the growth and function of a pancreatic progenitor cell population. We developed a rotating wall vessel bioreactor and established a culture system using a pancreatic cell line. Cells in the bioreactors showed robust proliferation, enhanced transcriptional signaling, and improved translation of pancreatic genes compared with two-dimensional static culture. Cells also gained the ability to respond to glucose stimulation, which was not observed in the control cultures. These findings suggest that a 3D microgravity bioreactor environment mimics the niche of the pancreas yielding a cell source with potential for cell-based therapy in the treatment of diabetes.
Introduction
Many of the initial 3D culture systems used continuous stirring or mechanical agitation as a method for keeping cells in suspension but the mechanical agitation was detrimental to fragile cell populations (i.e., progenitors).10,17–20 The negative consequences of these techniques were the impetus for researchers to develop and examine bioreactor cultures that have features of minimal mechanical agitation (e.g., bubble-free oxygenation and higher viscosity culture media). 21 The most successful work was pioneered by the National Aeronautics Space Administration's (NASA) zero-gravity culture experiments that led to the development of earthbound microgravity bioreactor systems created on a rotating wall vessel (RWV) platform.20,22 Studies of RWVs showed that cells grew three dimensionally and gained the ability to aggregate into tissues that had not been observed with monolayer culture.23,24
Since the conception of the RWV along with its commercialization by Synthecon (Houston, TX) multiple cell types including bone, cardiovascular, liver, neural, and renal, have been tested in simulated zero-gravity conditions.25–29 Bioreactors also offer benefit in the cultivation of cells from pancreatic origin. Frequently used methods to create a 3D environment that augments pancreatic or islet cell cultures include collagen gels, microencapsulation, and creation of tissue-engineered chambers.6,30–32 Pancreatic cell lines from rat exocrine tissue are highly proliferative in a rotating 3D culture and maintain their cell markers and potential for differentiation. 18 Embryonic stem cells cultured in 3D environments can be induced to differentiate to pancreatic progenitor cell (PPC) phenotypes. 33 Porcine endocrine tissues (i.e., aggregates of islet-like structures) also have been cultured in 3D and demonstrate successful growth with a 7.5-fold higher number of insulin-positive cells that have the ability to secrete insulin in response to increased glucose concentrations. 17 Using 3D culture enhances the function of human islets by allowing the formation of channels with external openings that transport nutrients and oxygen into the islets after transplant. These islets are less immunogenic and more effective at creating euglycemia in diabetic animal models than the same populations cultured in monolayer.34,35 Similarly, islets cocultured with unique cell populations in 3D culture devices demonstrate enhanced cellular aggregation and allow the islets to maintain their ability to secrete insulin when challenged with glucose. 36 With this background we created a 3D system to propagate and regulate differentiation of a pancreatic progenitor population. 37
Materials and Methods
Cell sources
The cell line Beta-TC-6 (CRL-11506) was obtained from American Type Tissue Culture (Manassas, VA). These cells are derived from β-cell tumors of transgenic mice with the SV40 T-antigen driven by the insulin gene enhancer-promoter region. They are glucose responsive and secrete insulin and glucagon in levels more significant and stable than other previously studied cell lines.38–44
Cell cultures were maintained in media consisting of Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 15% fetal bovine serum (FBS).
All mice were obtained from The Jackson Laboratory (Bar Harbor, ME), including the C57BL/6 strain (stock # 000664; for our purposes termed Wild Type or WT), and mice with an enhanced EGFP cDNA constitutively expressed using the β-actin promoter (stock # 003291, termed GFP+) on a C57BL/6 background. Animals were maintained on standard rodent chow under a constant 12 h day/night cycle. C57BL/6 and GFP positive mice were used for isolation of a progenitor cell population from murine pancreas, which we have extensively studied. The cells are selected based on expression of the surface marker Sca-1 and are designated Sca-1 pancreatic progenitor cells (Sca-PPCs). 37 Care and use of animals was approved by the Institutional Animal Care and Use Committee at the University of North Carolina (UNC) at Chapel Hill in accordance with the principles and procedures outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Reverse transcription–polymerase chain reaction
Polymerase chain reaction analysis was performed on total RNA extracted from freshly isolated and cultured Beta-TC-6 cells. Islet RNA was used as a control. A total of 2 μg of total RNA was used as a template to create complementary DNA using RETROscript® kit (Ambion, Austin, TX) per the manufacturer's protocols. The following pancreatic genes were studied: Nestin, PDX-1, Pax 4, Pax6, Ngn3, Neuro D, insulin 1, insulin 2, GATA 4, GATA6, Isl1, amylase, and ptf1α. Basic conditions included a 94°C denaturation followed by 35 cycles of denaturation at 94°C, annealing at specified melting points, and elongation at 72°C, all followed by a final elongation at 72°C and a final hold of 4°C.
