
Abstract
The murine-derived MC3T3-E1 cell line provided by the American Type Culture Collection (ATCC) is a well-known osteogenic cell culture model system to test materials in vitro. However, the effect of passaging on its mineralization capacity has never been described and their culture supplements can be further optimized. Therefore, we evaluated the influence of the passage number and different osteogenic culture supplements, including ascorbic acid (AsAP) and dexamethasone (Dex) on the osteogenic capacity of MC3T3-E1 cells. This capacity was measured by the deposited calcium, the alkaline phosphatase activity, and the expression of osteogenic-related genes, including bone sialoprotein (BSP), osteocalcin (OC), and osteopontin (OPN). The results indicated that the mineralization capacity of MC3T3-E1 cells significantly decreased during passaging and got exhausted at passage 34, as assessed by measuring calcium deposition after 28 days of osteogenic induction. Moreover, the combination of AsAP and Dex triggered significantly more mineralization in MC3T3-E1 cells than the ATCC recommended addition of AsAP alone, as indicated by increased calcium deposition and higher expression of BSP and OPN. However, Dex alone could not trigger this effect, but only in combination with the AsAP, which indicates that Dex has no direct effect on mineralization. In conclusion, the passage number of MC3T3-E1 cells is of great importance and the use of cells above 30 passages should be avoided. In addition, the favored osteogenic supplements providing an improved osteogenic differentiation of MC3T3-E1 cells are the combination of AsAP and Dex.
Introduction
C
However, it is known that the number of passages can influence cellular function. Significantly decreased differentiation potentials of mesenchymal stem cells due to passaging have been observed. 5 For example, a previous study showed that undifferentiated rat bone marrow cells lost their osteogenic potential after three times of passaging. 6 In another study, it was demonstrated that the osteogenic capacity of adipose-derived stem cells also gradually decreases during long-term manipulation. 7 This can also occur with cell lines, which involves the risk that they are used beyond safe passage. 8 This is important because it is known that in studies concerning osteogenesis, and in which MC3T3-E1 cells were used, the passage number was not always clearly defined, which is an obstacle to make an objective evaluation and comparison among different studies. Cell lines are usually obtained from colleagues or another source without the information of the passage number, increasing the likelihood of leading to inaccurate or irreproducible results. 8 The osteoblastic function of MC3T3-E1 cells, as measured by osteocalcin (OC) secretion and alkaline phosphatase (ALP) activity, decreases significantly at late passage (>65) as compared to the early passage (<20). 9 Additionally, population doubling time of MC3T3-E1 cells declines from 15 h at passage 30 to 20 h at passage 65. 10 However, the mineralization ability of MC3T3-E1 cells during continuous passaging, as measured by the deposited amount of calcium has not been reported, and whether there exists a difference in the osteoblastic function between early and late passages still needs to be clarified. Another question of importance is how often MC3T3-E1 cells can be passaged before they lose their osteogenic capacity, so that the cells can be used for research without over-passaging.
In addition to the passage number, the culturing conditions are also essential for osteogenic differentiation. It is well known that the presence of beta-glycerophosphate (βGP) is required for the mineralization of mature cultures. 3 In addition, The most widely used osteogenic supplements include ascorbic acid (AsAP) and dexamethasone (Dex). AsAP is essential for the maturation and deposition of collagen and it contributes to the induction of ALP activity. 3 It has been reported that MC3T3-E1 cells exhibit high levels of osteoblast differentiation with the addition of βGP and AsAP. 4 Therefore, the combination of these two supplements is recommended by the American Type Culture Collection (ATCC) as the osteogenic stimulator for MC3T3-E1 cells. Another frequently used osteogenic inducer is Dex, which has been employed for osteogenic induction of a variety of cells, including mesenchymal stem cells,11,12 gingival fibroblasts, 13 periosteal-derived cells, 14 and dental follicle cells. 15 Dex treatment can increase the ALP activity of mesenchymal stem cells, and was essential for their mineralization of the extracellular matrix. 12 Till now, the effect of Dex on the osteogenic ability of MC3T3-E1 cells has not been described. In addition, the osteogenic capacity of different combinations of these culturing supplements remains to be investigated.
