
Abstract
Liver transplantation, utilized routinely for end-stage liver disease, has been constrained by the paucity of organ donors, and is being complemented by alternative strategies such as liver cell transplantation. One of the most promising forms of liver cell transplantation is hepatic stem cell therapies, as the number of human hepatic stem cells (hHpSCs) and other early hepatic progenitor cells (HPCs) are sufficient to provide treatment for multiple patients from a single liver source. In the present study, human adult livers were exposed to cold ischemia and then processed after <24 or 48 h. Cells positive for epithelial cell adhesion molecule (EpCAM), a marker on early lineage stage HPCs, were immunoselected and counted. Approximately 100,000 EpCAM+ cells/gram of tissue was obtained from surgical resection of livers subjected to cold ischemia up to 24 h and comparable numbers, albeit somewhat lower, were obtained from those exposed to 48 h of cold ischemia. The yields are similar to those reported from livers with minimal exposure to ischemia. When cultured on plastic dishes and in Kubota's Medium, a serum-free medium designed for early lineage stage HPCs, colonies of rapidly expanding cells formed. They were confirmed to be probable hHpSCs by their ability to survive and expand on plastic and in Kubota's Medium for months, by co-expression of EpCAM and neural cell adhesion molecule, minimal if any albumin expression, with EpCAM found throughout the cells, and no expression of alpha-fetoprotein. The yields of viable EpCAM+ cells were surprisingly large, and the numbers from a single donor liver are sufficient to treat approximately 50–100 patients given the numbers of EpCAM+ cells currently used in hepatic stem cell therapies. Thus, cold ischemic livers for up to 48 h are a new source of cells that might be used for liver cell therapies.
Introduction
Human hepatic stem cells (hHpSCs) and several forms of hepatic progenitor cells (HPCs) have been found to be positive for epithelial cell adhesion molecule (EpCAM) and are evident in livers of all donor ages.16,17 They have far greater expansion and are more readily cryopreserved than are mature liver cells. 18 EpCAM+ cells from donor livers of any age could represent a new source of cells for cell therapies and cellular models for physio-pathology and research in the pharmaco-toxicology area.19,20 Hepatic progenitors were described for the first time by Farber in 1956, 21 and since then, numerous different origins were described such as intra- or extra-hepatic cells.13,22,23 Of great interest is the fact that one can isolate EpCAM+ cells from adult livers with minimal ethical issues.
The isolation, the characterization, the culture conditions, and the differentiation of human hHpSCs and human hepatoblasts (hHBs) and committed progenitors have been established by Reid and associates.16,17,24–26 They have developed an innovative protocol of stem cell and progenitor cell isolation from livers of any donor age.16,17,25 The protocol makes use of immunoselection for several cell adhesion molecules that are lineage-stage specific and which include EpCAM, neural cell adhesion molecule (NCAM), and intercellular cell adhesion molecule (ICAM). EpCAM is a homotypic calcium-independent cell adhesion molecule that has been identified in most normal epithelial cells and gastrointestinal epithelial carcinomas (colon, rectum, liver, and pancreas carcinomas). 27 This marker is expressed in hHpSCs, hHBs, and variably in committed hepatic progenitors but is not expressed by adult parenchymal cells.25,27,28 Distinguishable phenotype traits among the stem/progenitor cell subpopulations are that hHpSCs, but not hHBs or committed progenitors, express NCAM. By contrast, ICAM is expressed by the hHBs and the committed progenitors but not by the hHpSCs.16,25 Reid et al. have previously reported that the percentage of [EpCAM+ NCAM+] cells, the hHpSCs, is stably present throughout life and with an average of 0.5%–1.5% in normal livers; it increases significantly in ischemic livers (up to 2%–3% in livers subjected to cold ischemia), as the mature parenchymal cells but not the hHpSCs and hHBs are selectively lost due to ischemia. This translates to ∼109 cells for an average adult liver. By contrast, the [EpCAM+ ICAM+] cells, the hHBs and committed progenitors, are age dependent, ranging from more than 80%–90% of the parenchyma in fetal livers; approximately 50% in neonatal livers; and only ∼0.001% to 0.01% in adult livers. 16 Thus, in fetal livers, isolation of EpCAM+ cells yields cells that are predominantly hHBs, whereas EpCAM+ cells from adult livers yield primarily the hHpSCs with a tiny subpopulation of hHBs and committed progenitors. The EpCAM+ cells isolated from both fetal and adult livers were able to expand and to differentiate into both hepatic and biliary lineages in vitro and in vivo.16,29,30 When they were injected into an NOD/SCID mouse model treated with CCl4, a significant engraftment was observed. 16
Clinical trials of liver cell therapies have been done comparing CD34+ cells from bone marrow, MSCs, and EpCAM+ cells from fetal livers.20,31 Publications from treatments of the first 100 patients were recently issued. Although all stem cell populations resulted in alleviation of symptoms of liver failure, the most potent proved to be EpCAM+ cells. 20 Thus, there is considerable interest in establishing protocols for banking of EpCAM+ cells for use in forms of liver cell therapies.