Western blot
Cells were lysed in buffer containing 50 mM Tris-Cl, pH 7.6, 150 mM NaCl, 1% Nonidet P40 (Roche, Indianapolis, IN), 0.5% deoxycholate, 1 mL protease inhibitor cocktail for every 100 mL (5 μg/mL aprotinin, leupeptin, pepstatin, and soybean trypsin inhibitor), and 1 mL phosphatase inhibitor for every 100 mL. Lysates were purified by centrifugation at 14,000 g for 2 min and stored at −20°C. Protein concentrations were determined by using a standard Bradford assay on a BioTek microplate reader. Total cell proteins (50 μg/lane) were separated by SDS-PAGE, transferred to nitrocellulose membrane, incubated with blocking buffer (5% nonfat dry milk in Tris Buffered Saline with 0.05% Tween 20, pH 7.5), and probed with primary antibodies overnight. Antibodies were directed against Pdx1 (1:5000) and Glut-2 (1:1000) (Milipore, Billerica, MA). After 1 h incubation with appropriate HRP secondary antibodies (Dako, Carpinteria, CA), peroxidase activity was detected by enhanced chemiluminescence. Densitometric signals from western blots were analyzed with NIH-ImageJ software (
Insulin enzyme linked immunoabsorbance assay
Insulin secretion of cultured cells was measured using an enzyme linked immunoabsorbance assay (ELISA). Experiments began by placing cell cultures in buffers without glucose and buffers with high glucose concentrations (0 and 100 mM respectively). Our experiments used Krebs Ringer Bicarbonate Buffer (Zenbio, Research Triangle Park, NC), because the presence of FBS in culture media has the potential to affect readings. Experiments involved collecting supernatants over a 2–4 h period (30 min, 1, 2, and 4 h). All samples were frozen at −20°C. Samples were analyzed for insulin concentration according to manufacturer's protocol for the Mercodia Mouse Insulin ELISA (Alpco Diagnostics, Salem, NH). Assays were performed by the UNC Center for Gastrointestinal Biology and Disease (CGIBD) immunotechnology core facility. Values were expressed in ng/mL/μg protein. Single cell suspensions were difficult to obtain from bioreactors for the purpose of cell counting. To compare and quantify secretion data between dishes and bioreactors protein isolation and quantification was employed. Therefore, in the ELISA data graphs all accounts for the unconventional expression of values are expressed as ng/mL/μg protein.
Microcarrier preparation
Cytodex 3 microcarrier beads were purchased from GE HealthCare (Piscataway, NJ). Beads were rehydrated and prepared according to the manufacturer's protocol. Dry microcarriers were hydrated in Ca2+- and Mg2+- free PBS at 50 mL/g for 3 h at room temperature. They were sterilized by removing supernatant and adding 70% isopropyl alcohol. The carriers were washed twice in this manner and incubated overnight in the 70% ethanol solution. The next day the solution was removed and the beads washed thrice with sterile PBS. Cell lines were mixed with beads immediately after the third PBS wash.
Bioreactor culture
Bioreactor cultures were contained in VueLife™ transparent fluoroethylene propylene (FEP) closed culture bags. (American Fluoroseal Corporation, Gaithersburg, MD). Microcarrier beads were added to bags at a concentration of 0.02 g/bag in 1 mL fresh warmed culture media. β-TC6 cells (catalog number CRL-11506) in standard DMEM with 15% FBS (American Type Tissue Culture, Manassas, VA) were added at a concentration of 1–2×106 cells per bioreactor. Bags were then placed into a rotating culture device set at ∼25 revolutions per minute (rpm) and continuously rotated. The bioreactor is capable of rotation rates of 2–30 rpm. Media was changed in bioreactors twice weekly by stopping rotation, allowing cells and beads to settle, and then removing the media via a 22G syringe through the bag's port. Fresh media was then substituted through the same port via another 22G syringe.
Statistical analysis
Student t-test was performed to assess statistically significant differences in test samples.
Results
Development and culture of a 3D bioreactor system
Bioreactor culture systems were configured and created through an interdepartmental collaboration with Dr. Robert Dennis in the Department of Biomedical Engineering at UNC. The base design was modeled after the RWV bioreactor created by NASA and mimics Synthecon's (Houston, TX) commercially available RWVs (Fig. 1). The device measures 8×8×6 inches and is maintained inside a 5% carbon dioxide infused incubator.

Design of 3D tissue culture bioreactor. The bioreactor consists of a pivoting platform that rotates around a fixed point. A motor behind the platform controls power and revolution speed. 3D, three-dimensional. Color images available online at
Our previously characterized murine PPC is present in small numbers per organ, yielding ∼1–2 million cells per pancreatic excision. 37 For this reason initial 3D culture experiments used the commercially available Beta-cell line, Beta-TC-6 (β-TC6) and ATCC CRL-11506 (Manassas, VA).