Based on the aforementioned research gaps, the aim of the present study was to define the effects of continuous passaging and composition of culturing supplements on the osteogenic ability of MC3T3-E1 cells.
Materials and Methods
Cell culture and seeding
MC3T3-E1 Subclone four cells (ATCC CRL-2593) from passage 16 were cultured in a proliferation medium containing the αMEM (Gibco) supplemented with 10% FCS (Gibco) and 100 units/mL pen/strep (Gibco) in a humidified incubator set at 37°C and 5% CO2. Upon 80% confluency, cells were detached using trypsin/EDTA (0.25% w/v trypsin/0.02% EDTA) and continuously subcultured at a density of 3500 cells/cm2 for 3 days or 2500 cells/cm2 for 4 days. The cells were expanded for five passages and afterward frozen in the α-MEM (Gibco) containing 20% FCS (Gibco) and 10% DMSO (Sigma) at passage 21 in liquid nitrogen for future use. From passage 22, cells were seeded every four passages (passage 22, 26, 30, 34, 38, 42, 46, and 50) at a density of 5000 cells/well in 24-well plates, and 10,000 cells/well in12-well plates in a control medium [proliferation medium+10 mM βGP (Sigma)], control medium+50 μg/mL AsAP (Sigma), control medium+10 nM Dex (Sigma), or control medium+combination of AsAP and Dex. On days 1, 4, and 7, the morphology of the cells was observed by an inverse phase-contrast microscope (Leica DMIL) at×10 magnification.
Cell viability
The viability of cells from passage 22 through passage 50 was evaluated every four passages using a Live/Dead Viability/Cytotoxicity Kit (Molecular Probes) after 2 days of culture. Live cells were labeled with calcein AM and dead cells were labeled with ethidium homodimer. Cells were washed in phosphate-buffered saline (PBS), incubated in 2 mM calcein-AM and 4 mM ethidium homodimer in PBS for 30 min at 37°C, rinsed in PBS for two times, and then observed by using Zeiss Imager Z1 together with the AxioCam MRc5 camera using AxioVision 4.6.3 software (Carl Zeiss Microimaging GmbH).
Calcium content
After 14, 21, and 28 days of culture, the amount of deposited calcium by MC3T3-E1 cells in 24-well plates was quantified (n=4) using the ortho-cresolphthalein complexone method (Sigma). Briefly, cells were washed twice with PBS, and then incubated in 1 mL of 0.5 N acetic acid on a shaking platform overnight. For analyses, a 10 μL sample or standard was incubated with 300 μL of work reagent in a 96-well plate at room temperature for 10 min. The standards (0−100 μg/mL) were generated by serial dilutions of a CaCl2 stock solution. Then, the plate was read in an ELISA reader (Bio-Tek Instruments) at 570 nm.
DNA content
After 14 days of culture, cells were washed with PBS, homogenized in 1 mL of MilliQ water, frozen, and thawed twice before analysis. The analysis was performed by using a PicoGreen dsDNA quantification kit (Molecular Probes) following the manufacturer's instructions. Briefly, 100 μL of the DNA sample or standard was incubated with 100 μL of a working solution in the dark for 10 min. Then, the DNA content was measured using a fluorescence microplate reader (Bio-Tek Instruments) with an excitation filter 485 nm and emission filter 530 nm.
ALP activity
ALP activity assay was performed after 14 days of culture according to a previous study on MC3T3-E1 cells. 16 Cells were harvested the same way as the samples for the DNA assay. For the analysis, 80 μL of a sample or standard (serial dilutions of 4-nitrophenol at the concentrations of 0–25 nM) and 20 μL of a buffer solution (0.5 M 2-amino-2-methyl-1-propanol) were added into a 96-well plate. Then, 100 μL of a substrate solution (p-nitrophenyl phosphate) was added to all the wells and subsequently, the mixture was incubated at 37°C for 1 h. After incubation, the reaction was stopped by adding 100 μL of 0.3 M NaOH, and the ALP activity was measured in an ELISA reader (Bio-Tek Instruments) at 405 nm and it was normalized to the DNA content.