Interestingly, EpCAM+ cells from adult livers and ES cells are more resistant to hypoxia and ischemia than are mature cells,32,33 perhaps in link with the relative hypoxic environment of their “niche.” Ezashi et al. showed that human embryonic cells reduced their differentiation and maintained a pluripotent state when cultured in hypoxic conditions. 34 More recently, Jozefczuk et al.32,33 have used the resistance property of HPCs to selectively isolate them from adult human liver submitted to a prolonged cold ischemic period (for up to 7 days) before cell isolation. As they obtained HPCs with morphological and antigenic characteristics similar to HPCs from other groups, they concluded to a high tolerance of HPCs to ischemic stress. Under these experimental conditions, no mature hepatocytes were detected after 48 h of ischemia. By contrast, HPCs appeared after 2 days of culture-forming dense colonies containing well-defined cells with a low cytoplasm-to-nucleus ratio. The authors suggested that ischemia could help the emergence of stem and progenitor cells and reported that HPCs isolation was possible for up to 7 days of cold ischemia, though their data suffered due to the lack of a statistical analysis. In addition, strengthening the special attention that should be given to the ischemia resistance property of HPCs with regard to their isolation are the well-known examples of influence of the oxygenation ratio on the replication and differentiation of hematopoietic stem cells 35 or MSCs. 36 It has been suggested that liver samples from nonheart beating donors could then be considered a source for HPCs. 16
The first objective of the present study was to develop a protocol of HPCs isolation from adult human livers, by adapting the well-described protocol by the group of Reid with livers of all donor ages from fetal to adult livers. 16 The second aim was to analyze the resistance of HPCs to ischemic stress. For this purpose, we performed cell isolation from adult human livers by splitting them into two pieces of equivalent weight. The isolation procedure was applied within 24 h after the beginning of cold ischemia (T0–24) or after 48 h of cold ischemia (T48). The number of EpCAM+ cells isolated from both pieces was compared and subjected to statistical analyses. In addition, cells were characterized at the mRNA and protein level in order to confirm their status as stem cells and/or progenitors.
Materials and Methods
Adult human livers
Human liver samples (n=27 including Caroli disease, cholangiocarcinoma, cirrhosis, colorectal and non colorectal metastasis, focal nodular hyperplasia, and hepatic adenoma) were obtained from therapeutic partial hepatectomies. All experimental procedures were performed according to French laws and regulations after approval by the National Ethics Committee. Informed consent was obtained from the patients for the use of liver tissue for research purposes. Samples were immediately stored in IgL-1® (Institut Georges Lopez) or Celsior® (Genzyme) solutions at 4°C after hepatectomy defining the initial point for the cold ischemia period. From each resection, two pieces weighting 10.5±0.7 g were utilized. Human HPC isolation was performed on one of these pieces within a 24 h period after the beginning of cold ischemia (referred as T0–24) and on a second piece after 48 h of cold ischemia (referred as T48), the sample being stored in the same solution at 4°C without another treatment. Both pieces were then processed using the same procedure.
Human HPC isolation
The procedure employed to isolate HPCs was based on Reid's protocol, described for both fetal and adult livers. 26 The liver piece was first physically dissociated and then digested using a buffer consisting of 150 U/mL collagenase supplemented with 0.03% desoxyribonuclease (w/v) (Sigma-Aldrich) in RPMI 1640 (Life Technologies, Gibco), 0.1% (w/v) bovine serum albumin fatty acid free, 30 nM selenium (Sigma-Aldrich), and antibiotic-antimycotic mixture (Life Technologies, Gibco). After incubation at 37°C for 20 min, the suspension was filtered through 1000 and 500 μm stainless steel meshes (Fischer Scientific), and finally centrifuged at 200 g for 5 min at 4°C. The resulting pellet was incubated in a lysis buffer (v/v) (Red Blood Cell Lysis Buffer; StemCell technologies) for 5 min at room temperature and centrifugated at 200 g for 5 min to remove hematopoietic cells. The suspension was washed in RPMI 1640 medium supplemented with 0.1% (w/v) bovine serum albumin fatty acid free, 30 nM selenium, and antibiotic-antimycotic mixture and filtered through 100 μm mesh. The cell count was determined by using an automated cell counter system (Scepter® Millipore).
Immunoselection for EpCAM+ cells
Cell sorting for EpCAM+ cells was performed using EasySep® Human EpCAM Positive Selection Kit according to the manufacturer's instructions (StemCell technologies). Briefly, a cell suspension at 1.108 cells/mL that had been prepared in HBSS buffer supplemented with 0.1 mg/mL DNase I and 2% of fetal bovine serum (FBS) was incubated at 4°C with EpCAM selection cocktail for 20 min at the concentration of 100 μL per mL of total suspension. Magnetic nanoparticles were added at 50 μL/mL cells, and the suspension was incubated for 15 min at 4°C. Finally, the suspension was placed into the EasySep® Magnet, to recover labeled and positively enriched cells. The purity degree of the labeled cells was assessed by staining cells with a monoclonal antibody 5E11-FITC (StemCell technologies). A cell count of EpCAM+ cells was performed by using Scepter®system.