Cells were cultured in bioreactor devices with the use of Cytodex 3 microbeads (Fig. 2a) and FEP culture bags (Fig. 2B). Cytodex 3 microbeads are 60–87 μm and have a thin layer of denatured collagen and cross-linked dextran allowing a biologically inert substrate that facilitates cellular attachment. Microcarriers provide a support surface for cell attachment while the bags create a microenvironment where the cells expand during culture (Fig. 2). FEP bags measure 3.95 inches by 3.3 inches and have an internal area of 15.7 cm2 holding a 10 mL volume. FEP bags allow for exchange of oxygen, carbon dioxide, nitrogen, and water across their surface, creating a sterile environment for cell culture. Individualβ-TC6 cells initially attach to Cytodex beads at individual points along the bead surface, eventually forming cellular clusters on the individual beads. Over time, clusters detach from the microbeads and form free-floating suspended aggregates (Fig. 3). These aggregates vary in size from 50 μm up to 200 μm. The 3D cytoarchitecture of these clusters morphologically resembles viable islets (Fig. 4A) and is strikingly different than control experiments with cells cultured on a monolayer dish.

Microcarrier beads and culture bags for bioreactor experimentation. Cytodex 3 microcarrier beads are used as a substrate for cells to adhere in rotating culture
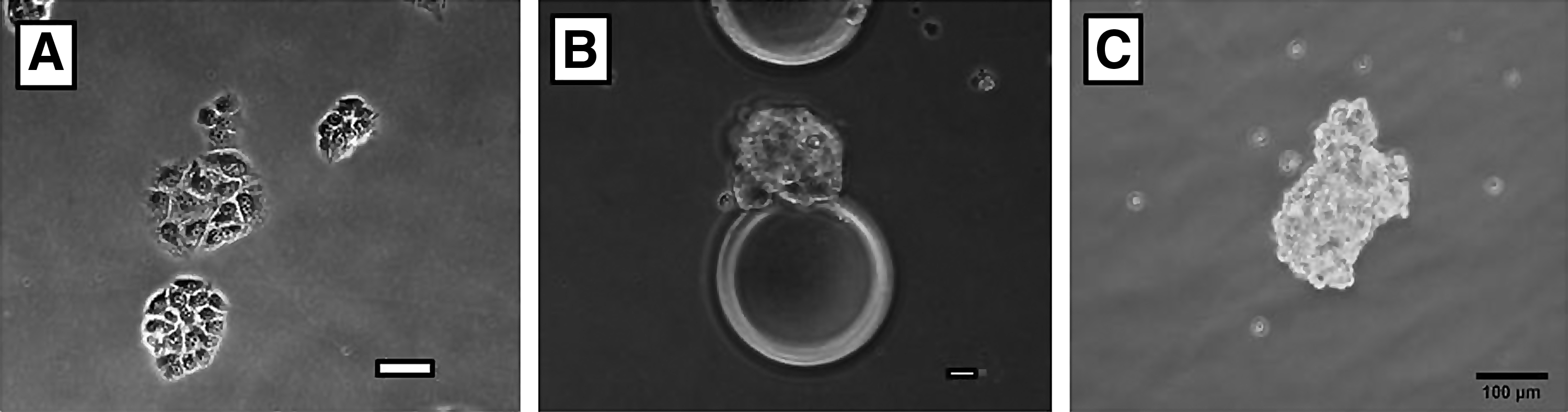
Morphology of β-TC6 cells when grown in monolayer versus bioreactor culture.

Cell clusters formed by β-TC6 cells in bioreactors resemble normal islets in architecture. Islet from physiologically normal mouse pancreas
Transcriptional regulation in bioreactors
Gene transcription was selected based on the association with certain gene signals with pancreatic development, endocrine or exocrine differentiation. At day 5 of culture the semiquantitative analysis from cells in bioreactors and dishes is similar. However, by day 12 there are differences in expression in bioreactor cultures compared with the day 12 dishes. In Figure 5 PDX-1 expression has doubled while Pax 4, Pax 6, GATA 4, GATA 6, Nestin, Hnf4α, and Insulin 2 are tripled. Expression of NeuroD, Nkx6.1, and Isl1 are >3×higher than values obtained for day 12 dish cultures. Interestingly, the developmental marker Ngn3 and exocrine markers, Amylase and Ptf1α, are expressed at day 12 in the bioreactors, but not expressed in 2D static dish cultures at the same time point.