RNA isolation
After 14 days of culture, the total RNA was isolated from the cells in 12-well plates (n=3) using the TRIzol® reagent (Invitrogen) according to the manufacturer's instructions. In brief, after removing the culture medium, 1 mL of the TRIzol® reagent was added to each well for cell homogenization. The cell extract was mixed vigorously with 0.2 mL of chloroform and centrifuged at 12,000 g for 15 min at 4°C. The aqueous phase of the sample was collected and mixed with 0.5 mL of 100% isopropanol. After incubation at room temperature for 10 min, the extract was centrifuged and then washed with 75% ethanol. Successively, the RNA pellet was dissolved in RNase-free water and the concentration and purity were determined by NanoDrop (ND-2000; Thermo Scientific). RNA samples with an A260/A280 ratio <1.8 were excluded.
Reverse transcriptase reaction
After the isolation of RNA, the reverse transcriptase reaction was performed with Superscript™ III First-strand Synthesis System for RT-PCR (Invitrogen) following the manufacturers' protocol. Briefly, the collected 1 μg total RNA in 8 μL RNA free water, 0.5 μL random hexamers, 0.5 μL oligo (dT) primers, and 1 μL of dNTPs were incubated for 5 min at 65°C. Then, the following components were subsequently added: 2 μL reaction buffer, 4 μL 25 mm MgCl2, 2 μL 0.1 m DTT, 1 μL RNase OUT, and 1 μL Superscript III RT. The reaction mix was incubated for 10 min at 25°C for further primer annealing, 50 min at 50°C for reverse transcription, and 5 min at 85°C to terminate the reaction. This cDNA solution was stored at −20°C for further use.
Real-time PCR
The cDNA was further amplified and the expression of specific genes was quantified using the IQ SYBR Green Supermix PCR kit (BioRad) in a real-time PCR (BIORAD, CFX96™ real-time system). For this reaction, 2 μL cDNA, 1.5 μL former and 1.5 μL reverse primers, 12.5 μL master mix, and 7.5 μL RNase-free water were mixed and centrifuged. Subsequently, PCR was performed at 60°C. Osteoblastic differentiation-related primers were used to check the gene expression at the RNA level, including bone sialoprotein (BSP), OC, and osteopontin (OPN). GAPDH was referred as the housekeeping gene (sequences in Table 1). Specific sense and antisense primers for the genes were designed according to published cDNA sequences of GenBank. The specificity of the primers was tested separately before the reaction and the negative control consisted of RNase-free water. The calculation of gene expression was performed via the 2−ΔΔCt method. 17
Statistical analysis
The calcium content and ALP activity assays were always performed in quadruplicate and samples for real-time PCR were assayed in triplicate. All data were reported as mean±SD. Statistical analysis of quantitative data was performed by using one-way ANOVA combined with the Student–Newman–Keuls multiple comparison post test and the significance level was 0.05 (Instat 3.05; GraphPad Software, Inc.).
Results
Cell morphology
To evaluate the effect of the passage number and culturing supplements on the morphology of MC3T3-E1 cells, morphological observation was performed on days 1, 4, and 7. No change in MC3T3-E1 cell morphology was observed among all the passages. On day 1, MC3T3-E1 cells displayed a typical fibroblast-like appearance regardless of culturing supplements (Fig. 1A). On day 4, all the cells exhibited short spindle-shaped morphology and no difference was observed in different groups (Fig. 1B). On day 7, all the cells grew in multilayers and lost the fibroblastic characteristics (Fig. 1C, D). However, most cells in the control and AsAP groups showed a cuboidal appearance (Fig. 1C), while cells in the Dex and AsAP/Dex groups developed an elongated shape (Fig. 1D).