Cell culture
After immunoselection, EpCAM+ cells were seeded (105 cells/well) onto six-well plastic plates in Kubota's medium. 37 This medium consisted of RPMI 1640 supplemented with 0.1% bovine serum albumin fatty acid free (w/v), 0.054% niacinamide (w/v), 0.0005% insulin (w/v), transferrin/Fe (10 μg/mL), 30 nM selenium, free fatty acid mixture, 10−8M hydrocortisone, 50 μM β-mercaptoethanol, 2 mM L-glutamine, and antibiotic-antimycotic mixture. During the first 24 h of culture, 10% FBS was added to this medium. After that, the serum-free medium was changed every 2–3 days. The cultures were maintained at 37°C in a humidified incubator in a mixture of 95% air and 5% CO2. Cell colonies were observed every day for 4–6 days, were maintained in culture, and were regularly observed during 3 months.
Confocal microscopy
EpCAM+ and HepG2 cells were cultured in eight-well glass culture slides (BD Falcon, Becton Dickinson) in Kubota's medium. After 1 week of culture for HepG2 cells and 30 days of culture for HPCs, cells were fixed using 4% paraformaldehyde (Sigma-Aldrich) for 10 min and permeabilized using 0.1% Triton×100 for 4 min. Blocking of the slides was performed with goat serum (10%) for 30 min at room temperature, followed by incubation with primary antibodies (a mouse primary antibody targeted against EpCAM, dilution 1:750; MicromMicrotech/Francheville/France, a rabbit primary antibody targeted against NCAM, dilution 1:100; Epitomics, and a mouse primary antibody targeted against ICAM-1, dilution 1:180, Invitrogen) in phosphate-buffered saline (PBS) containing 1% bovine serum albumin for 40 min at room temperature. In parallel, negative controls (no addition of a primary antibody) were performed for the detection of nonspecific binding of the secondary antibody. After washing the slides, Alexa Fluor 488 goat anti-mouse antibody (diluted at 1:500; Molecular Probes/Life Technologies), Alexa Fluor 633 goat anti-mouse antibody (diluted at 1:500; Molecular Probes/Life Technologies), and TRITC goat anti-rabbit antibody (diluted at 1:1000; Rockland) were applied in the slides for 40 min at room temperature in the dark. Nuclei of the cells were stained with DAPI (Sigma-Aldrich). Slides were mounted in Fluorescence Mounting Medium (Dako), and stored at 4°C in the dark until analysis. Immunofluorescence was observed using an Olympus FluoView FV1000 Confocal Laser ScanningBiological Microscope and analyzed with Olympus FV-Viewer Software (Olympus).
Reverse-transcriptase polymerase chain reaction
Total RNA was extracted by using Trizol reagent according to the manufacturer's instructions (Life Technologies). cDNA was synthesized from 5 ng of total RNA using the iScript cDNA Synthesis kit (Bio-Rad) at 42°C for 30 min. The primer sequences (Table 1) were designed using Beacon Designer® software or have been previously described. 30 Quantitative reverse-transcriptase polymerase chain reaction (RT-PCR) was performed from 5 μL of diluted RT reaction (1/1000) using iQ-SYBR Green Supermix (Bio-Rad) in duplicate. The PCR parameters were as follows: a denaturation step at 95°C for 3 min, an amplification step for 40 cycles of PCR (denaturation at 95°C for 10 s and annealing extension at 58°C for 60 s). Amplification specificity was evaluated by determining the product melting curve and was obtained after 80 cycles of 10 s at 55°C. The relative expression of mRNA in HPCs was estimated by the delta-delta method 38 with β-actin and GAPDH as the reference genes. Moreover, mRNA expressions in these cells are expressed as a ratio to the mRNA expression levels measured in a pool of mature human hepatocytes (n=5), isolated as previously described by us.39,40
Flow cytometry
Cells were incubated for 30 min at 4°C with a fluorescent-labeled monoclonal antibody: EpCAM-FITC (clone EBA-1; BD Biosciences) and CD54-APC (clone HA-58; BD Biosciences) according to the manufacturer's instructions. Cells were fixed with 4% paraformaldehyde in PBS for 15 min at room temperature in the dark. An analysis was performed using an FACSCalibur flow cytometer (FC500; Beckman coulter). Nonspecific binding with isotype-matched control antibodies (BD Biosciences) was used to establish gating.
Statistical analysis
A statistical analysis was performed to compare the number of EpCAM+ cells at T0–24 and T48 that corresponded to a quantitative variable in paired series. Since these data did not follow a normal distribution and there was no equality of variances between the two groups, we used a nonparametric test, that is, the Wilcoxon test for paired data. The level of statistical significance was set at 5%.