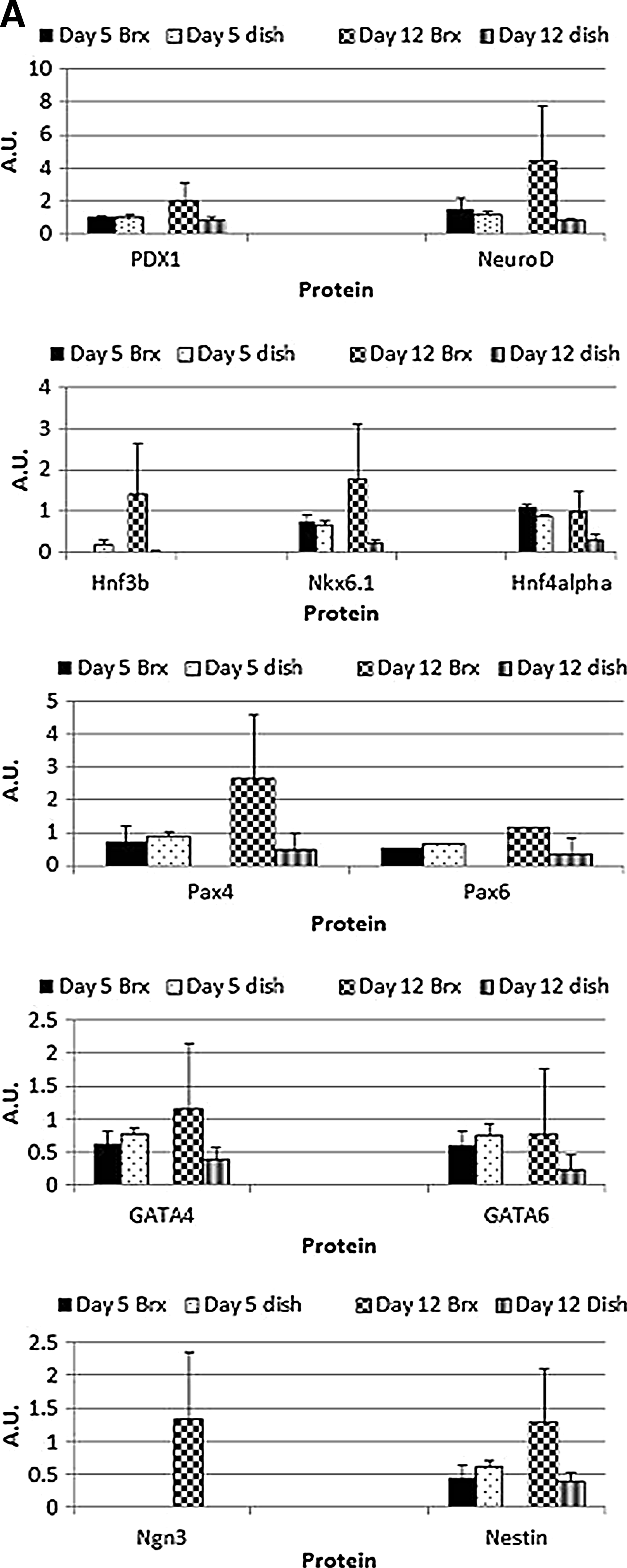
Semiquantitative PCR results for Beta-TC6 cell line in dish and in bioreactor culture. Cells were cultured on a tissue culture dish or in bioreactors for 5 and 12 days and RNA was collected. PCR analysis was performed to independently compare the amount of mRNA present at day 5 and day 12 in bioreactors versus control dishes for genes associated with pancreatic development and endocrine differentiation
Translation of beta-TC-6 cells in bioreactors
Further analysis was performed to determine if the gene transcription of selected pancreatic markers was consistently demonstrated with protein expression. By western blot analysis Glut2 and PDX-1, developmental markers associated with pancreatic endocrine cell development, are increased in the bioreactor cultures compared with the static 2D culture controls (Fig. 6). By day 5 there is a 10-fold increase in PDX-1 in cells in the bioreactors compared with cells on the control dishes, 4.9±7.4 and 0.48±0.34 AU respectively. By day 7, expression is reduced in the control dishes and the bioreactors, but the bioreactors still express twice the amount of PDX 1.6±1.3 AU, compared with dishes, 0.46±0.6 AU. By day 14, PDX expression has increased in control and experimental conditions with bioreactors having >4×the expression, 3.8±4 AU, compared with the 2D static control dishes, 0.9±0.8 AU. A similar pattern exists for Glut2 where day 5 control dishes have 1/5th the expression of cells in the bioreactors, 0.31±0.1 AU compared with 1.5±1 AU, respectively. By day 7, control dishes increase to 0.54±0.2 AU and bioreactors decrease to 0.84±0.4 AU. Finally, by day 14, the control dishes express Glut2 at 0.54±0.2 AU and bioreactors 1.61±1.2 AU. (Fig. 6).