The effect of culturing supplements on the morphology of MC3T3-E1 cells. On day 1, MC3T3-E1 cells exhibited a typical fibroblast-like appearance regardless of culturing supplements
Cell viability
To detect whether the passage number and culturing supplements influenced the viability of MC3T3-E1 cells, cell survival was assessed after 2 days of culture using a live/dead assay from passage 22 through passage 50 every 4 passages. Qualitative data (Supplementary Fig. S1; Supplementary Data are available online at
Calcium content
To investigate the effect of continuous passaging and culturing supplements on the mineralization of MC3T3-E1 cells, cells were cultured in a control medium, or a control medium containing either AsAP, Dex, or AsAP/Dex for up to 28 days. From passage 22 through passage 50, mineralization was evaluated through quantitative calcium content analysis every four passages on day 14, 21, and 28. Over the first 3 weeks of osteogenic induction, no calcium deposition was detected for any of the groups. By the 28th day of culture, a substantial amount of calcium deposition was detected with cells cultured in AsAP and AsAP/Dex from passage 22 through 30 (Fig. 2). A dramatic decrease in calcium deposition was observed at an increasing passage number, and no calcium deposition at all was found from passage 34 (Fig. 2). The addition of only Dex led to no change in the calcium content as there was no significant difference with the control culture (Fig. 2). In contrast, the mineralization capability of MC3T3-E1 cells was enhanced significantly by the addition of AsAP or AsAP/Dex as compared to the control and Dex groups (p<0.05). Moreover, the enhanced calcium content was more prominent in cell cultures supplemented with AsAP/Dex than only AsAP (Fig. 2). At passage 22 and 26, the addition of AsAP/Dex stimulated significantly more calcium deposition than that in the group with only AsAP (p<0.01), with an increase of approximately 40 μg/mL for each passage (Fig. 2).
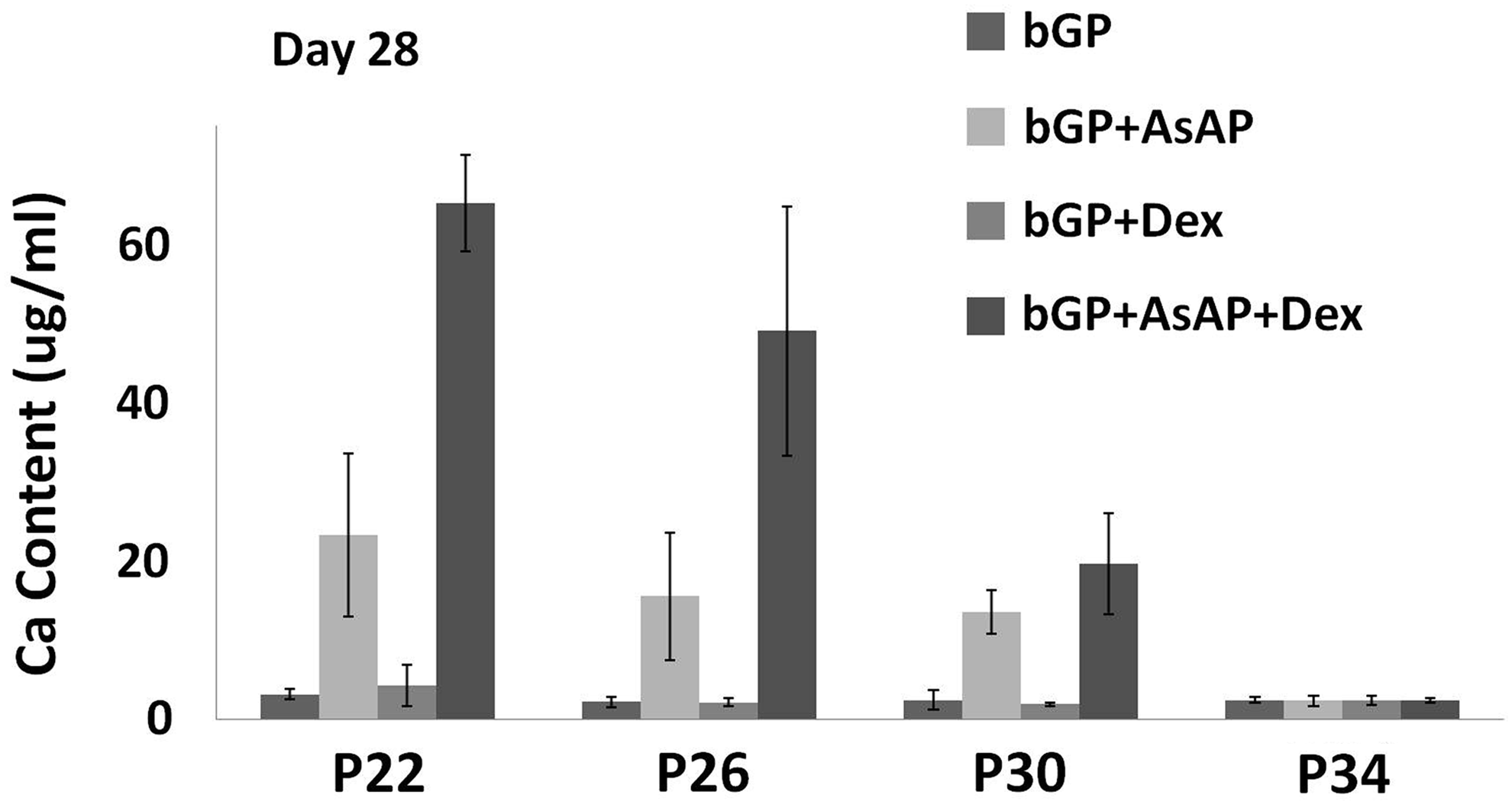
The effect of passage number and culturing supplements on the mineralization capacity of MC3T3-E1 cells. The results indicate that MC3T3-E1 cells display a significant decrease in the calcium content during the process of continuous passage and lose the osteogenic capacity within 34 passages. The combination of AsAP and Dex stimulates a higher calcium deposition by the cells than a single factor does. Error bars represent standard error of the mean (n=4).