Results
Influence of cold ischemia on the number of EpCAM+ cells (T0–24 vs. T48 groups)
Purity of the enriched EpCAM+ cells was typically around 60% by flow cytometry, recovery was 1% when expressed as a median, and viability of the enriched EpCAM+ cells always exceeded 90% (n=27 human livers processed). The yields of EpCAM+ cells from 24 h versus 48 h (T0–24 vs. T48) was assessed and noted, expressed as a median, in Figure 1a, individual data being detailed in Figure 1b. In the T0–24 group, the median was 98,150 (ranging from 14,450 to 591,724) cells per gram liver, whereas after 48 h of ischemia (T48), the median was 79,300 (ranging from 13,231 to 915,588) cells per gram of liver. This corresponds to a statistically significant (nonparametric Wilcoxon's test, p=0.014) lower (−20%) cell yield after the period of 48 h under cold ischemia conditions. Of particular interest was the finding that the morphology of EpCAM+ cell colonies from the T0–24 group was identical to the colonies from the T48 group (data not shown).

Influence of cold ischemia on the number of viable EpCAM+ cells. EpCAM+ cell number obtained per gram of liver in the two groups T0–24 and T48 and expressed as a median of n=27 samples
Growth characteristics of EpCAM+ cells isolated from human liver samples at T0–24
The EpCAM+ cells obtained when using human liver samples within a 24 h period after the beginning of cold ischemia (T0–24) were then seeded on plastic support, and their morphology was observed over time (Fig. 2). After 48 h of culture, small colonies of cells appeared (Fig. 2a). The first colonies typically contained were three to four layers thick and with the cells being uniform. These cells were small, with an average diameter of 7 to 9 μm, with a round nucleus occupying the majority of the cell. Up to four nucleoli were observed. Each cell was well defined. After 5 days of culture (Fig. 2b), colonies of cells had expanded, and cells became polygonal and with more abundant cytoplasm. These cells contained two or more nucleoli and never reached confluence. The average doubling time was between 24 and 48 h in culture. The colonies grew over time (Fig. 2c) but after 3 weeks of culture (Fig. 2d), the structure of the cell colony partially degenerated and totally disappeared after 3 months of culture. The type of cell colonies and the cell features were equivalent to the features obtained with colonies of progenitors adult human livers.16,29,30