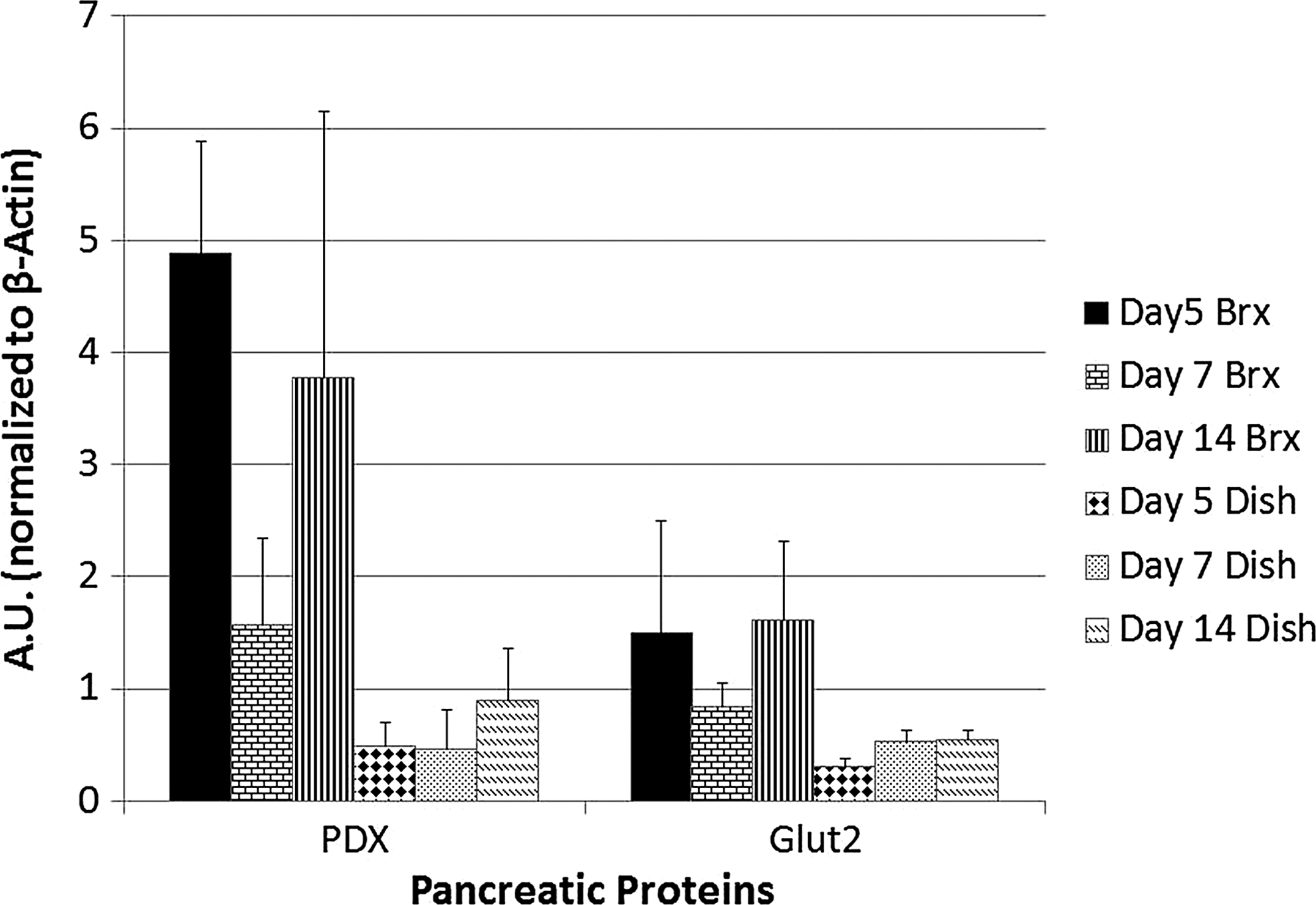
Comparison of PDX-1 and Glut2 expression from β-TC6 cells contained in culture dishes and 3D bioreactors. Protein expression is increased in bioreactor cultures compared with dish cultures. Units are expressed in A.U. normalized for 50 μg of protein. Error bars represent standard deviation.
Improved insulin secretion with bioreactor culture
We subsequently measured the cell line's production of insulin in response to glucose stimulation (Fig. 7). During a 1 h assay there is only a minor response by the β-TC6 cells on control dishes after switching from 0 mM to 100mM glucose concentration, 22.1±3.7 ng/mL produced at 30 min to25.3±3.7 ng/mL at 1 h. Cells cultured in the absence of glucose (0 mM) surprisingly produced higher amounts of insulin 26.9±5.4 ng/mL at 30 min, 45.7±40 ng/mL at 1 h than cells in 100 mM conditions. All 0 mM glucose condition bioreactor cultures resulted in lower readings of insulin production 55.8±52 ng/mL at 30 min, 78.4±44 ng/mL at 1 h compared with their high glucose equivalents. β-TC6 cells cultured in bioreactors at 100 mM glucose concentrations showed a statistically significant increase in the amount of insulin production at the 30 min time point compared with corresponding dish cultures at the same time point (51.6±14.1 to 22.1 2.2±ng/mL).