ALP activity
To evaluate the effect of continuous passaging and culturing supplements on the osteogenic capacity of MC3T3-E1 cells, the ALP activity on day 14 was further measured at passage 22, 26, and 30. Comparable to the calcium content study, Figure 3 showed significantly higher levels of the ALP activity for cells at passage 22 and 26 versus cells at passage 30. In contrast to the calcium content study, the addition of AsAP or AsAP/Dex resulted in a lower ALP activity in MC3T3-E1 cells compared to the control group and Dex alone group (Fig. 3).
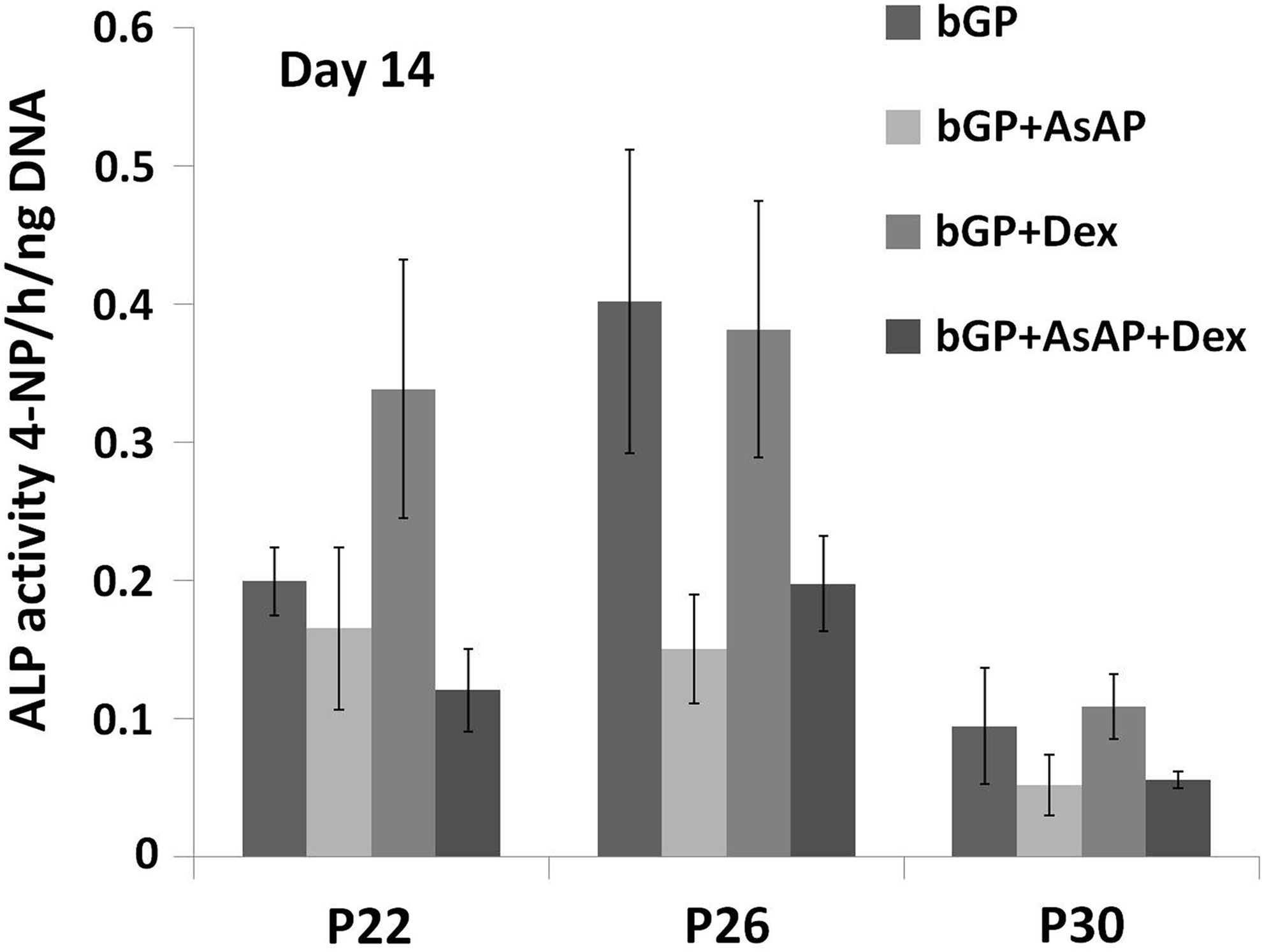
The effect of passaging and culturing supplements on the alkaline phosphatase (ALP) activity of MC3T3-E1 cells. Significantly higher levels of the ALP activity were observed for cells at passage 22 and 26 versus cells at passage 30. The addition of AsAP or AsAP/Dex did not stimulate a higher ALP activity in MC3T3-E1 cells.
Gene expression
To verify the effect of osteogenic culture supplements on the osteogenic capacity of MC3T3-E1 cells, expression of genes involved with osteogenesis (BSP, OC, and OPN) was further determined by performing real-time quantitative RT-PCR on the cells cultured for 14 days. Quantification of gene expression was only performed at passage 22, 26, and 30 since these cells still possess the capacity to mineralize. The Dex alone group was excluded because no calcium deposition was detected for this group. The highest expression levels of BSP were seen in the AsAP/Dex groups, which displayed a significantly higher expression of BSP expression than the AsAP alone groups (Fig. 4A). For OC gene expression, a significant increase was seen when AsAP or AsAP/Dex was added compared to the control group (Fig. 4B). There was also an increase in the AsAP/Dex group compared with the AsAP alone group, but this increase was not significant. As shown in Figure 4C, a significantly higher expression of OPN was found by the addition of AsAP/Dex compared to the control and AsAP groups at passage 22.

The effect of culturing supplements on the expression of bone sialoprotein (BSP), osteocalcin (OC), and osteopontin (OPN). Cells cultured with AsAP/Dex displayed significantly higher levels of BSP, compared to those of the AsAP group
Discussion
The purpose of the current study was to investigate the effects of Dex and AsAP on the mineralization of MC3T3-E1 cells at different passages. We hypothesized that the osteogenic ability was compromised in MC3T3-E1 cells undergoing continuous passaging. Secondly, it was supposed that a combination of osteogenic supplements would further stimulate mineralization. Our results demonstrated a significantly decreased mineralization capacity of MC3T3-E1 cells during passaging without negatively affecting their viability (determined by deposited calcium content). At 34 in vitro passages, the mineralization capacity of the cells was observed to be exhausted as there was no calcium deposition. The osteogenic ability of MC3T3-E1 cells, as induced by the combination of AsAP and Dex and measured by the calcium content and expression of osteogenetic genes (BSP and OPN), was significantly higher than the traditional addition of AsAP alone.