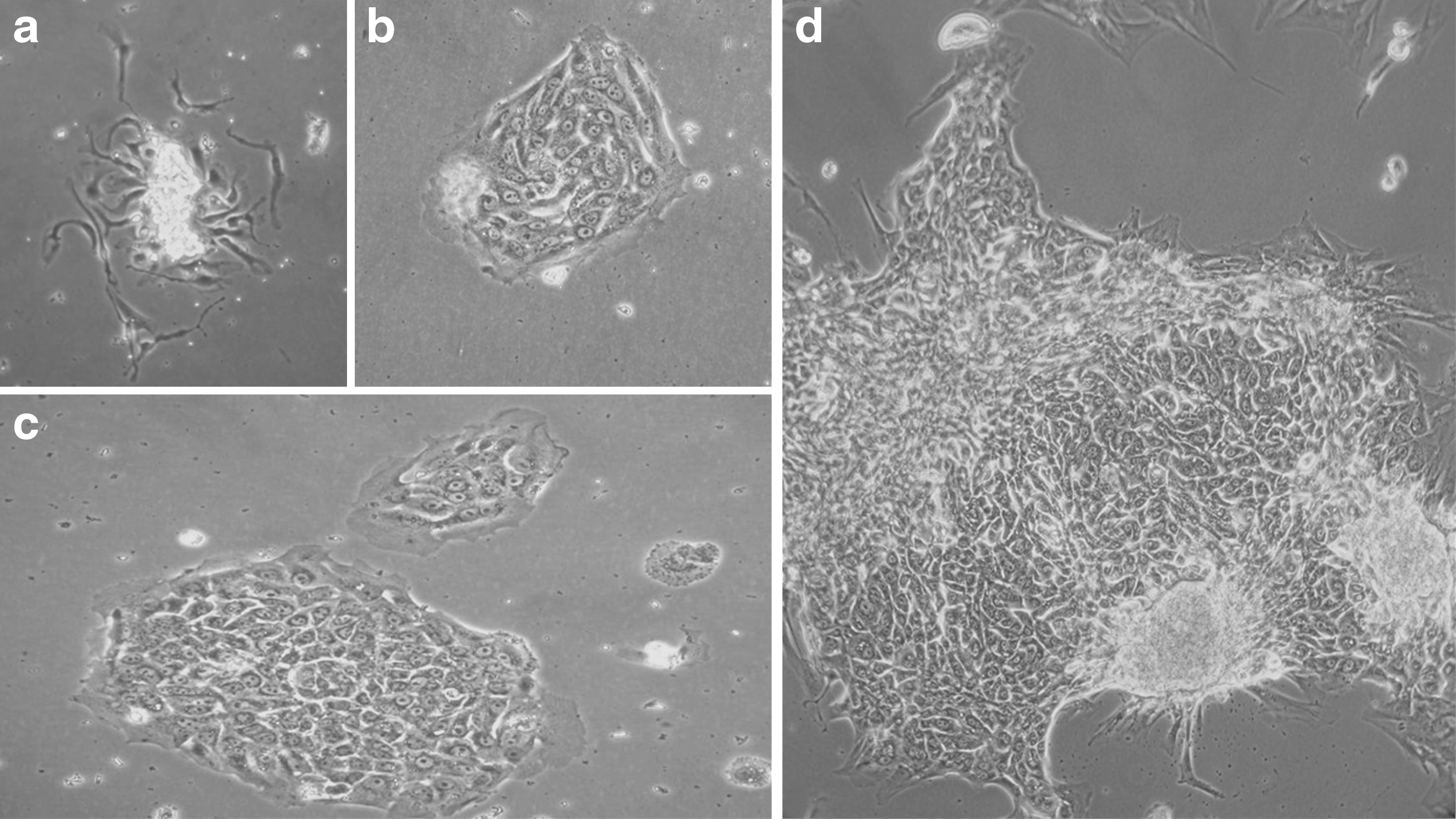
Morphology of EpCAM+ cells freshly isolated from human liver samples (T0–24 group)observed by phase-contrast microscopy. Monitoring over time of cells isolated from human liver sample at T0–24 and cultured on plastic support. The first adherent cells appeared 2 days after cell isolation
Characterization of the cells by RT-PCR assays for the mRNA and by confocal microscopy for protein expression levels in EpCAM+ cells from ischemic livers
In order to assess the expression of HPCs-related markers, we evaluated EpCAM and NCAM mRNA expression by using real-time PCR analysis in T0–24 and T48 groups (Fig. 3). The mRNA expression of both markers in these cells was compared with freshly isolated, adult human hepatocytes. For this, a pool of adult hepatocytes (n=5) was processed, and their mRNA expression was set at 1. In the EpCAM+-labeled cells obtained after cell sorting, EpCAM mRNA was highly expressed and ranged between 6 and up to 65-fold changes depending on the donor and the group, as compared with adult hepatocytes. By contrast, NCAM mRNA was moderately expressed (ranging from 1- to 19-fold changes) as compared with mature hepatocytes. Albumin mRNA expression was lower than in adult hepatocytes, the corresponding fold changes ranging from −8 to −74, depending on the donor. Enriched EpCAM+ cells expressed CK18 and CK19 mRNA (<3.3 and <3.9-fold, respectively, as compared with human hepatocytes). No expression of alpha-fetoprotein was detected in the EpCAM+-labeled cells. Finally, a very subtle expression of ICAM-1 at the mRNA level was detected (data not shown) that corresponds to the low percent (approximately 1%) of EpCAM+ cells being ICAM-1+ on flow cytometry analysis. For all these markers, no difference between T0–24 andT48 groups was obtained.
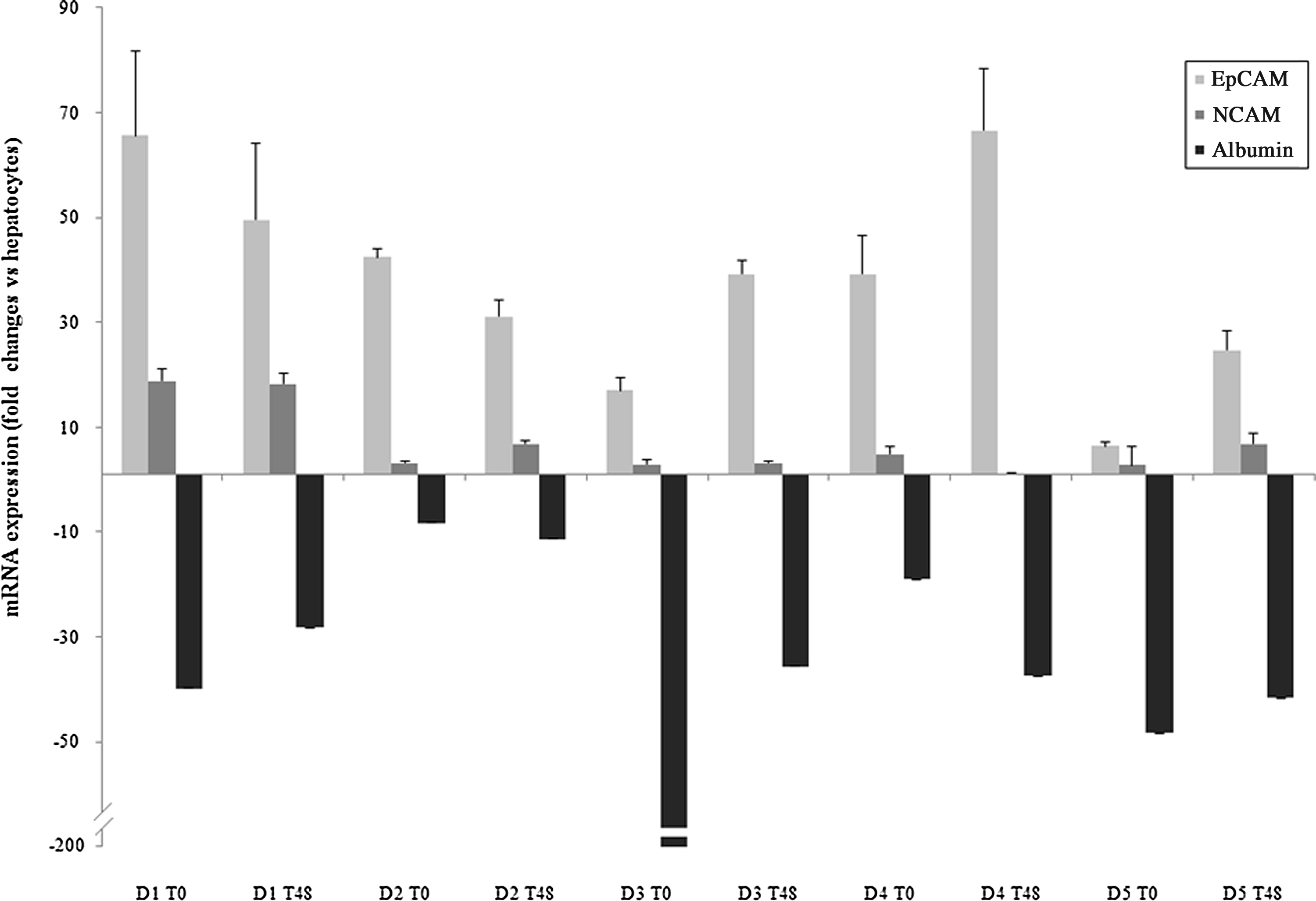