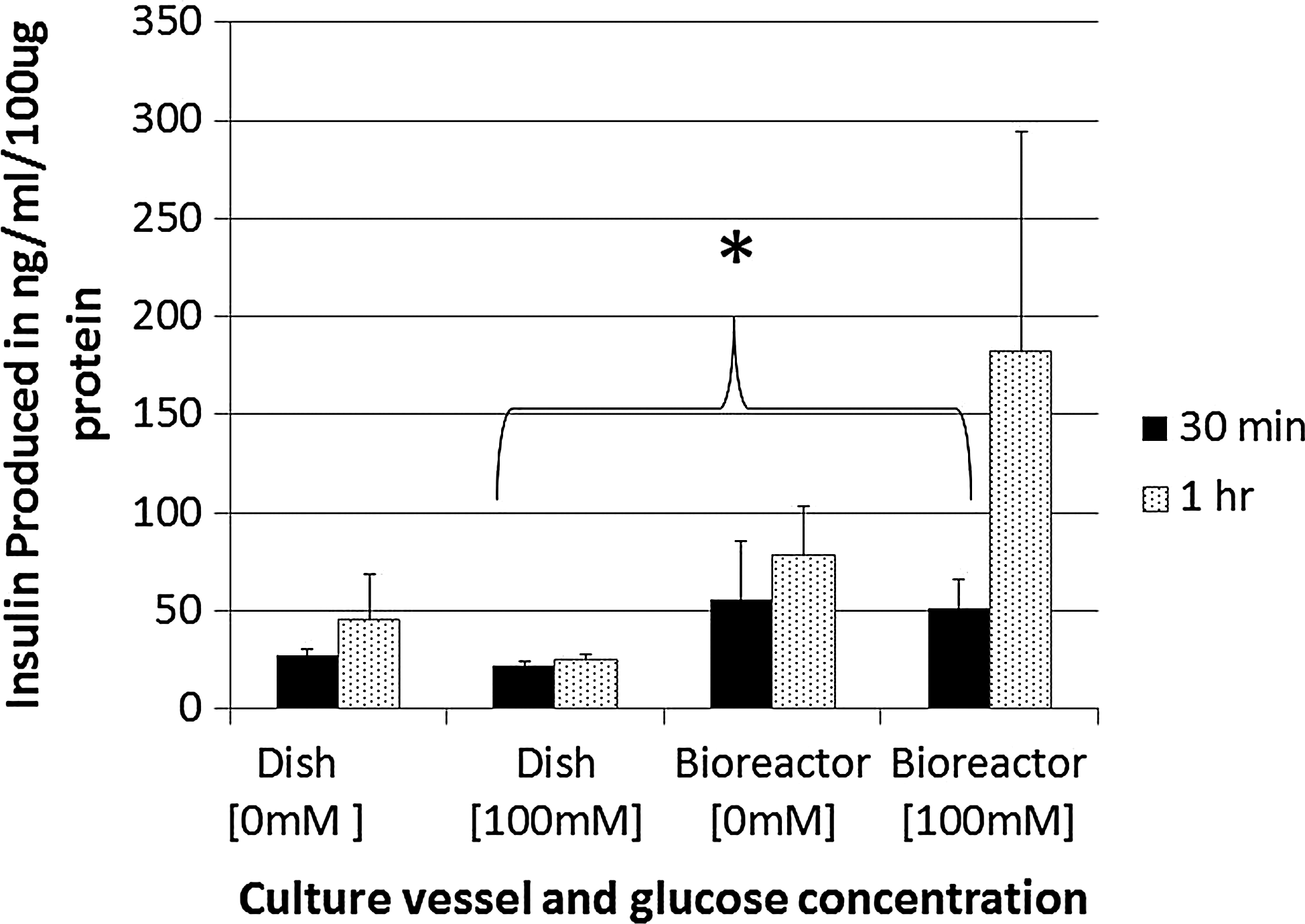
Bioreactors allow for increased secretion of insulin in response to glucose compared with monolayer cultures. Cells cultured in monolayer dishes produce very little insulin in response to increases in glucose concentrations while cells in 3D cultures produce measurable amounts. Cells were initially cultured in a 0 mM glucose environment and subsequently stimulated by changing to a high glucose (100 mM) concentration for 1 h, with samples taken at 30 min and 1 h. Error bars represent standard error. Asterisk indicates statistical significance of 0.047 between dish and bioreactor cultures in high glucose conditions at 30 min.
PPCs in 3D culture
Based on the previous results we designed an experiment to study the effect of the microgravity 3D culture system on a previously isolated and characterized PPC population. 37 Approximately 4 h after loading cells into bioreactor cultures, PPCs attach to microbeads at single contact points. Over the course of 2–3 weeks the cells begin proliferating and eventually cover the entire bead. After the beads are covered, the cells begin to form cellular bridges with adjacent beads thus generating neo-tissue aggregates among multiple beads (data not shown).
After the PPCs proliferate to the point where they cover the microcarrier beads they were removed from the bioreactors (maintained at 5.5 mM glucose) and placed in media containing 55.5 mM glucose to be analyzed for their ability to secrete insulin over a 2 h interval. Over the first 2 h of exposure to the 55.5 mM glucose environment there is a demonstrable increase in insulin production from an average of 2.4 ng/mL at 30 min, 10.9 ng/mL at 1 h, and 18.26 ng/mL at 2 h (Fig. 8A).In a static 2D culture, the PPCs do not demonstrate a spike in insulin production after switching to the higher glucose concentration media. Representative images of PPCs grown in 2D versus 3D are seen in Figure 8B.