The observed reduced osteogenic capacity during passaging is in agreement with previous research. 9 Chung et al. reported that the osteoblastic function of MC3T3-E1 cells from late passage (>65) was significantly inhibited as compared to the early passage (<20). 9 Comparable to their results, significantly higher levels of the ALP activity for cells at passage 22 and 26 were detected in comparison to cells at passage 30. Our data also indicate that MC3T3-E1 cells are losing their functional capacity of mineralized nodule formation between passage 30 to 34 without affecting their morphology and viability. The effect of passage number on cell lines is not limited to MC3T3-E1 cells. It has been reported that the passaging process significantly influences the physiological development and transport properties of the Caco-2 cell line.18,19 This change in behavior can be caused by genetic mutations,8,18 or due to a positive selection of fast-growing cells from the population of heterogeneous cell lines during passaging,8,18 since heterogeneity has been observed in the MC3T3-E1 cell line as well as in other cell lines.4,20
Recently, more and more national and international collaborations take place, which favor scientific discovery significantly. The exchange of cell lines between laboratories is frequently part of the collaborative efforts. However, this harbors the risk that important information, like the passage number of the cells, gets lost. Our results suggest that over-passaged cells and cell lines without a defined passage number should not be used.
The other purpose of the current study was to further optimize osteogenic culture conditions of MC3T3-E1 cells, of which, the traditionally supplemented osteogenic inducer is AsAP. 4 In contrast, rat bone marrow cells mineralize only when Dex is added to the culture medium. 6 Therefore, we decided to assess whether Dex can further optimize the osteogenic induction of MC3T3-E1 cells. The used doses were chosen according to previous studies, in which, 50 μg/mL AsAP triggered an optimal increase in osteogenic differentiation of osteoblast-like cells from the human facial bone 21 and 10 nM Dex was the optimal condition for mineralization of human mesenchymal stem cells. 22 However, it has to be pointed out that different species may have their optimal doses for osteogenic culture supplements. Therefore, different doses of AsAP and Dex for mouse MC3T3-E1 cells may give rise to different results. For RT-PCR analysis, BSP and OC were selected as discriminatory genes, because a good correlation has been observed between their mRNA expressions and the ability of a given subclone to form a mineralized extracellular matrix in culture. 4
Our results indicated that the combination of AsAP and Dex promotes osteogenic ability of MC3T3-E1 cells when compared to AsAP alone, as shown by the significantly higher calcium deposition. These results highlight the importance of Dex in influencing the mineralization of MC3T3-E1 cells. However, no change in mineralization was observed in the Dex alone group as compared to the control, indicating Dex has an indirect effect on mineralization of MC3T3-E1 cells. This investigation was described similar by others. 14 Roberts et al. reported that Dex augmented osteogenic differentiation of periosteal-derived cells in combination with all trans-retinoic acid (atRA) and bone morphogenic protein 2 (BMP2), but had no direct effect on the differentiation. 14 This phenomenon can be interpreted that Dex acts directly and/or indirectly to promote the osteoprogenitor cell numbers in cultures. 23 The higher cell numbers stimulated by Dex can also explain the elongated cell shape in Dex groups on day 7, as there is a significant amount of overlap between the cells.
Another surprising phenomenon that has to be pointed out is, in contrast to the mineralization result, that the addition of AsAP or AsAP/Dex stimulated a lower ALP activity in MC3T3-E1 cells compared to the control group and Dex alone group. This is comparable to another recently published study, in which, inositol hexakisphosphate (IP6) decreased mineralization and gene expression of osteoblast markers in MC3T3-E1 cells without negatively affecting the ALP activity. 16 Besides, Beck et al. reported that ALP levels can be dissociated from mineralization of MC3T3-E1 cells, as mineralization can occur in the absence of ALP induction. 19 Cell lines exhibiting severe repression of the ALP activity can differentiate to a mineral-secreting phenotype. 19 These results may indicate that ALP activity alone is not sufficient for the determination of the osteogenic capacity of MC3T3-E1 cells.
In conclusion, our findings indicate that (1) the passage number of MC3T3-E1 cells should be clearly defined in all scientific studies; (2) the use of MC3T3-E1 cells for osteogenic differentiation should be avoided after passage 30; (3) AsAP and Dex are suggested as the supplements for the osteogenic induction of MC3T3-E1 cells.
Footnotes
Acknowledgment
Xiangzhen Yan and Wanxun Yang are grateful to the program of China Scholarship Council for the financial support (No. 2010622061, 2010627030). This work was supported by the Royal Netherlands Academy of Arts and Sciences (KNAW; project number PSA 08-PSA-M-02).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.