mRNA expression of epithelial cell adhesion molecule (EpCAM) and neural cell adhesion molecule (NCAM) in cultures of hepatic progenitor cells from ischemic adult livers utilizing reverse-transcriptase polymerase chain reaction (RT-PCR). The mRNA expression of EpCAM and NCAM was evaluated in cultures derived from 5 donors. Albumin mRNA expression was also evaluated to confirm the immature stage lineage of HPCs. Results are expressed as fold change (mean±SD) as compared with adult human hepatocytes (pool of 5 donors). Analyses were performed in duplicate with consistent results, and the negative scale was truncated to avoid dwarfing values.
The cell colonies that we obtained from EpCAM+-labeled cells after cell sorting were immunostained using EpCAM and NCAM expression markers and observed by confocal microscopy. HepG2 cell line was first used to develop optimized immunostaining protocols and to serve as a positive control, as this cell line expresses both markers studied (Fig. 4a–d). Cells isolated from the T0–24 group were immunostained after 30 days of culture on glass slides; no degeneration of the cell colonies was noticed in this type of support (eight-well culture plate) in contrast to six-well plastic culture dishes (Fig. 4). A marked staining was observed for EpCAM expression, whereas NCAM was less expressed in HPCs and no staining appeared for ICAM-1. Immunoreactivity for EpCAM was observed in all cells, both in their cytoplasm (Fig. 4f) and on their cell membrane (Fig. 4h). There was NCAM+ staining in several cells within the colony (Fig. 4k), resulting in a double staining of these two markers (Fig. 4l). An enhanced labeling of EpCAM was observed at the edges of the colony, a finding previously found in very primitive stem cells from the liver and biliary tree; these primitive stem cells do not express EpCAM but acquire it as they begin to undergo differentiation toward mature cells.41,42 Thus, EpCAM is not the earliest marker of hepatic stem cells but an intermediate one. The ischemic conditions have been apparently selected for more primitive stem cell subpopulations from the tissues. EpCAM and NCAM staining of the T48 group was identical to that observed in the T0–24 group (data not shown).