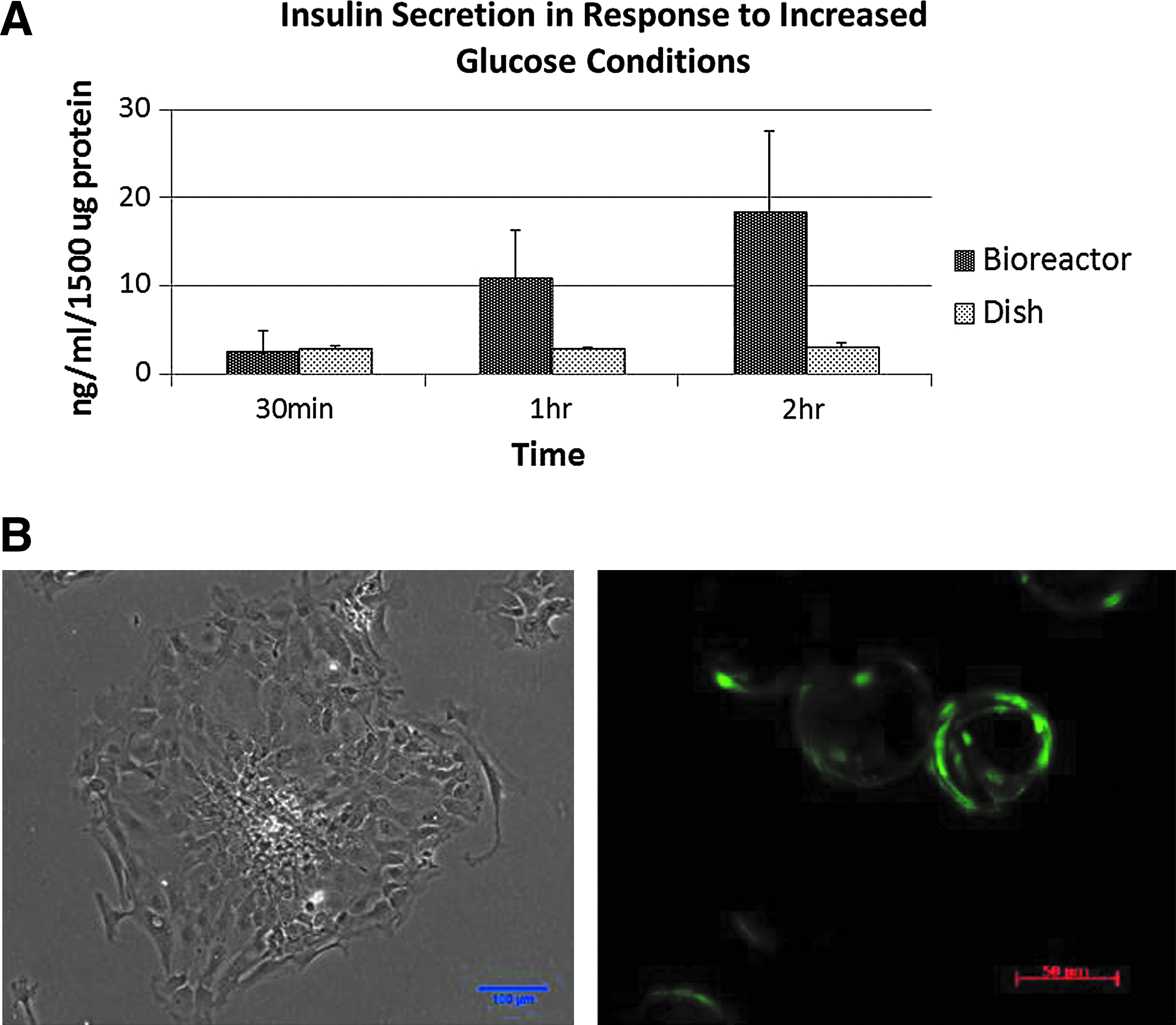
PPCs demonstrate improved response to increased glucose in 3D bioreactor cultures when compared with dish cultures. Insulin secretion was measured after switching the PPC cultures from 5.5 mM glucose to a 55.5 mM glucose-supplemented media. Error bars represent standard error
Discussion
Three-dimensional cell cultures offer an environment that is superior to that provided in monolayer conditions by creating a highly complex environment without physical constraints. The 3D systems allow cell populations to interface with one another and foster complex cellular expansion similar to that seen during in vivo development. Compared with static dish cultures the 3D systems allow for more robust growth of cell populations. Cells have expanded surface area to differentiate and divide without the constraints of a rigid tissue culture dish. The bioreactor culture allows cell division to occur through a series of naturally occurring events that rely on spatial and temporal organization and mechanical cues, communication between cells and their matrix, and communication between individual cells. 45 We modeled our bioreactor design after an RWV that maintains cells in a state of simulated zero gravity with very low turbulence and shear stress to the cells. 46 Our model also allows for the integration of FEP culture bags. These bags are highly permeable to oxygen and carbon dioxide allowing for gas exchange across their surface, eliminating the need for infusion of biologic gases by physical methods.