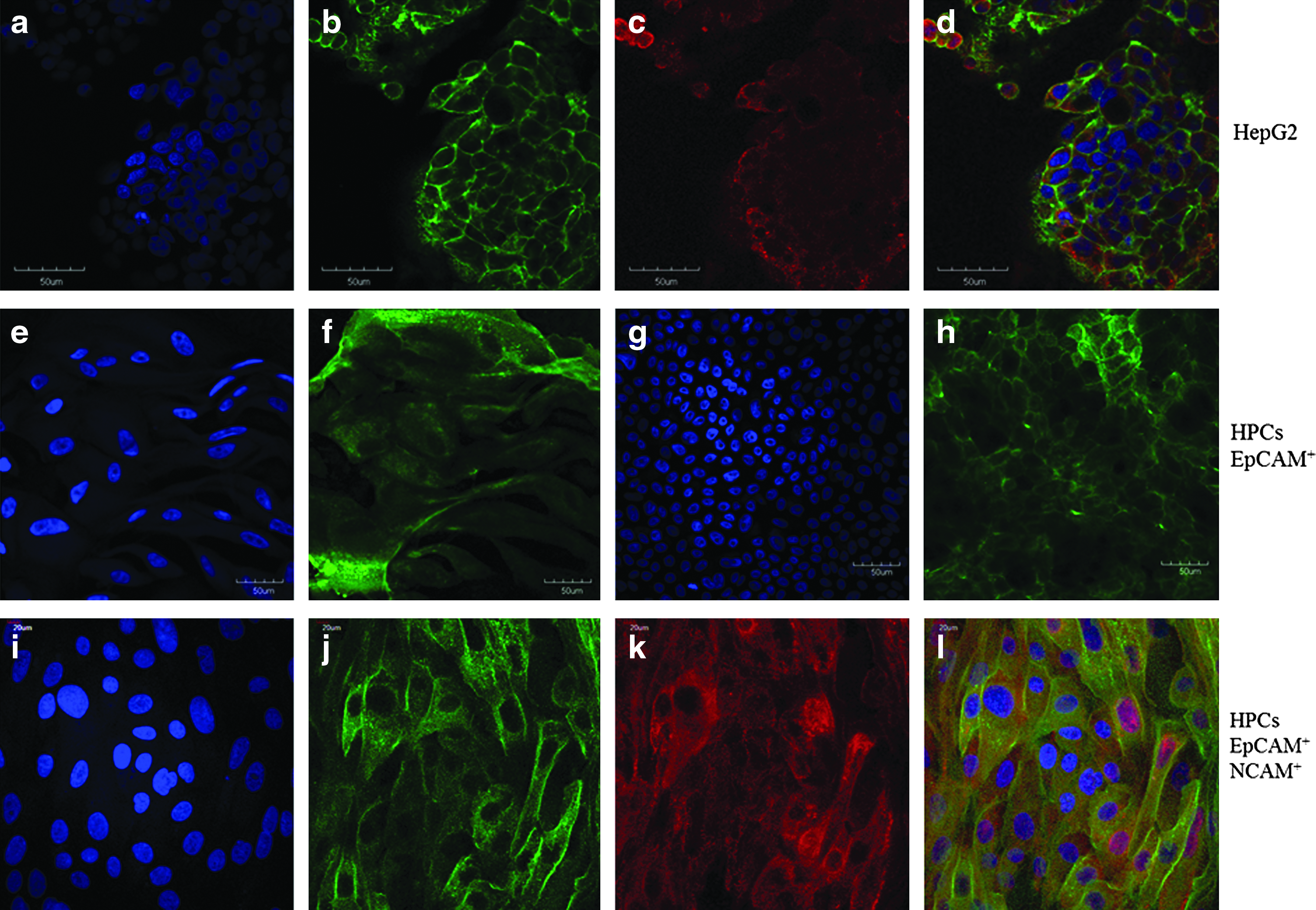
Confocal microscopy of hepatic progenitor cell colonies derived from ischemic livers. HepG2 cell line was used as a positive control for EpCAM and NCAM expression and co-expression markers
Discussion
Liver cell therapies are the hope for future treatments of patients with liver disease, a hope that depends on sourcing of tissues and on identifying strategies for cryopreservation and thus stockpiling of cell populations for use in the treatments. 6 In addition, liver cell therapies can be achieved for multiple recipients with cells from a single donor. Efforts are ongoing with immunocompromised hosts being used to assess the efficacy of various stem cell and mature cell populations.7,43 Stem cell and progenitor cell populations are logical candidates, given their capacity for expansion, cryopreservation, and maturation to mature cell types.44–46 An advantage of obtaining these cells from adult tissues is that the ethical and legal issues are minimal. If adult liver tissue can be used as a source, it also makes possible the isolation of cell populations from biopsy and from resected liver tissue for use in autologous transplantation strategies, thus eliminating immune rejection phenomena. We assessed the influence of cold ischemia on the efficiency of HPC isolation. Indeed, several reports suggested that a selection of liver stem cells after a prolonged cold ischemia period could be possible. Schmelzer et al. (2007) reported that hHpSCs were evident in cold ischemic livers from all age donors but did not perform sufficient livers under ischemic conditions to achieve statistical significance. Jozefczuk et al. reported a high resistance of adult HPCs to hypoxia and ischemia and successes in their isolation from livers after 7 days of cold ischemia.16,32 Reid et al. 26 reported that hHpSCs, in contrast to hHBs, were viable for several days after cardiac arrest (and even with warm ischemia), confirming their extraordinary tolerance to ischemia.16,17 In the present work, we compared the yield, the morphology, and the immunological characteristics of cells isolated from the same livers when submitted to short ischemia times (T0–24 group) and longer ischemia (T48 group). Whatever the duration of cold ischemia, the EpCAM+ cells remained morphologically identical in cultures and were sustained in their expression of EpCAM and NCAM both in freshly isolated cells and in the cultures. Jozefczuk et al. described two types of cells that could respectively correspond to a cell population already committed to biliary differentiation, as they expressed CK7 and CK19 markers 47 and another cell population committed to hepatocyte differentiation, because they expressed albumin and cytochrome P450. 48 The finding that stem cells can be successfully isolated from human livers after their storage for several days in cold buffer without any oxygenation before being processed opens the possibility of using liver samples from nonheart beating donors, which may represent a considerable source for HPCs for future experimental studies and for clinical programs. 12
We succeeded in isolating EpCAM+ cells utilizing the protocol developed for the isolation of hepatic progenitors from livers of all donor ages.19–23 Interestingly, we were able to isolate ∼100,000 (105) EpCAM+ cells/gram of tissue with 24 h of cold ischemia and with comparable numbers from livers even after 48 h of cold ischemia. These numbers are very similar to those obtained in control adult liver; indeed, the average number of cells/gram in adult human liver being ∼1–5×108, the percent of EpCAM+ cells (0.5%–1.5%) corresponds to 5-10×105 cells. Moreover, an average adult liver is ∼1500–2000 g, meaning that the protocol can isolate, theoretically, ∼109 EpCAM+ cells from an average adult liver. Interestingly, the yields from the livers subjected to up to 48 h of cold ischemia are very similar to those found from livers with minimal, if any, ischemia. 16 The clinical trials ongoing for cell therapies with EpCAM+ cells have utilized 80 million cells per treatment. This translates to ∼100 patients who might be treated by the EpCAM+ cells isolatable from an adult human liver. In cells isolated from all donors, in both the T0–24 and T48 groups, the EpCAM+-labeled cells strongly expressed EpCAM mRNA, to a lesser extent NCAM, CK18, and CK19, while the mRNA expression of the mature liver cell marker albumin was only marginally expressed as expected for HPCs.21,27,41
In cultures, we observed the emergence of progenitor cell colonies within 48 h of culture of EpCAM+-sorted cells on plastic and in Kubota's Medium. The morphology of the cell colonies, even from livers after 48 h of cold ischemia, was similar to that for colonies of hHpSCs and hHBs from fetal and adult human livers.24,49 The cells we obtained in culture appeared to be stem cells and not hepatoblasts for three reasons:
• it has been shown that hepatoblasts survive only about a week on plastic support and in Kubota's Medium (they require additional matrix and growth factor signals), whereas stem cells can survive 6 months or longer under these conditions;
12
• stem cells are EpCAM+ and NCAM+ but negative for alpha-fetoprotein and ICAM-1, whereas hepatoblasts are EpCAM+ but NCAM- and more positive for albumin, and strongly positive for ICAM-1 and alpha-fetoprotein;25,50 • the EpCAM+ staining in hHpSCs is throughout the cell and at the plasma membrane, whereas in hepatoblasts it is confined to the plasma membrane.12,37
Our colonies increased in size and survived for up to 3 months when cultured on plastic support and in Kubota's Medium. Three stages were observed: a phase of extensive expansion, a plateau phase, and then finally, cell collapse. This is suggestive of stem cells but ones that have clearly been affected by the ischemic conditions. It is likely that other culture conditions will have to be adopted to enable the cells to overcome whatever has occurred from the ischemic conditions. Candidate modifications are to embed the cells in hyaluronan hydrogels that have been shown to be permissive for hHpSCs to be maintained essentially indefinitely and with expansion (vs. differentiation) occurring depending on factors added to Kubota's Medium and the hyaluronans. 51 Our isolated cells were EpCAM+ NCAM+ and ICAM-1-, this label having been observed only in hHpSCs.16,25,29,30 In addition, the EpCAM+ staining we obtained was both on the cytoplasm and on the cellular membrane. Taken together, these findings suggest that we successfully isolated HPCs from adult human liver that express morphological and immunological characteristics of hHpSCs. Proof of this interpretation will have to derive from the findings of future studies assessing self-replication and multipotency.
In conclusion, we have successfully isolated HPCs from adult human livers after up to 48 h of cold ischemia. The cells isolated were identified as probable stem cells based on their ability to survive and expand in culture on plastic and in Kubota's Medium and given their gene expression properties. Our results open new opportunities for sources of HPCs for cell therapy, such as livers no more suitable for organ transplantation due to excess of ischemia period and nonheart beating donors, given HPCs will show efficacy in migrating and engrafting into the recipient livers of animal models of liver failure is currently under assessment in the animal models of liver failure developed by our group.
Footnotes
Acknowledgments
This work has been supported as a part of the HepaTTox-Cell Project (A0910010I) supported by OSEO réseau Est, and Franche-Comté and Alsace regions. The authors would like to thank Lola M. Reid at the university of North Carolina for critical review and useful discussions and also Eliane Wauthier at the university of North Carolina for useful discussions. They thank J. Godet, biostatistics consultant in Strasbourg teaching hospital, France, for performing a statistical analysis; Technical assistance of L. Coste and A. Baze is acknowledged and V. Mougey at the french blood agency (EFS), France, for performing flow cytometry analysis.
Disclosure Statement
The authors declare that there are no conflicts of interest.