Our bioreactor design provides a culture environment that allows β-TC6 cells of pancreatic origin to aggregate and form “islet-like” clusters. These clusters are not observed when the same cell line is grown in a monolayer rigid environment where nonuniform shapes and cell colonies develop. Our findings are similar to those observed with another pancreatic cell line, MIN6, which forms insulin secreting islet-like configurations in 3D culture. 47
The impact of the bioreactor cultures was assessed by studying the expression of transcription factors associated with pancreatic development and/or differentiated pancreatic cells. Through the first 5 days of culture in our bioreactor design transcriptional regulation is similar to standard dish culture. Early pancreatic developmental markers PAX4, PAX 6, GATA 4, GATA 6, Nestin, and Hnf4α are tripled in day 12 bioreactor samples compared with their controls and the developmental marker PDX-1 is doubled in bioreactor cultures. Similar increases are also seen with mature pancreatic cell markers Insulin 2 and Isl1, the developmental markers Nestin and Ngn3, and exocrine markers Amylase and Ptf1α, which were not detectable in controls at day 5 or 12. Transcription of pancreatic markers was consistently upregulated over time in bioreactor cultures, compared to monolayer dish cultures. While no statistical significance was observed, a trend toward higher readings in bioreactors compared with dishes after 12 days of culture was clearly detectable (Fig. 5).
Based on our initial experiments we evaluated cellular differentiation in the bioreactors. Both bioreactor cultures and dish cultures were assayed for cellular production of PDX-1 and Glut-2 during a 2-week culture. Bioreactor protein expression was consistently higher across all time points compared with corresponding dish conditions, with PDX-1 expression increased 10-fold in bioreactors compared with control dishes at day 5 of culture. Glut 2 is similarly increased in bioreactors.
A physiologic response of insulin secretion in response to an increase in glucose conditions is an essential feature of pancreatic β-cell function. When cultured in monolayer dishes β-TC6 cells do not demonstrate the ability to acutely respond to glucose stimulation and these cells demonstrate unregulated and autonomous secretion of insulin, 15.6 ng/mL more insulin than when the same cells are cultured in a high glucose concentration. In contrast, when the cell line was cultured in our bioreactor 3D design the cells acutely responded in a physiologic manner with cells in high glucose conditions secreting a statistically significant 51.6 ng/mL of insulin in bioreactors compared with 26.9 ng/mL in dishes. Thus, the bioreactor culture demonstrates gain of physiologic function with respect to insulin secretion. It is unclear what mechanisms are associated with this enhanced physiologic function in bioreactors but it has been hypothesized that 3D culture allows for formation of nutritional channels and openings in cellular aggregates that allow exchange of nutrients and transport of cellular secretions. 34
By simply removing PPCs from a monolayer culture and placing them into a 3D environment there was a demonstrated improvement in physiologic function. It is possible that differences in physical constraints and extracellular matrix composition are major factors contributing to this enhanced function. Future experiments will aim to address the optimal scaffold and matrix structure upon which to culture cells in a novel bioreactor environment. Future experiments will also aim to assess whether PDX-1 is capable of activation in 3D culture and if it is being translocated to cellular nuclei thus imparting enhanced β-cell activity. Our previous work demonstrates that PPC colonies are not solely composed of β-like cells. The colonies contain cells similar to the other cell types that comprise the normal pancreatic islet (alpha, delta, etc.). 37 It is possible that culture our 3D environment allows the cells to proliferate and cluster in a manner that favors insulin secretion in the bioreactors compared with the colonies on control dishes. Future experiments will investigate the topology of the cell clusters addressing if the cells arrange themselves in a fashion similar to normal islets, (e.g., beta cells forming the core of islets and other cell types surrounding the beta cell core).Taking away the physical constraints associated with adherence on a dish allows the cells to develop and function in a more natural physiologic manner than is seen in 2D culture. It is possible that a 3D environment allows for improved cell–cell interactions and communication thus allowing the cells to receive the signals needed to differentiate toward an islet-like phenotype.
This study shows that a 3D microgravity culture environment positively affects proliferation and function of a pancreatic cell population. Further, we show that the 3D environment is beneficial to this PPC population allowing for long-term culture without passage and the ability to differentiate and secrete insulin in response to a glucose challenge. We have created a simple and scalable system in which the pancreatic islet-like morphologic assembly is possible with long-term survival and enhanced physiologic function. It is feasible that future treatments aimed to treat diabetes mellitus could involve transplantation of β-like cells expanded in 3D culture.
Footnotes
Acknowledgment
We would like to thank Robert Dennis, Ph.D. for designing and fabricating all bioreactor devices.
Disclosure Statement
No competing financial interests exist.